










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.25 n.2 Bogotá jul./dez. 2007
FITOMEJORAMIENTO, RECURSOS GENÉTICOS Y BIOLOGÍA MOLECULAR
Luz Nayibe Garzón1, Matthew W. Blair2 y Gustavo A. Ligarreto3
1. Programa de Doctorado en Ciencias Agrarias, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. lgarzong@unal.edu.co
2 Investigador, International Center for Tropical Agriculture (CIAT), Cali. m.blair@cgiar.org
3 Profesor asociado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.galigarretom@unal.edu.co
Fecha de recepción: mayo 23 de 2007. Aceptado para publicación: octubre 1 de 2007
RESUMEN
La antracnosis causada por Colletotrichum lindemuthianum puede llegar a generar pérdidas hasta del 95% de la producción en fríjol (Phaseolus vulgaris L.), por eso es considerada una de las enfermedades más limitantes. Con el presente trabajo se inició un programa de mejoramiento utilizando selección asistida por marcadores. Se usó como parental resistente el cultivar Mesoamericano G2333, para incorporar a fríjoles volubles comerciales de Colombia los genes Co-5 y Co-42, que confieren resistencia a C. lindemuthianum. Se obtuvieron progenies RC1F1 provenientes de cruces entre G2333 y siete parentales volubles tipo Bola roja y Rojo moteado cultivados en zonas productoras de fríjol de Cundinamarca y Boyacá. La selección de plantas RC1F1 se realizó con los marcadores moleculares SCAR SAB3 y SAS13, ligados a los genes Co-5 y Co-42, respectivamente. En la evaluación genotípica de 1.271 plantas RC1F1, 608 amplificaron con SAB3 y 603 con SAS13. La segregación para cada marcador se ajustó a la razón 1:1 esperada en pruebas de chi-cuadrado (X2 = 2,38, P = 0,12 para SAB3 y X2 = 3,32, P = 0,07 para SAS13). Un total de 299 plantas RC1F1 amplificaron con ambos SCAR, ajustándose a la razón esperada (X2 = 1,11, P = 0,78) y confirmando la segregación independiente. Estas plantas fueron seleccionadas para continuar el programa de mejoramiento, ya que se espera que porten los genes Co-5 y Co-42. Se logró implementar la selección asistida por marcadores para antracnosis, y acelerar la introducción y piramidación de genes de resistencia en fríjol de importancia económica en Colombia.
Palabras clave: Colletotrichum lindemuthianum, fitomejoramiento, genes Co-42 y Co-5, SAB3, SAS13.
ABSTRACT
The anthracnose caused by Colletotrichum lindemuthianum is one of the most significant diseases in beans (Phaseolus vulgaris L.) and can produce yield losses up to 95%. In this study, a breeding program utilizing molecular marker assisted selection (MAS) was initiated. In order to obtain commercial Colombian climbing beans resistant to C. lindemuthianum, the program incorporated genes Co-5 and Co-42taken from G2333, a resistant Mesoamerican cultivar. From the crosses between G2333 and seven cultivated Andean beans, were obtained progenies RC1F1. The cultivated Andean beans included Bola roja and Rojo moteado types gathered from Cundinamarca and Boyacá crops. The BC1F1 plants were selected using the SCARs molecular markers SAB3 and SAS13 linked to Co-5 and Co-42 genes, respectively. In a genotypic evaluation of 1.271 RC1F1 plants, 608 and 603 were amplified with SAB3 and SAS13, respectively. The segregation expected ratio (1:1) in X2 tests was obtained for each molecular marker indicating X2 = 2,38, P = 0,12 for SAB and X2 = 33,32, P = 0,07 for SAS13. The amplification with both molecular markers was obtained in 299 plants BC1F1, according to the expected ratio (X2 = 1,11, P = 0,78) adjusted to the independent segregation. These plants were selected to continue the breeding program. In this study, a molecular marker assisted selection was implemented and introduction of resistant genes into commercially important Colombian beans was accelerated.
Key words: Colletotrichum lindemuthianum, breeding, genes Co-42 and Co-5, SAB3, SAS13.
