










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.25 n.1 Bogotá jan./jun. 2007
Sandra Aragón1, Fernando Cantor1, José Ricardo Cure2 y Daniel Rodriguez2
1 Bióloga, Universidad Militar Nueva Granada, Bogotá. e-mail: sandra_aragon@hotmail.com
2 Docentes, Facultad de Ciencias, Programa de Biología Aplicada, Universidad Militar Nueva Granada, Bogotá. e-mails: fcantor@umng.edu.co; jrcure@umng.edu.co; daniel.rodriguez@umng.edu.co
Fecha de recepción: 21 de noviembre de 2006. Aceptado para publicación: 06 de junio de 2007
Resumen: Praon pos. occidentale es un parasitoide promisorio para regular poblaciones de áfidos Macrosiphum euphorbiae en cultivos comerciales bajo invernadero de rosas. Se evaluó la capacidad parasítica de P. pos. occidentale bajo tres temperaturas constantes (18, 25 y 28ºC) y variando la densidad de su hospedero (5, 10, 20, 40, 80 y 150). En cada unidad experimental se liberó una pareja del parasitoide con 24 horas de edad y se mantuvieron durante 24 horas en incubadoras graduadas a cada temperatura en evauación, con 12 horas de luz y 12 de oscuridad. Se registró el número de áfidos parasitados y se permitió el desarrollo de los huevos depositados hasta adulto. Se estimó la tasa instantánea de búsqueda (a’) y el tiempo de manipulación (Th) a partir de los cuales se ajustó el modelo de respuesta funcional tipo II para cada temperatura. La tasa instantánea de búsqueda (a’) fue más alta a 18ºC con un valor de 0,1081, seguida de 28ºC con 0,0323 y 25ºC con 0,0103. El tiempo de manipulación (Th) más corto fue el que se presentó a 25ºC de 4,8913, seguido de 28º C con un tiempo de 5,7579 y 18ºC con 8,2697. El máximo número de individuos parasitados estimado fue de 4,9 a 25ºC. A 18ºC el 60% de los áfidos parasitados alcanzó la emergencia del adulto, el 74,2% a 25ºC y el 88% a 28ºC. No existe ningún efecto significativo de la densidad del hospedero ni de la temperatura en la proporción sexual de Praon pos. Occidentale.
Palabras clave: control biológico, parasitoides, áfidos, respuesta funcional, cultivos bajo invernadero.
Abstract: Praon pos. occidentale is a promising parasitoid that regulates populations of the aphid Macrosiphum euphorbiae in commercial rose production under greenhouse conditions. It was evaluated the parasitic ability of Praon pos. occidentale under three constant temperatures 18, 25 and 28ºC and varying host density 5, 10, 20, 40, 80 y 150. Each experimental unit consisted of one couple of the parasitoid with 24 hours old kept during 24 hours in graduated incubators with a 12:12 photoperiod. The number of parasitized aphids was recorded and development of the laid eggs was allowed. The instantaneous rate of search (a’) and the handling time (Th) of the model of functional response type II were estimated. The instantaneous rate of search (a’) was higher at 18ºC with 0.1081, followed by 0.0323 at 28º C and 0.0103 at 25ºC. The shorter handling time (Th) was 4.8913 at 25ºC, followed by 5.7579 at 28ºC and 8.2697 at 18ºC. The maximum number of parasitized aphids estimated was 4.9 at 25ºC. At 18ºC the 60% of the parasitized aphids reached the adult emergence, whereas this number was 74.2% at 25ºC and 88% at 28ºC. There was no significant effect of the host density, neither temperature on the sexual ratio of Praon pos. occidentale was recorded.
Key words: biological control, parasitoids, aphids, functional response, commercial greenhouse crop.
Introducción
Se considera al áfido Macrosiphum euphorbiae (Hemiptera: Aphididae) una de las plagas más importantes de cultivos de invernadero porque es capaz de generar grandes pérdidas económicas. Para su control se han sugerido diferentes estrategias entre las que se encuentra el uso de enemigos naturales, principalmente en Europa y Estados Unidos.
