










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.25 n.2 Bogotá jul./dez. 2007
Estudio preliminar para el establecimiento de un protocolo de crioconservación para palma de aceite (Elaeis guineensis Jacq.)
Alba Lucía Villa1, Pablo Édgar Jiménez2, Raúl Iván Valbuena3, Silvio Bastidas4 y Víctor Manuel Núñez5
1 Investigadora Bancos de Germoplasma In Vitro, Centro de Investigación Tibaitatá, Corporación Colombiana de Investigación Agropecuaria (Corpoica),Mosquera (Cundinamarca). avilla@corpoica.org.co
2 Auxiliar de Laboratorio CBB, Centro de Investigación Tibaitatá, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Mosquera (Cundinamarca). pejo66@gmail.com
3 Investigador máster asociado, Centro de Investigación Tibaitatá, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Mosquera (Cundinamarca). valbuena@corpoica.org.co
4 Investigador máster principal, Centro de Investigación El Mira, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Tumaco (Nariño). sebastidas@corpoica.org.co
5 Investigador asociado, Centro de Investigación Tibaitatá, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Mosquera (Cundinamarca).vnunez@corpoica.org.co>
Fecha de recepción: julio 10 de 2007. Aceptado para publicación: octubre 1 de 2007
Resumen
El objetivo de esta investigación fue desarrollar e implementar un protocolo de crioconservación para embriones cigóticos de palma de aceite (Elaeis guineensis Jacq.), como alternativa para complementar la conservación de genotipos de palma ante las dificultades que se presentan en la colección de campo en la actualidad. Se evaluaron las etapas de pre y poscongelación de embriones cigóticos con la metodología de desecación en silica gel a diferentes tiempos de exposición (2, 2,5 y 3 h), seguido de un congelamiento rápido y una etapa de recuperación en un medio de cultivo Murashige y Skoog (1962). Todos los tratamientos de crioconservación fueron evaluados con un fotoperíodo de 16 h luz y 8 h de oscuridad, con una temperatura media de 28 ºC. Previamente se determinó el estado de desarrollo adecuado de los embriones para crioconservar. Además, se realizó la prueba de viabilidad con Tetrazolio para embriones desnudos como un control inicial del lote de semilla solicitado. Se logró establecer un protocolo de crioconservación a partir de embriones cigóticos desnudos de palma, el cual permitió obtener plántulas de embriones congelados en nitrógeno líquido (-196 ºC); alcanzando una sobrevivencia del 84,37% para embriones crioconservados. En cuanto a la diferenciación de estructuras de los embriones crioconservados, el tratamiento de dos horas obtuvo valores para plúmula del 88,9%, radícula del 55,57% y haustorio del 66,7%. Esta metodología permitiría conservar material valioso a largo plazo como copia de seguridad complementaria de la colección de campo y reduciría los costos de mantenimiento y conservación.
Palabras clave: viabilidad, conservación, embriones cigóticos, desecación.
Abstract
The objective of this research was to develop and implement a cryoconservation protocol for zygotic embryos of oil palm (Elaeis guineensis Jacq.) as a complement alternative to the conservation of oil palm genotypes due to the difficulties that occur in the field collection. Pre- and post-freezing steps of zygotic embryo cryoconservation were evaluated using the methodology of desiccation in silica gel at different exposure times (2, 2,5, and 3 h), followed by a rapid freezing and a recovery step in Murashige and Skoog (1962) culture medium. All the cryoconservation treatments were evaluated under a photoperiod of 16 h light and 8 h darkness at an average temperature of 28 ºC. The appropriate embryo development for the cryoconservation was previously determined. Additionally, the viability test with tetrazolium was applied to naked embryos as an initial control of the batch seed required. It was achieved a cryoconservation protocol for naked zygotic embryos of palm, which allowed obtaining plantlets from embryos frozen in liquid nitrogen (-196 ºC) with up to 84.37% survival. In relation to structure differentiation of the cryoconserved embryos, the two hour treatment gave values of 88.9% for plumula, 55.57% for radicle, and 66.7 % for haustorium. This methodology might allow a long-term conservation of valuable material as a complementary security procedure to copy the field collection and reduce the costs of maintenance and conservation.