Introducción
El fríjol es una de las leguminosas más importantes en el mundo (Broughton et al., 2003). En Colombia tiene gran impacto por su volumen de producción, valor alimenticio, generación de empleo y amplia aceptación en la dieta popular con 3,86 kg de consumo por persona al año (Mincomex, 2003). En 2001, el fríjol generó un ingreso bruto aproximado para los agricultores colombianos de US$ 98´922.935, además de 12,5 millones de jornales directos, en los que participaron cerca de 125.000 productores (Mincomex, 2003). Colombia, a pesar de ser un país con variabilidad genética en fríjol, con cultura de producción y adaptación del cultivo a variados ambientes, presenta una reducción y déficit en la oferta para suplir el mercado interno, por esta razón tiene que realizar importaciones para poder cubrir la demanda nacional (Mincomex, 2003). Una de las causas principales que ha llevado a la reducción del área cultivada y disminución en la producción es la ocurrencia de enfermedades, de las cuales la antracnosis, ocasionada por el hongo C. lindemuthianum, es considerada una de las más limitantes (Pastor-Corrales, 1988). La gran variabilidad del patógeno en Colombia quedó demostrada por diferentes investigaciones que evaluaron 236 aislamientos de C. lindemuthianum provenientes de Antioquia, Caldas, Cauca, Cundinamarca, Nariño, Santander y Valle, en los que se identificaron 61 razas diferentes (Otoya et al., 1995; Santana y Mahuku, 2002; Jara et al., 2004). Para el control de la antracnosis se han diseñado diversas estrategias de manejo como el físico, cultural y químico; sin embargo, la resistencia varietal se considera la fuente de control más eficiente y económica, pues contribuye a la conservación del medio y reduce la aplicación de agroquímicos (Balardin y Kelly 1998; Beebe y Pastor-Corrales, 1991; Castanheira et al., 1999; Silva y Santos, 2001).
La resistencia a antracnosis en fríjol está condicionada principalmente por diez genes mayores independientes, numerados de Co-1 a Co-11 (Kelly y Vallejo, 2004; Goncalves- Vidigal et al., 2007). El gen Co-11 es el último gen descrito y fue reportado por Goncalves-Vidigal et al. (2007). Con excepción de Co-7 y co-8, todos los loci descritos han sido mapeados en diferentes grupos de ligamiento. La independencia de estos genes distribuidos a lo largo del genoma, ofrece a los mejoradores la oportunidad de piramidar genes complementarios y, con base en su espectro de resistencia, escoger los alelos más efectivos para obtener una resistencia durable (Kelly y Vallejo, 2004).
El cultivar diferencial G2333 constituye un ejemplo de la ventaja de que un genotipo posea más de un gen para el control de la antracnosis en fríjol (Young y Kelly, 1996b). G2333 pertenece al acervo mesoamericano y tiene una semilla de color rojo, pequeña, con hábito de crecimiento indeterminado tipo IV (Pastor-Corrales et al., 1994). Pastor-Corrales et al. (1994) determinaron que la resistencia en G2333 era controlada por los genes Co-4 y Co-7. Balardin et al. (1997) describieron a uno de los genes de G2333 como Co-42, reportando este alelo como uno de los más efectivos frente a razas de C. lindemuthianum con un índice de resistencia del 97%. Young y Kelly (1996a) mostraron que G2333 también portaba el gen Co-5, el cual no fue observado por Pastor-Corrales et al. (1994), porque utilizaron la raza 521, virulenta frente a este gen. Los múltiples genes en G2333 hacen de él un parental apropiado como fuente de resistencia para obtener variedades mejoradas, especialmente para los fríjoles andinos volubles, ya que G2333 también es voluble.
La piramidación de genes es difícil de conseguir con métodos clásicos de mejoramiento debido a que los genes de resistencia pueden presentar relaciones epistáticas o penetrancia incompleta de la resistencia; el patógeno puede presentar variabilidad en agresividad y a veces no está disponible y la reacción a la enfermedad por las condiciones ambientales es variable (Kelly y Vallejo, 2004). Adicionalmente, algunas evaluaciones para enfermedad requieren mucho tiempo y solo pueden ser realizadas enalgunas zonas, épocas, condiciones agroecológicas o estados específicos del desarrollo de la planta (Adam-Blondon et al., 1994; Marcia y Joao, 2001; Michelmore, 1995; Winter y Kahl, 1995; Young y Kelly, 1996b). El uso de selección asistida por marcadores moleculares (SAM), facilita el proceso de mejoramiento y puede acelerar la generación de nuevas variedades, ya que permite la asociación de caracteres fenotípicos con los loci genómicos responsables de esta (Winter y Kahl, 1995).
Dentro de los marcadores que se han descrito por estar ligados a genes de resistencia a antracnosis en fríjol se encuentran varios de tipo SCAR (sequence characterised amplified region), dos de los cuales son específicos para los genes de G2333. El primero es SAS13, que amplifica una sola banda dominante y polimórfica con un peso de 950 pb, presente sólo en los materiales resistentes a antracnosis que portan el gen Co-42 (Young et al., 1998) y el segundo es SAB3, que amplifica una sola banda polimórfica de 450 pb asociada a la resistencia conferida por el gen Co-5 (Vallejo y Kelly, 2001). Estos marcadores han sido utilizados para el mejoramiento de fríjoles arbustivos (Alzate-Marín et al., 2004; Miklas y Kelly 2002; Miklas et al., 2003) en Brasil y Estados Unidos, pero nunca se han utilizado los marcadores y los genes en fríjoles volubles de la zona Andina.