En Colombia son escasos los estudios de control biológico de áfidos en cultivos de hortalizas y ornamentales mantenidos bajo invernadero. Sin embargo, se han realizado algunos aportes del Centro de Investigaciones y Asesorías Agroindustriales (CIAA) de la Universidad Jorge Tadeo Lozano, en conjunto con investigadores de la Universidad Militar Nueva Granada (UMNG) en la Sabana de Bogotá. En una evaluación de diferentes enemigos naturales introducidos para el control de plagas en cultivos comerciales de rosa se encontraron avispas del género Praon pos. occidentale (Hymenoptera: Braconidae) atacando naturalmente poblaciones de áfidos M. euphorbiae con mayor eficiencia que los enemigos naturales introducidos para el control de los mismos áfidos.
En consecuencia de lo anterior, se iniciaron estudios para evaluar el potencial biológico de Praon pos. occidentale que estuvieron relacionados específicamente con la duración de su ciclo de vida a diferentes temperaturas (Arias, 2003). Con el fin de complementar la información relacionada con el potencial biológico de estas avispas se evaluó la capacidad controladora de P. pos. occidentale en las instalaciones del CIAA, ubicado en Chía (Cundinamarca), por medio de la estimación de la respuesta funcional, el porcentaje de emergencia y la proporción sexual en condiciones de laboratorio en seis densidades de hospedero bajo tres temperaturas (18, 25 y 28ºC).
La respuesta funcional como medida de la capacidad controladora de los enemigos naturales ha sido ampliamente utilizada (Everson, 1979; Hofsvang y Hagvar, 1983; Donelly y Phillips, 2001). Según Begon et al. (1996) la respuesta funcional es la relación que existe entre la tasa de consumo de un individuo y la densidad local de alimento. Solomon (1949) la define como el cambio del número de presas consumidas por unidad de tiempo en relación con la variación de la densidad de la presa; esta respuesta se basa en la densidad de un solo tipo de hospedero o presa (Schenk y Bacher, 2002).
La respuesta funcional es un componente esencial en la dinámica de la asociación hospedero:parasitoide y es un determinante importante de la estabilidad del sistema (Wang y Ferro, 1998); puede ser utilizada como una medida de la eficiencia de búsqueda cuando se evalúan agentes potenciales de control (Waage, 1990).
Los factores que pueden afectar la respuesta funcional incluyen el tiempo de exposición de la presa al depredador, la proporción de éxito en la búsqueda y el tiempo de manipulación requeridos para cada presa. Todos estos factores están incluidos en la ecuación de disco de Holling (1959) la cual describe la respuesta curvilínea de varios insectos depredadores.
Rogers (1972) identificó que el comportamiento de ataque de los parasitoides es biológicamente diferente a la de los depredadores por lo que describió un modelo que permite la estimación de la respuesta funcional específicamente para insectos parasitoides:
Donde Na es el número de hospederos parasitados, Nt es la densidad del hospedero, a’ es la tasa instantánea de búsqueda, T es el periodo de tiempo en horas que están expuestos los áfidos al parasitoide, Pt es el número de parasitoides y Th es el tiempo de manipulación estimado en horas.
Holling (1959) clasificó la respuesta funcional en tres tipos diferentes dependiendo de la densidad ofrecida de la presa y la densidad de presas atacadas. En la respuesta funcional tipo I el número de presas atacadas por depredador es directamente proporcional a la densidad de la presa. Sin embargo, en determinada densidad de presa el depredador se sacia y no consume mas presas. En la tipo II, el número de encuentro también aumenta con la densidad de la presa, pero a una tasa decreciente debido a que consume una proporción alta del tiempo disponible en la manipulación de la presa. Finalmente, en la respuesta funcional tipo III la relación es sigmoide puesto que la tasa de incremento del ataque del depredador es lenta y va aumentando en función de la densidad de la presa, posteriormente cuando el depredador alcanza la saciedad esta tasa decrece.