Key words: viability, conservation, zygotic embryos, desiccation.
Introducción
La colección de palmáceas está conformada por 339 accesiones de palma, chontaduro y cocotero, conservada en condiciones de campo en el Centro de Investigación El Mira Tumaco (Nariño). Esta conservación en campo hace que la colección esté expuesta constantemente a factores bióticos y abióticos que pueden ocasionar una erosión genética por la pérdida de materiales. Además, en la colección existen individuos muy antiguos que tienen entre 24 y 35 años de edad y con altura de planta superior a 15 metros, lo que dificulta el manejo agronómico de la colección; por tanto, se hace imperativo desarrollar investigaciones encaminadas a establecer alternativas de conservación más eficientes para evitar su pérdida.Desde este punto de vista, la semilla de palma de aceite (Elaeis guineensis Jacq.) ha sido clasificada como recalcitrante (King y Roberts, 1979). Este tipo de semillas presentan lesiones en la deshidratación y no pueden ser almacenadas por extensos períodos (Roberts, 1973). A su vez, estas pueden ser almacenadas de dos a tres años y los embriones aislados toleran la desecación (Grout et al., 1983). Sin embargo, dada la reducción de la viabilidad a las temperaturas de almacenamiento (0 a -20 ºC) y el bajo contenido de humedad de algunos cultivares, este tipo de semillas parece mostrar un comportamiento de almacenamiento intermedio (Ellis et al., 1991). Esto podría ser considerado benéfico para la conservación de los recursos genéticos de esta especie y significa que al mantener unas condiciones óptimas se lograría un máximo de longevidad. A la especie Elaeis guineensis le ha sido asignada una alta prioridad (prioridad 3) para la conservación genética por la Internacional Board for Plant Genetic Resources (IBPGR) (Annon, 1981 y Ellis et al., 1991) ya que su germinación y almacenamiento es complicado debido a que las semillas presentan latencia y pérdida de viabilidad a corto plazo, y a que es una semilla de tipo recalcitrante (Corley, 1982).
Las investigaciones en conservación de semillas de palma de aceite se han desarrollado en principio, para establecer una metodología que permita almacenarlas por un largo tiempo sin que pierdan la viabilidad y puedan ser recuperadas. Grout et al. (1983) demostraron que es posible restablecer y germinar embriones cigóticos de palma de aceite después de ser congelados en nitrógeno líquido (-196 ºC). Las bajas temperaturas de este elemento cesan todos los procesos metabólicos y no permiten la ocurrencia de cambios genéticos durante el almacenamiento. Dumet et al. (1993), de acuerdo con los resultados de anteriores trabajos en palma de aceite, mejoraron el proceso de crioconservación de embriones somáticos, los cuales se sometieron a un proceso de precrecimiento de siete días en 0,75 M de sacarosa y el material a una desecación con aire de cámara de flujo laminar o en cajas con silica gel. En la especie Cocos nucífera se emplearon embriones cigóticos maduros (10 a 12 meses después de la polinización) de cuatro variedades que se crioconservaron y desarrollaron plántulas; estas se pretrataron con desecación con aire de cámara de flujo laminar durante cuatro horas y posteriormente se sometieron de 11 a 20 horas en un medio que contenía 600 g·L-1 de glucosa y 15% de glicerol. El promedio de recuperación estuvo entre el 33 y el 93% dependiendo de la variedad (Assy-Bah y Engelmann, 1992b).
El propósito de la presente investigación fue desarrollar e implementar un estudio preliminar que permitiera establecer un protocolo de crioconservación para embriones cigóticos de palma de aceite, a través de la metodología de desecación en silica gel con diferentes tiempos de exposición, teniendo en cuenta el estado de desarrollo de los embriones los cuales se evaluaron en las etapas de pre y poscongelación en nitrógeno líquido. El desarrollo de esta metodología permitirá conservar este valioso material a largo plazo como copia de seguridad de la colección de campo, así como también reducir los costos de mantenimiento y conservación.