Con este trabajo se inició un programa de mejoramiento asistido por los marcadores SAB3 y SAS13, encaminado a la obtención de variedades de fríjol voluble con resistencia a antracnosis, dirigido a las zonas productoras de Cundinamarca y Boyacá. Se obtuvieron poblaciones RC1F1 provenientes de los cruces entre G2333 como fuente de resistencia y siete variedades andinas tipo Bola roja (Cabrera, Comercial, D. Moreno, Pesca y Simijaca) y Rojo moteado (Agrario y Cargamanto rojo), que a pesar de presentar gran demanda son susceptibles a C. lindemuthianum. El fríjol Bola roja se caracteriza por pertenecer al acervo Andino, tener hábito de crecimiento voluble, semilla de color rojo y tamaño grande, adaptada a climas medios y fríos, mientras que el fríjol Moteado es similar pero tiene grano rojo con crema (Cargamanto) o rosado con rojo (Agrario) (Ligarreto, 1997; Santana y Mahuku, 2002).
Materiales y métodos
Material vegetal
El parental resistente G2333 fue suministrado por el banco de germoplasma del CIAT (Cali, Colombia). Los fríjoles volubles de las variedades Bola roja y Rojo moteado utilizados en el presente trabajo provienen de zonas productoras de Cundinamarca y Boyacá, y fueron colectados por el programa de mejora genética de fríjol de la Universidad Nacional de Colombia, Facultad de Agronomía, durante el primer semestre de 2004. Se denominaron: 1) Agrario, 2) Bola roja comercial, 3) Cabrera, 4) Cargamanto, 5) D. Moreno, 6) Simijaca, y 7) Pesca. La denominación hace referencia al lugar en el que se cultiva o al nombre local. Algunas características de los materiales utilizados se encuentran en la tabla 1.
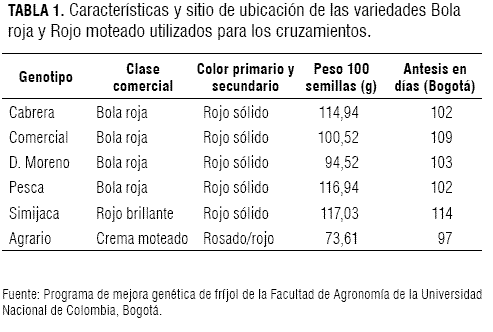
Obtención de progenies F1 y F1RC1
Para transferir los genes de resistencia del parental G2333, hacia los fríjoles volubles andinos se realizaron cruces en condiciones de campo, mediante técnicas de hibridación desarrolladas en la Universidad de Puerto Rico, que no requieren emasculación. La F1 se obtuvo realizando cruzas simples tanto directas (voluble andino x G2333) como recíprocas (G2333 x voluble andino). Los cruces se hicieron en el segundo semestre de 2004 en los viveros de cruzas de las estaciones experimentales del CIAT, ubicadas en Popayán (Cauca, 1.737 msnm, temperatura promedio: 19,8 ºC, precipitación: 1.941 mm) y Darién (Valle del Cauca, 1.485 msnm, temperatura promedio: 18 ºC) y en el invernadero del proyecto de leguminosas de la Universidad Nacional de Colombia, sede Bogotá (2.650 msnm, temperatura promedio: 14,5 ºC). Cada progenie se sembró en dos surcos de 3 m, 30 semillas por cada surco, para un total de 60, la distancia entre surcos fue de 1 m. Para coincidir en floración, G2333 tuvo dos épocas de siembra diferentes: 15 días antes y el mismo día que la siembra de las variedades comerciales.
La semilla F1RC1fue obtenida por retrocruzamientos entre la F1 con cada una de las variedades comerciales como parentales recurrentes. En este caso también se hicieron cruzas recíprocas, tomando la F1 como parental madre o padre. Se utilizó una tercera parte de la semilla F1 para el vivero de retrocruzas sembrado en febrero de 2005 en el invernadero de Bogotá, otro tercio para la estación experimental del CIAT en Darién sembrada en el mes de marzo y el resto se guardó como reserva. Cada una de las variedades comerciales tuvo dos épocas de siembra: 15 días antes y el mismo día de la siembra de la semilla F1, para asegurar que las épocas de floración coincidieran y se pudieran realizar los retrocruzamientos correspondientes. El tamaño de las parcelas varió dependiendo de la disponibilidad de semilla para cada cruzamiento, la distancia entre plantas fue 10 cm y entre surcos 1 m. En los meses de mayo y junio se presentó la floración tanto de los parentales como de las progenies F1 y de esta manera se realizaron los retrocruzamientos hacia cada una de las variedades comerciales.