La respuesta funcional tipo II o cyrtoid ha sido mostrada como la más común entre artrópodos predadores bajo condiciones de laboratorio (Holling, 1959; Hasell,1978; Kfir y Luck, 1985; Wang y Ferro, 1998) al igual que en artrópodos parasitoides (Huffaker y Gutiérrez, 1999).
Materiales y métodos
En un diseño completamente al azar con cinco repeticiones por tratamiento (densidad de hospedero) se evaluó el efecto de la temperatura (18, 25 y 28ºC ) y la densidad de áfidos ofrecidos (5, 10, 20, 40, 80 y 150 áfidos) sobre la respuesta funcional, el porcentaje de emergencia y la proporción sexual de Praon pos. occidentale. Cada temperatura se evaluó una vez dentro de incubadoras graduadas en la temperatura a evaluar y con un fotoperiodo de 12 horas de luz y 12 de oscuridad.
En recipientes de plástico con dimensiones de 30 x 20 x 25 cm con una ventana de aireación en la parte superior cubierta con malla metálica y una tira de plástico amarillo con miel, se introdujeron dos folíolos de tomate de 8 semanas de edad con el pecíolo introducido en un ‘aqua-pin’ con solución floral. Sobre cada foliolo se ubicaron 5, 10, 20, 40, 80 y 150 áfidos adultos según el tratamiento.
Una vez ubicados los áfidos se procedió a separar las 30 parejas de avispas para cada tratamiento; éstas tenían 24 horas de emergencia y no habían tenido contacto alguno con áfidos. De ésta manera se garantizó que las avispas aún poseían su carga original de huevos. Los áfidos y las avispas eran obtenidos de la cría permanente de los laboratorios de la UMNG y el CIAA; en esta cría permanente las avispas, una vez emergidas, eran aisladas por un periodo de 24 horas para asegurar la cópula.
De acuerdo con la metodología propuesta por Sigsgaard (2000), durante una hora se ubicaron los áfidos y las avispas por separado en las incubadoras con la temperatura a evaluar. Después de la hora de acondicionamiento, las avispas fueron liberadas en los diferentes tratamientos y retiradas al cabo de 24 horas. Posteriormente, los montajes eran retirados de las incubadoras y mantenidos en condiciones de laboratorio con condiciones controladas de temperatura (22ºC), humedad relativa (67%) y fotoperiodo de 12 horas luz y 12 horas oscuridad.
Para garantizar el recurso de los áfidos parentales en los diferentes tratamientos, diariamente se retiraron los nuevos áfidos emergidos con la ayuda de un compresor y un aspirador y cada tercer día se realizó cambio de folíolos hasta la formación de la primera pupa. Se registró la cantidad de áfidos parasitados reconocidos por la formación de momias blancas con una estructura conocida como ‘pedestal’ que caracteriza la pupa de este parasitoide.
Para obtener información acerca del porcentaje de emergencia y proporción sexual de este parasitoide en las tres temperaturas, se permitió la emergencia de los adultos de P. pos. occidentale y se determinó su proporción sexual comparando las características del ovipositor y la genitalia del macho con las descritas en los estudios de Schlinger y Hall (1960) y Marsh et al. (1987).
Se ajustó el modelo para parasitoides propuesto por Rogers (1972). Los parámetros a’ y Th fueron estimados mediante una regresión no lineal utilizando el procedimiento NLIN del programa estadístico SAS versión 8.0® (SAS, 1999) ingresando todos los datos originales.
Con el propósito de unificar un criterio de comparación para las curvas de repuesta funcional en las tres temperaturas, se decidió utilizar la asíntota de cada una de las curvas como el valor representativo de la máxima capacidad parasítica estimada del parasitoide. El análisis de los datos se realizó comparando el 80% del valor de esta asíntota para cada temperatura y extrapolando al valor correspondiente de densidad de hospederos ofrecida en la cual se alcanza este potencial estimado de parasitación. Se decidió utilizar el 80% del valor de la asíntota por considerar que valores superiores no representaban incrementos significativos en la tasa de ataque de los parasitoides en cada una de las tres temperaturas.