Materiales y métodos
El estudio se llevó a cabo en el Programa de Recursos Genéticos y Mejoramiento Vegetal en el laboratorio del Banco de Conservación de Germoplasma In vitro del Centro de Investigación Tibaitatá, ubicado en el km 8 vía Mosquera (Cundinamarca) a 4º 71´ 28,2” latitud norte y 74º 18´ 30,5” longitud oeste.
Material vegetal
Se emplearon embriones de semillas de palma de aceite (Elaeis guineensis Jacq.) provenientes de racimos maduros del Centro Experimental El Mira Tumaco (Nariño), en donde se encuentra el Banco de Germoplasma de Palmáceas; el municipio de Tumaco se encuentra ubicado desde Punta Guascama (norte de Nariño) a 02° 37’ 20” latitud norte y 78° 24’ 20” longitud oeste hasta el río Mataje, y el límite de Colombia con Ecuador 01° 25’ 00” latitud norte y 78° 54’ 59” longitud oeste.
Tamaño de semilla
Se realizó la evaluación del tamaño de las semillas con relación a la longitud de los embriones procedentes de un muestreo estratificado de un racimo maduro, que seclasificó de acuerdo con el estado de madurez fisiológica, incluyendo periferia externa (semillas maduras), tercio intermedio (semillas medio maduras) e interior del racimo (inmaduras o verdes). Se realizaron mediciones de ancho, grosor y largo de la semilla con el fin de establecer una relación del tamaño de la semilla, y el embrión, para seleccionar el tamaño adecuado de los embriones en el proceso de crioconservación.
Pruebas de viabilidad de Tetrazolio
Con el propósito de establecer el estado fisiológico óptimo de los embriones para crioconservación, cada lote de semilla se sometió a la prueba de sal de Tetrazolio (2,3,5- Triphenyl-2HTetrazolium chloride, TTC), para determinar la viabilidad de los embriones de una forma rápida, la cual le da una coloración rojiza de los tejidos vivos del embrión mediante un proceso de reducción de la sal de TTC a formazán por la acción de las enzimas deshidrogenasas. Se siguieron instrucciones de la Internacional Seed Testing Association (Ista, 1992) para semilla de palma utilizando una concentración 0,5% con un tiempo de incubación de 24 h en oscuridad a 28 ºC, y así determinar la viabilidad de los embriones mediante verificación de la actividad respiratoria. Esta prueba permite visualizar oloraciones que van desde el rojo-púrpura, rosado y blanco. Cuando se presentan coloraciones rojas indica que el material se encuentra vivo, la coloración rosada corresponde al inicio de pérdida de viabilidad y la coloración blanca total o parcial significa que hay pérdida de viabilidad. Además, se determinó la viabilidad de los embriones desnudos al finalizar el proceso de desecación en silica gel y después del congelamiento. Los embriones sometidos a esta prueba se derivaron de todos los lotes de semilla provenientes del Centro de Investigaciones El Mira. Se generó una escala de tinción para cada una de las partes del embrión de la semilla de palma de aceite (tabla 1).

Metodología de crioconservación-desecación con silica gel
A las semillas de palma cosechadas de la colección en campo se les extrajo la almendra y fueron desinfectadas de acuerdo con el protocolo establecido para la especie,que consiste en hipoclorito de calcio al 1% + Tween-20 durante 20 min (con intercambio de la solución cada 10 min). Después se enjuagaron una vez con agua destilada estéril; posteriormente se colocaron las almendras en una solución de Ridomil al 1,5% durante 16 horas; se realizaron tres enjuagues con agua destilada estéril; se desinfectaron con hipoclorito de sodio al 0,5% + 5 gotas de Tween-20 durante 15 min y se lavaron tres veces con agua destilada estéril durante 10 min cada enjuague. El aislamiento de los embriones de las almendras se realizó con la ayuda de un bisturí. Los embriones se colocaron en cajas de Petri con papel filtro humedecido durante 24 h a 20 ºC. Luego, los embriones fueron colocados en láminas pequeñas de papel filtro en frascos con silica gel 25 g/frasco, durante 2, 2,5 y 3 h como tratamientos para secado en silica gel. La mitad de los embriones desecados se colocaron en el medio de recuperación como control del tiempo de secado y la otra mitad se colocó en criotubos de 2 mL, los cuales fueron sumergidos en nitrógeno líquido durante mínimo una hora. El descongelamiento se realizó sumergiendo los criotubos al baño de María durante tres minutos; posteriormente, la recuperación de los embriones se logró en el medio M&S (Murashige y Skoog, 1962), y finalmente se evaluaron cada 15 días. En cada uno de los procesos de desecación, congelamiento en NL y descongelamiento, se determinaron las variables porcentaje de sobrevivencia y diferenciación de embriones teniendo en cuenta la formación de la radícula, la plúmula, el haustorio y la altura de la planta.