Selección asistida por marcadores moleculares (SAM)
La semilla RC1F1 fue sembrada en el invernadero de la Universidad Nacional de Colombia, sede Bogotá, en agosto de 2005. En promedio, se sembró un 69% de cada uno de los retrocruzamientos y el resto se guardó como reserva. Por esta razón, el tamaño de las parcelas varió de acuerdo con la cantidad de semilla para cada progenie. La distancia entre plantas por disponibilidad de espacio fue de 20 cm. A los veinte días se marcó cada planta con una etiqueta que identificaba la genealogía correspondiente y se realizó la cosecha del material vegetal para iniciar el proceso de SAM.
Extracción de ADN y amplificación con SCAR
El ADN necesario para la amplificación con los SCAR se obtuvo siguiendo el protocolo de la extracción alcalina, una metodología rápida y económica propuesta por Klimyuk et al. (1993), que permite manejar un gran número de muestras en tiempos cortos. Usando un folíolo joven con un sacabocados se tomó un disco de tejido de 6 mm de diámetro y con unas pinzas se transfirió a una placa ELISA de fondo redondo (Corning). Cada vez que se tomaba una muestra nueva tanto el sacabocados como la pinza fueron esterilizados con etanol al 70%, este procedimiento se realizó hasta completar los 96 pozos de la placa. Como control positivo se colocó un disco de G2333 cada 30 muestras y en el siguiente pozo una muestra del parental susceptible de la cruza que se estuviera cosechando. Las placas con el material vegetal se mantuvieron en neveras de icopor con hielo y se llevaron al laboratorio donde fueron procesadas. Para hacer más eficiente la extracción, los reactivos se aplicaron utilizando una pipeta multicanal (5-50 μL). A cada muestra se le agregó 40 μL de NaOH 0,25 M y la placa se incubó en baño María a 100 oC durante 2 minutos, el tejido se maceró y se le agregó 40 μL de HCl 0,25 M. Inmediatamente después se neutralizó adicionando 20 μL de Tris-HCL 0,5 M pH 8,0, se llevó nuevamente a baño María 100 oC durante 2 minutos. Finalmente, se dejó enfriar y con el extracto obtenido se hizo una dilución 1:20 (5 μL del extracto de ADN:95 μL de agua).
Amplificación por PCR
Cada reacción de amplificación por PCR, con un volumen final de 15 μL, contenía 5 μL de ADN diluido; 1X de buffer PCR (500 mM de KCl, 10 mM de Tris HCl pH 8,8, 1% de Triton X-100, 1 mg/mL albúmina sérica bovina); 0,10 μM de cada cebador (Invitrogen Corp. San Diego, California), 0,25 mM de cada dNTP, 2,5mM de MgCl2, 1 unidad de Taq y agua de calidad HPLC para completar al volumen final. Las amplificaciones de PCR se hicieron en placas de 96 pozos selladas con tapas en silicona y utilizando un termociclador PTC200 (MJ Research). A cada producto de PCR amplificado se le adicionó 2 μL de buffer carga (glicerol 30%; azul de bromofenol 0,25%) y se analizó en cámara de electroforesis horizontal (FB-SB-2025), utilizando geles de agarosa al 1,5%, teñidos con 0,5 μL/mL de bromuro de etidio. Los geles se colocaron en buffer TBE 0.5X (90mM de Tris-borato, 2 mM de EDTA, pH 8,0) y se corrieron a 200-220 V durante 30 minutos. Las bandas se visualizaron bajo luz ultravioleta y fueron fotografiadas con cámara Kodak DC290.
Con los datos de amplificación obtenidos para cada marcador se construyó una matriz de ceros y unos, que correspondían a ausencia y presencia de la banda, respectivamente. Con la sumatoria de esta información representando la segregación global y dentro de cada pedigree se realizaron pruebas X2 de ajuste para determinar si las segregaciones obtenidas correspondían a las esperadas 1:1, 1:3 o 1:15.
Resultados y discusión
En el presente trabajo se realizaron en total 1.214 cruces simples entre G2333 y las variedades Agrario (225), Comercial (149), Cabrera (168), Cargamanto (196), D. Moreno (190), Pesca (132) y Simijaca (159) (tabla 2). En los tres viveros de cruzas se cosechó un total de 589 vainas, esto correspondió a un 49% de eficiencia en el proceso de hibridación variando entre 9 y 85% entre cruzas y 37 y 65% entre localidades. La eficiencia es alta considerando que no se usó emasculación y que CIAT (1982) reporta que la eficiencia de la hibridación con emasculación es a veces baja, con valores entre el 35 y 50 %. En el caso de Darién se observó un aumento en este porcentaje, comparado con los otros ambientes, alcanzando un 65%. Esto puede deberse al ambiente más favorable de Darién para uno de los parentales específicamente G2333. Se obtuvo un excesode semilla híbrida de cruzas simples para poder distribuirla en las tres localidades (Bogotá, Darién y Popayán), realizar un gran número de retrocruzamientos con los parentales comerciales y guardar un remanente.