Resultados y discusión
En la figura 1 se puede observar que la respuesta funcional para las tres temperaturas fue de tipo II. La curva de 18ºC aumenta más rápido que las de 25ºC y 28ºC antes de alcanzar la fase de estabilidad. Este fenómeno es interesante ya que se esperaba que la fase inicial de la curva tuviera una tasa de incremento mayor en la medida que aumentaba la temperatura. Esta situación podría estar relacionada con una situación de estrés térmico que puede inducir el aumento de la tasa de oviposición como mecanismo de sobrevivencia.
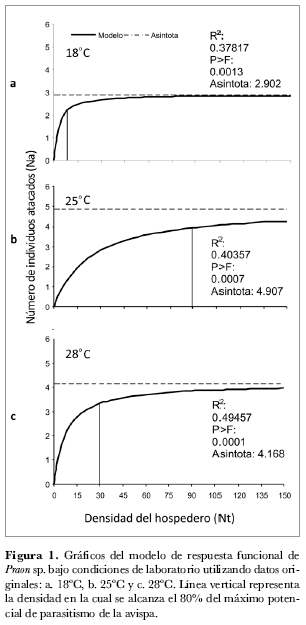
Gutiérrez (1996) y Schreiber y Gutiérrez (1998) mencionan que para estudios de respuesta funcional es necesario considerar mayor realismo, por lo que se deben incorporar parámetros de la fisiología y del comportamiento de los organismos involucrados a partir de modelos con enfoque tritrófico en donde aspectos como la calidad del alimento y otros factores ecológicos tienen un papel importante.
Al observar el máximo número de individuos parasitados en cada condición térmica representado por las asíntotas en la figura 1, se aprecia que el número aproximado de áfidos parasitados fue de tres, cinco y cuatro a 18ºC, 25ºC y 28ºC respectivamente. El máximo número de áfidos parasitados corresponde al máximo potencial biológico estimado en cada una de las temperaturas evaluadas y este control se presenta cuando se ofrecen 10 áfidos en 18ºC, 89 en 25ºC y 30 en 28ºC áfidos ofrecidos.
En la tabla 1 se resumen los resultados obtenidos para los diferentes parámetros evaluados en cada una de las temperaturas. Los valores de Th son más cercanos entre 25ºC y 28ºC que entre estos dos y 18°C. De acuerdo con la información presentada en las tablas 1 y 2 se puede observar que el máximo valor del parámetro Th se relaciona con la mínima densidad de áfidos parasitados. Estas observaciones están de acuerdo con lo reportado por Menon et al. (2002) y Hassell (1978) quienes mencionan que un tiempo de manipulación (Th) corto incrementa el tiempo disponible para búsqueda y, por tanto, la probabilidad de encontrar más hospederos.
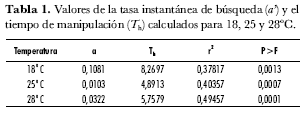
Al analizar el parámetro a’, que se refiere a la tasa instantánea de búsqueda (tabla 1) se observa que este parámetro es mayor a 18ºC. Lo anterior indica que P. pos. occidentale responde más rápido a la presencia de áfidos a bajas temperaturas, a pesar de que el máximo potencial biológico sea menor que en las otras dos temperaturas.
Porcentajes de emergencia de Praon pos. occidentale
Se observó que por cada momia presente emergía una sola avispa. Se presentaron momias de color blanco y café oscuro. Las momias blancas alcanzaban a emerger durante el período de evaluación. Por el contrario, las momias que presentaban una coloración café oscura no emergieron en este período, sino mucho tiempo después de transcurrido el tiempo de seguimiento.