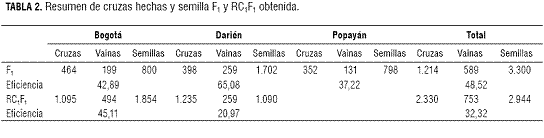
Aparte de la efectividad de cruzamientos, también hubo variación en el número de semillas producido por vainas. De los cruzamientos simples recíprocos hechos en todo el estudio se cosecharon 589 vainas con un total de 3.300 semillas F1. La distribución de producción en los tres ambientes se muestra en tabla 2. Se observó una diferencia considerable entre la cantidad de semilla F1 obtenida cuando G2333 se tomó como parental padre (1.080 semillas) y cuando G2333 fue parental madre (622 semillas). Estos valores muestran un aumento considerable del 42% cuando la polinización se efectuó con G2333 como madre. Además, G2333 produjo en promedio de 7 a 8 semillas por vaina, que explica el alto número de semilla F1 obtenida de las cruzas con este parental.
El éxito del proceso de hibridación sin emasculación usado en este estudio, al parecer, se debió a que al obviar la emasculación de la flor, se maltrató menos la flor, quedando las estructuras reproductivas protegidas por los pétalos y la quilla. Adicionalmente, el tiempo de manipulación fue más corto, factores que finalmente incidieron en la obtención de un porcentaje mayor de híbridos y en la disminución del aborto o la caída de botones en los cuales se realizó el cruzamiento.
Antes de realizar los retrocruzamientos, se verificó que las plantas fueran híbridas y se descartaron las autofecundadas. Para esto se tuvieron en cuenta las aracterísticas más contrastantes entre las plantas F1 versus las plantas autofecundadas. Estas características incluyeron color de flor, precocidad de maduración, presencia de estrías en las vainas y presencia de hilium. Se determinó que de las semillas F1 sembradas hubo en total 5% de autofecundaciones. Este valor es bajo y permite concluir que se puede trabajar con la semilla obtenida de cruzamientos sin emascular y que la probabilidad de polinizaciones efectivas es de 95%. Estos resultados corroboran la eficiencia del cruzamiento sin emasculación, que puede ser implementado de manera confiable en los programas de mejoramiento de fríjol aumentando la eficiencia, ya que esta técnica disminuye el tiempo necesario para realizar cada cruzamiento, por lo que se puede hacer un mayor número en menor tiempo.
Al realizar los retrocruzamientos hacia los parentales Bola roja y Rojo moteado, de 2.330 cruzamientos se cosecharon 753 vainas con un total de 2.944 semillas (tabla 2). En Darién, aunque se realizaron 1.235 retrocruzamientos, las vainas híbridas cosechadas fueron 259, esto corresponde a una eficiencia del 21%, la tercera parte de la eficiencia obtenida cuando se hicieron los cruzamientos simples. Los fríjoles comerciales fueron utilizados como parentales recurrentes, equitativamente como padres o madres. Las variedades comerciales al proceder de Cundinamarca y Boyacá estuvieron menos adaptadas a las condiciones ambientales de Darién, y esto pudo haber explicado la baja eficiencia. Este mismo argumento explica por qué se mantuvo la eficiencia en hibridación con los cruzamientos realizados en Bogotá donde se alcanzó un 45%. La buena cantidad de semillas F1 obtenidas (3.300), permitió hacer los retrocruzamientos con un buen número de plantas, realizar hibridaciones en varios lugares y, así generar el gran número de semillas RC1F1 (2.944), cantidad sobresaliente con la que se logra mantener la variabilidad presente en los parentales, y aumentar la probabilidad de encontrar segregantes o recombinantes favorables (Singh, 1998; Vallejo y Estrada, 2002). El hecho de haber obtenido una buena cantidad de semilla RC1F1, permite contar con una amplia base genética para continuar el proceso de mejoramiento basado en evaluaciones fenotípicas y moleculares que se describen a continuación.
Selección asistida con los SCAR SAB3 y SAS13
Debido a que en SAM el número de muestras que se evalúa es grande, es necesario utilizar un tipo de extracción que permita manejar con confiabilidad gran cantidad de muestras en un tiempo corto. La extracción alcalina propuesta por Klimyuk et al. (1993), utilizada en el presente trabajo con algunas modificaciones, permitió obtener el ADN de las plantas RC1F1 con suficiente calidad para una buena amplificación con los SCAR. De esta manera se logró disminuir el tiempo, los reactivos y el costo de selección. En total se realizó evaluación de marcadores en 1.355 muestras, discriminadas en 1.271 plantas RC1F1, 42 controles negativos y 42 positivos.
El SCAR SAB3 amplificó una banda de 450 pb, como fue reportado por Vallejo y Kelly (2001) (figura 1). Con SAS13 la banda amplificada fue de 950 pb, correspondiendo a la reportada por Young et al. (1998). En algunas muestras se observó la amplificación de una banda con aproximadamente 250 pb. Esta banda inespecífica también fue reportada por Melotto et al. (2004) y lo atribuyen a secuencias repetitivas; sin embargo, la banda inespecífica no está asociada a resistencia o susceptibilidad, por lo que no se tuvo en cuenta para la lectura de los geles para la selección.
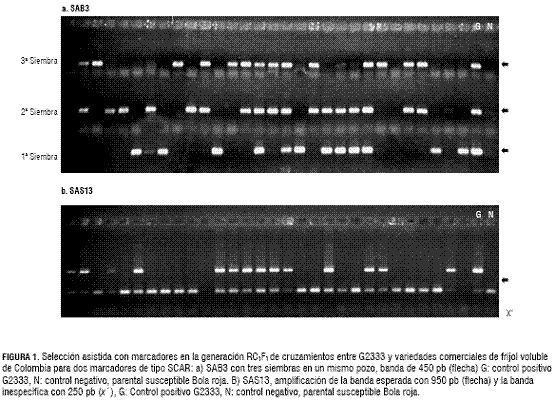
En general, la segregación de los SCAR de manera individual y conjunta se ajustó a las razones esperadas para cada retrocruza evaluada (tabla 3). Para el total de las 1.271 muestras RC1F1 evaluadas, en 608 se observó la banda amplificada con SAB3 y 663 no tuvieron la banda, ajustándose a la segregación esperada 1:1 (X2 = 2,38, P = 0,12). Esta segregación esperada de 1:1 también se obtuvó con SAS13, donde 603 plantas amplificaron la banda y 668 no lo hicieron (X2 = 3,32 y P = 0,07).
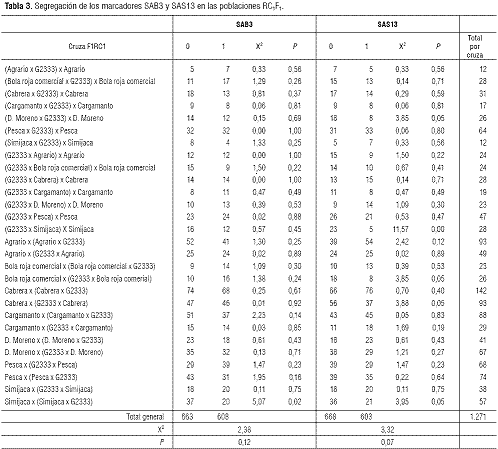
La combinación de los SCAR SAB3 y SAS13 permitió seleccionar 299 plantas RC1F1 que amplificaron con ambos marcadores (esta cantidad se ajusta a lo esperado para segregación digénica y se confirma con el valor X2 = 1,11 (P = 0,78). Estas plantas fueron seleccionadas por la probabilidad de tener los dos genes deseados, Co-42 y Co-5. La presencia y complementariedad de estos dos genes confiere un espectro de resistencia amplio frente a C. lindemuthianum (Alzate-Marín et al., 2004; Campa et al., 2005; Kelly y Vallejo, 2004).
Sin embargo, algunas consideraciones deben tenerse en cuenta con el background de los materiales seleccionados mediante la evaluación con los marcadores, dentro de ellos el hecho de que el SCAR SAB3 se encuentra a 12,98 cM del gen Co-5 y se pueden presentar eventos de recombinación que generen falsos positivos (Vallejo y Kelly, 2001). Adicionalmente, aunque G2333 ha sido utilizado de manera exitosa para generar materiales mejorados con resistencia (Miklas y Kelly, 2002; Alzate-Marín et al., 2004), los reportes hechos por Ernest y Kelly (2004) enfatizan la necesidad de realizar inoculaciones en alguna etapa del proceso de mejoramiento para corroborar la evaluación genotípica, puesto que el uso de Co-4 puede estar limitado a ciertas regiones o grupos varietales. Es importante tener en cuenta que el uso de los marcadores moleculares es una herramienta que ayuda a acelerar el proceso de piramidación de genes, pero no reemplaza las técnicas de mejoramiento convencional y además es necesario continuar explorando la complementariedad entre el mejoramiento clásico y el asistido con herramientas biotecnológicas. La presente investigación se convierte en el primer trabajo en Colombia que reporta el uso de SAM para complementar el proyecto de mejoramiento encaminado a la búsqueda de materiales de fríjol Andino con resistencia a C. lindemuthianum.