En la figura 2 se observa que cuando aumenta la temperatura se incrementa la capacidad de ataque de las avispas. Esta capacidad de ataque está representada por el aumento en el número de hospederos parasitados (línea discontinua). Además se observa que el máximo número de hospederos parasitados se presenta cada vez en densidades mayores cuando aumenta la temperatura de 18ºC a 28ºC. A 18°C se presenta el máximo número de áfidos parasitados y momificados a una densidad de 20 individuos (figura 2a), a 25°C el máximo se alcanzó en una densidad del hospedero de 80 individuos (figura 2b) y a 28°C el máximo de ataque se alcanzó en 150 hospederos (figura 2c). Así mismo a 18°C el máximo promedio de avispas emergidas fue de 4,2 en 25°C fue de 3,6 y a 28°C de 6 nuevas avispas por hembra liberada (línea continua).
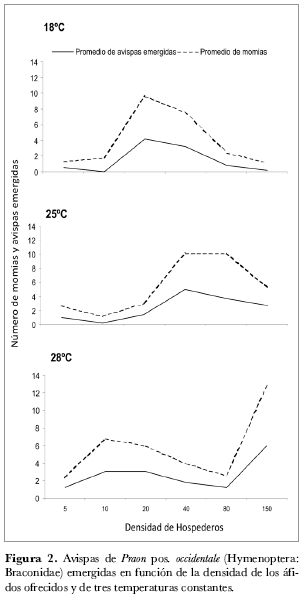
A 18ºC sólo se presentaron emergencias de nuevas avispas en el 60% del total de áfidos parasitados que presentaban la momificación, mientras que para 25ºC fue de 74,2% y para 28ºC de 88%. De acuerdo a lo anterior, se podría asumir que el aumento en el número de emergencias de nuevas avispas puede estar relacionado con el aumento de la temperatura.
Las momias que presentaron coloración café a 18°C no emergieron durante el periodo de evaluaciones, sino tiempo después cuando se dejaron a 22oC en el laboratorio. Por lo anterior, durante las evaluaciones del ensayo se registró un menor porcentaje de emergencia de nuevas avispas a 18oC pero, posteriormente a las fechas de evaluación bajo la nueva temperatura de 22oC, las avispas de este tipo de momias emergieron normalmente. Aparentemente esta coloración indica un cambio fisiológico en la pupa que retarda la emergencia de las avispas.
Proporción sexual de Praon pos. occidentale
En la figura 3a se puede observar que a 18ºC y 28ºC se presenta mayor emergencia de machos que de hembras, mientras que a 25ºC la proporción de machos y hembras es de 1:1. Estas observaciones coinciden con los resultados obtenidos por Arias (2003) quien evaluó la proporción sexual de P. pos. occidentale a 17ºC, 20ºC, 25ºC, 28ºC y 30ºC y observó que a temperaturas más bajas se presenta una mayor emergencia de machos que en las temperaturas altas. Kfir y Luck (1979), realizaron un ensayo con el género Aphytis (Hymenoptera: Aphelinidae) en el cual sugieren que las temperaturas extremas pueden reducir la viabilidad y movilidad del esperma y también pueden afectar el comportamiento de fertilización y apareamiento.
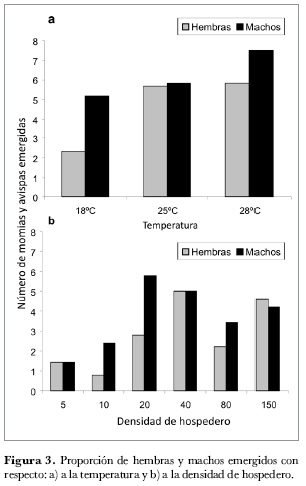
El efecto de la densidad del hospedero sobre la proporción sexual del parasitoide se presenta en la figura 3b. Al realizar las pruebas para evaluar el efecto de la densidad de los hospederos ofrecidos sobre la proporción sexual de avispas emergidas no se encontró ninguna relación significativa entre estas dos variables (P=0,05, F= 0,16 hembras y 0,41 machos). Lo anterior indica que la densidad del hospedero no afecta la proporción sexual de P. pos. occidentale.
Se ha reportado que el superparasitismo afecta la proporción sexual de avispas parasitoides debido a la mortalidad diferencial sexo-específica de huevos y larvas en desarrollo. Los machos son más aptos que las hembras para completar su desarrollo dentro de un hospedero que posea más de un huevo del parasitoide, posiblemente debido a su desarrollo más rápido o a sus requerimientos nutricionales más bajos. Como resultado, una alta frecuencia de superparasitismo puede producir una proporción sexual basada en machos (Heinz y Parrella, 1990).