La eficiencia de la selección indirecta de plantas resistentes para antracnosis se ha reportado en trabajos como el de Miklas y Kelly (2002), quienes lograron ingresar el gen deresistencia Co-42 en fríjoles cultivados arbustivos “pinto” también usando el marcador SAS13, y liberar una línea resistente registrada como USPT-ANT-1. Adicionalmente, Alzate-Marín et al. (2004) realizaron la selección con RAPD de la cual se derivaron los SCAR SAB3 y SAS13 (OPABO3 ligado al gen Co-5 y OPAS13 ligado al gen Co-4) y lograron desarrollar líneas de fríjol arbustivo también, pero tipo “carioca” resistentes. El objetivo a largo-plazo de esta investigación es obtener segregantes de fríjoles volubles de tipo Bola roja, Agrario y Cargamanto similares a los parentales comerciales usados en este estudio, pero con resistencia a C. lindemuthianum conferida por los genes Co-42 y Co-5 transferidos desde el cultivar G2333. Se espera que estas líneas complementen variedades liberadas anteriormente como las LAS109 para el departamento de Antioquia y dos de tipo mortiño para Nariño que tuvieron resistencia a antracnosis pero que no tuvieron aceptación comercial (Ligarreto, 1997).
Agradecimientos
Los autores agradecen al Instituto Colombiano para el Desarrollo de la Ciencia y Tecnología Francisco José de Caldas (Colciencias), por la financiación para la realización de este trabajo, en el marco del proyecto “Obtención de nuevas variedades de fríjol común con atributos de rendimiento y potencial para nuevos mercados, utilizando selección convencional y asistida por marcadores moleculares”. A los integrantes de la línea de investigación en leguminosas de la Universidad Nacional y del CIAT por su apoyo, a Alcides Hincapié, César Cajiao y Jesús Cuesto por su colaboración en campo, a Linda Rincón por su asesoría en fitopatología, al profesor David Cuéllar por su valiosa ayuda y a Héctor Buendía, Teresa Mosquera y Óscar Oliveros por el apoyo en laboratorio.
Literatura citada
Adam-Blondon, A.F., M. Sévignac, H. Bannerot y M. Dron. 1994. SCAR, RAPD and RFLP markers linked to a dominant gene (Are) conferring resistance to anthracnose in common bean. Theor. Appl. Genet. 88, 865-870. [ Links ]
Alzate-Marín, A.L., K.M. Arruda, K.A. Souza, E.G. Barros y M.A. Moreira. 2004. Introgression of Co-42 and Co-5 anthracnose resistance genes into "Carioca" Common bean cultivars. Crop Breeding Appl. Biotechnol. 4, 446:451. [ Links ]
Balardin, R.S., J.S. Smith y J.D. Kelly. 1997. Virulence and molecular diversity in Colletotrichum lindemuthianum, from South, Central and North America. Phytopathol. 87, 1184-1191. [ Links ]
Balardin, R.S. y J.D. Kelly. 1998. Interaction among races of Colletotrichum lindemuthianum and diversity in Phaseolus vulgaris. J. Amer. Soc. Hort. Sci. 123, 1038-1047. [ Links ]
Beebe, S.E. y M. Pastor-Corrales. 1991. Breeding for disease resistance. pp. 561-617. En: Schoonhoven, A. van y O. Voysest (eds.). Common beans: Research for crop improvement. CAB Internacional, Oxon, UK. [ Links ]
Broughton, W.J., G. Hernández, M. Blair, S. Beebe, P. Gepts y J. Vanderleyden. 2003. Bean (Phaseolus spp.) model food legumes. Plant Soil 252, 55-128. [ Links ]
Campa, A., C. Rodríguez-Suárez, A. Pañeda, R. Giradles y J.J. Ferreira. 2005. The bean anthracnose resistance gene Co-5, is located in linkage group B7. Annu. Rep. Bean Improvement Coop. 48, 68-69. [ Links ]
Castanheira, A.L.M., J.B. Santos, D.F. Ferreira y L.C. Melo. 1999. Identification of common bean alleles resistant to anthracnose using RAPD. Genet. Mol. Biol. 22(4), 519-524. [ Links ]
CIAT. 1982. Cruzamiento de fríjol. Guía de estudio para ser usado como complemento de la unidad audotutorial sobre el mismo tema. CIAT, Cali. 36 p. [ Links ]
Ernest, E.G. y J.D. Kelly. 2004. The Mesoamerican anthracnose resistance gene,Co-42, does not confer resistance in certain Andean genetic backgrounds. Annu. Rep. Bean Improvement Coop. 47, 245-246. [ Links ]
Gonçalves-Vidigal, M.C., C.R. Silva, F.P.S. Vidigal, A. Gonela y M.V. Kvitschal. 2007. Allelic relationships of anthracnose (Colletotrichun lindemuthianum) resistance in the common bean (Phaseolus vulgaris L.)cultivar Michelite and the proposal of a new anthracnose resistance gene, Co-11. Genet. Mol. Biol. 30(3), 589-593 [ Links ]
Jara, C., J. Fory, G. Castellanos, J.B. Cuasquer y G. Mahuku. 2004. Pathogenic characterization of Colletotrichum lindemuthianum isolates from different regions of Colombia. Annu. Rep. Bean Improvement Coop. 47, 160-161. [ Links ]
Kelly, J.D. y V.A. Vallejo, 2004. A comprensive review of the major genes conditioning resistance to anthracnose in common bean. HortScience 39(6), 1196-1207. [ Links ]
Klimyuk, V.I., B.J. Carroll, C.M. Thomas y J.D.G. Jones 1993. Alkali treatment for rapid preparation of plant material for reliable PCR analysis. Plant J. 3, 493-494. [ Links ]
Ligarreto, G. 1997. Variedades y mejoramiento de fríjol. Fenalce -Sena-SAC, Bogotá. 14 p. [ Links ]
Marcia, V.S. y B.S. João. 2001. Identificaçao de marcador RAPD ligado ao alelo Co-42 de resistencia do feijoeiro comun ao agente causal da antracnose. Ciencias Agrotecnicas, 25(5), 1097-1104. [ Links ]
Melotto M., M.F. Coelho, A. Pedrosa-Harand, J.D. Kelly y L.E.A. Camargo. 2004. The anthracnose resistance locus Co-4 of common bean is located on chromosome 3 and contains putative disease resistance-related genes. Theor. Appl. Genet. 109, 690-699. [ Links ]
Michelmore, R.W. 1995. Molecular approaches to manipulation of disease resistance genes. Annu. Rev. Phytopatol. 15, 393-427. [ Links ]
Miklas, P.N. y J.D. Kelly. 2002. The use of MAS to develop into bean germplasm possessing Co-42 gene for anthracnose resistance. Annu. Rep. Bean Improvement Coop. 45, 68-69. [ Links ]
Miklas, P.N., J.D. Kelly y S.P. Singh. 2003. Registration of anthracnose resistant pinto bean germplasm line USPT-ANT-1. Crop Sci. 43, 1889-1890. [ Links ]
Mincomex. 2003. Estudio de mercado de fríjol en Colombia. Subdirección de Inteligencia de Mercados, Bogotá. 10 p. [ Links ]
Otoya, M.M., S. Restrepo y M. Pastor-Corrales. 1995. Amplificación al azar del ADN polimórfico para evaluar la diversidad genética de Colletotrichum lindemuthianum en Colombia. Fitopatología Colombiana 19, 7-14. [ Links ]
Pastor-Corrales, M.A. 1988. La antracnosis del fríjol común, Phaseolus vulgaris, en América Latina. Documento de trabajo No. 113, CIAT, Cali. [ Links ]
Pastor-Corrales, M.A., O.A. Erazo, E.I. Estrada y S.P. Singh. 1994. Inheritance of anthracnose resistance in common bean accession G2333. Plant Dis. 78, 959-962. [ Links ]
Santana, G.E. y G. Mahuku. 2002. Diversidad de razas de Colletrotrichum lindemuthianum en Antioquia y evaluación de germoplasma de fríjol crema-rojo por resistencia a antracnosis. Agronomía Mesoamericana 13(2), 95-103. [ Links ]
Silva, M.V. y J.B. Santos. 2001. Identificação de marcador RAPD ligado ao alelo Co-42 de resistência do feijoeiro comum ao agente causal da antracnose. Revista Ciencia e Agrotecnologia 25, 1097-1104. [ Links ]
Singh, S.P. 1998. Uso de marcadores y selección de gametos para el mejoramiento simultáneo de caracteres múltiples de fríjol (Phaseolus vulgaris L.) para Mesoamérica y el Caribe. Agronomía Mesoamericana 9, 1-9. [ Links ]
Vallejo F.A. y E.I. Estrada. 2002. Mejoramiento genético de plantas. Universidad Nacional de Colombia, Palmira. 402 p. [ Links ]
Vallejo, V. y J.D. Kelly. 2001. Development of a SCAR marker linked to Co-5 locus in common bean. Annu. Rep. Bean Improvement Coop. 44, 121-122 [ Links ]
Winter, P. y G. Kahl. 1995. Molecular marker technologies for plant improvement. World Microbiol. Biotechnol. 11, 438-448. [ Links ]