Aunque no fue registrado en este trabajo, en ensayos realizados por Arias (2003) con Praon pos. occidentale a diferentes temperaturas, se presentó un alto porcentaje de superparasitismo en las seis temperaturas y altos porcentajes de emergencia de machos en las temperaturas más bajas. Por lo anterior, se podría afirmar que la alta proporción de machos encontrados en estos ensayos se realaciona directamente con este comportamiento del parasitoide. Un comportamiento similar fue observado por Schlinger y Hall (1960) quienes realizaron un ensayo donde confirmaron que el porcentaje de emergencia de machos está relacionado con la condición de superparasitismo en Praon palitans bajo condiciones de laboratorio y observaron que nunca se desarrolló más de un parasitoide por áfido.
Conclusiones y recomendaciones
Se determinaron los parámetros de las curvas de respuesta funcional de P. pos. occidentale en las tres temperaturas evaluadas (18, 25 y 28ºC). En todas las temperaturas las curvas de respuesta funcional fueron de tipo II, donde en un período de 24 horas una única avispa fue capaz de parasitar de tres a cinco áfidos diferentes.
Praon pos. occidentale responde más rápido a menores densidades de áfidos en una temperatura de 18oC. Al aumentar la temperatura de 18oC a 28oC se favorece el aumento, tanto del número de áfidos parasitados, como de avispas emergidas.
A pesar de que la respuesta de P. pos. occidentale a la presencia de bajas densidades de áfidos es más lenta en temperaturas de 25oC y 28oC, a estas temperaturas se obtienen mayores índices de parasitación y de emergencia de nuevas avispas.
A partir de los resultados y conclusiones del presente trabajo se recomienda realizar estudios que verifiquen el potencial controlador de la avispa bajo condiciones de invernadero en la Sabana de Bogotá para verificar el potencial de la avispa que se encontró en los ensayos de laboratorio.
Además, sería interesante conocer cual es el efecto de las bajas temperaturas sobre la formación de pupas de color café y su relación con posibles estados de latencia o dormancia que sólo se presentarían en países estacionales.
Se sugiere manejar los cultivos con baja presión de insecticidas para que estas avispas se presenten de manera natural. Además, se recomienda iniciar estudios de crías continuas de la avispa que sirvan de suministro constante para aquellos cultivos que sean fácilmente suscepti-bles al ataque de áfidos. En caso de iniciar estudios de cría continua de la avispa se recomienda tener en cuenta las condiciones térmicas y las densidades ofrecidas de áfidos que favorecen el desarrollo del mayor número de hembras de Praon pos. occidentale.
Agradecimientos
Al Centro de Investigaciones y Asesorías Agroindustriales de la Universidad de Bogotá Jorge Tadeo Lozano por el préstamo de sus instalaciones; a Asocolflores y la Vicerectoría de Investigaciones de la Universidad Militar Nueva Granada por la financiación del proyecto.
Literatura citada
Arias, D. 2003. Efecto de la temperatura en algunas variables de crecimiento de la población de Praon sp. (Hymenoptera: Braconidae) endoparasitoide nativo del áfido de los invernaderos Macrosiphum euphorbiae (Hemiptera: Aphididade). Trabajo de grado. Facultad de Ciencias, Universidad Militar Nueva Granada, Bogotá. 107 p. [ Links ]
Begon, M.; J. Harper y C. Tonnsend. 1996. Ecology individuals, populations and communities. 3th edition. Ed. Black Sciences, 1068 p. [ Links ]
Donelly B. y T. Phillips. 2001. Functional response of Xylocoris flavipes (Hemiptera:Anthocoridae): Effects of prey species and habitat. Environ. Entomol. 30(3), 617-624. [ Links ]
Everson, P. 1979. The functional response of Phytoseiulus persimilis (Acarina:Phytoseiidae) to various densities of Tetranychus urticae (Acarina:Tetranychidae). Can. Entomol. 111, 7-10. [ Links ]
Gutiérrez, A.P. 1996. Applied population ecology: a supply-demand approach. John Wiley & Sons, Inc., New York. 300 p. [ Links ]
Hasell, M. 1978. The dynamics of arthropod predator-prey systems. Princeton, Princeton University Press., New Jersey. 237 p. [ Links ]
Heinz, K. M. y M.P. Parrella. 1990. The influence of host size on sex ratios in the parasitoid Diglyphus begini (Hymenoptera: Eulophidae) Leafminer, male-biased sex ratio, differential mortality, Liriomyza trifolii, Diglyphus begini. Ecol. Entomol. 18, 391-399. [ Links ]
Hofsvang, T.; Hagvar, E. 1983. Functional response to prey density of Ephedrus cerasicola (Hym:Aphidiidae), an aphidiid parastoid of Myzus persicae (Hom: Aphididae). Entomophaga 28(4), 317-324. [ Links ]
Holling, C.S. 1959. The components of predation as revealed by a study of small-mammal predation of the European pine sawfly. Canadian Entomologist 91, 293-320 Huffaker, C. B. y A.P. Gutiérrez. 1999. Ecological entomology. Second Edition. John Wiley & Sons, New York. [ Links ] [ Links ]
Kfir, R. y .F. Luck. 1979. Effects of constant and variable temperature extremes on sex ratio and progeny production by Aphytis melinus and A. lingnanensis (Hymenoptera: Aphelinidae). Ecological Entomology 4, 335-344. [ Links ]
Marsh, P.; S. Shaw y R. Wharton. 1987. An identification manual for the North American genera of the family Braconidae (Hymenoptera). Memoirs of the Entomological Society of Washington Nº13. The Entomological Society of Washington. Washington. 98 p. [ Links ]
Menon, A.; P.W. Flinn y B.A. Dover. 2002. Influence of temperature on the functional response of Anisopteromalus calandrae (Hymenoptera: Pteromalidae), a parasitoid of Rhyzopertha dominica (Coleoptera: Bostrichidae). J. Stored Products Res. 27, 463- 469. [ Links ]
Rogers, D.J. 1972. Random search and insect population models. J. Animal Ecol. 43, 368-383. [ Links ]
SAS Institute Inc., 1999. SAS Users Guide. Version 8. [ Links ]
Schenk, D. y S. Bacher. 2002. Functional response of a generalist insect predator to one of its prey species in the field. J. Animal Ecol. 71, 524-531. [ Links ]
Schlinger, E. y J. Hall. 1960. The biology, behavior, and morphology of Praon palitans Muesebeck, an internal parasite of the spotted alfalfa aphid, Therioaphis maculata (Bukton) (Hymenoptera:Braconidae, Aphidiinae). Ann. Entomol. Soc. Amer. 53, 144-161. [ Links ]
Schreiber, S.J. y A.P. Gutierrez. 1998. A supply/demand perspective of species invasions and coexistence: applications to biological control. Ecol. Modelling 106(1), 24 - 45. [ Links ]
Sigsgaard, L. 2000. The temperature dependent duration of development and parastism of three cereal aphid parasitoids, Aphidius ervi, A. rhopalosiphi, and Praon volucre. Entomologia Experimentalis et Applicatta 36, 173-184. [ Links ]
Solomon, M.E. 1949. The natural control of animal populations. J. Animal Ecol. 18, 1-35. [ Links ]
Waage, J. 1990. Ecological theory and the selection of biological control agents. En: Mackauer, M., L.E. Ehler, y J. Roland (eds.). Critical issues in biological control. Intercept, Andover, pp. 135-157. [ Links ]
Wang, B. y D. Ferro. 1998. Functional responses of Trichogramma ostriniae (Hym: Trichogrammatidae) to Ostrinia nubialis (Lep: Pyralidae) under laboratory and field conditions. Environ. Entomol. 27(3), 752-758. [ Links ]