En cada fase del proceso pre y poscongelación se utilizó un diseño experimental completamente al azar con tres repeticiones. Cada repetición constaba de una unidad experimental, conformada por una aja de Petri con cuatro embriones. El procesamiento de la información se realizó con el sistema estadístico SAS, con el que se realizaron los análisis de varianza, comparación de medias y las regresiones
lineales.
Resultados y discusión
Tamaño de semilla
Se encontraron diferencias significativas en cuanto a la fuente de variación, estado de maduración de la semilla para las tres variables en estudio. Las semillas maduras registraron los promedios más altos en cuanto a ancho (22,28 mm), grosor (19,72 mm) y largo (28,72 mm) (figura 1).Se determinó que las semillas maduras presentan embriones con longitudes mayores a 2 mm, aunque estadísticamente no existe relación entre las variables ancho, grosor y largo de la semilla, respecto a la longitud del embrión, debido a la dispersión de los datos. Esto indica que se pueden encontrar semillas maduras con diversas longitudes; así mismo, se encontró que la longitud de los embriones de estas semillas puede estar entre un rango de 2,5 a 5 mm. Este resultado se puede relacionar con la estratificación del racimo, la cual se da en la forma en que maduran las semillas desde el momento de la polinización hasta el corte del racimo. Cuando los embriones se encuentran en el rango mencionado se pueden realizar los procesos de pre y poscongelación ya que permiten una alta recuperación, debido posiblemente a que tienen un estado de desarrollo adecuado. Por tanto, las semillas procedentes de la periferia externa del racimo (maduras) son adecuadas para realizar los procesos de crioconservación, a diferencia de los embriones inmaduros con longitudes inferiores a 2 mm que presentaron problemas de diferenciación (formación de callos y sin diferenciación de estructuras como plúmula, radícula y haustorio).
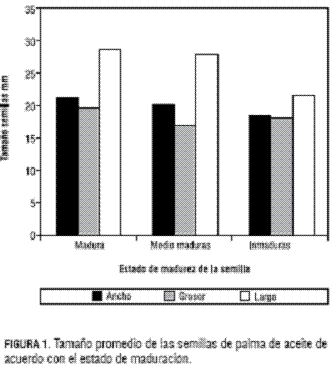
Estos resultados se relacionan con los obtenidos en embriones de la especie de Coffea arabica (Abdelnour et al., 1992), en donde el estado de madurez del embrión está relacionado con la respuesta a crioconservación. Se encontró que los embriones maduros presentan una sobrevivencia del 96%, y en inmaduros sólo del 50%, pero este último resultado lo relacionan al medio de cultivo, planteando una posible solución al adicionar en el medio de recuperación 100 mg·L-1 de AG3. En embriones de coco se confirman las apreciaciones anteriores, teniendo en cuenta que en los embriones inmaduros (siete-ocho meses después de la polinización) sólo un número limitado podría recuperarse como plántulas (Assy-Bah y Engelmann, 1992a). Caso contrario a los embriones maduros (11-12 meses después de la polinización) los cuales tienen una alta respuesta a la crioconservación (Assy-Bah y Engelmann, 1992b). Esto, posiblemente, se debe a unas condiciones inadecuadas de restablecimiento en los embriones inmaduros (Assy-Bah, 1992). También se ha encontrado que el proceso de deshidratación produce daños irreversibles en donde sólo se puede recuperar el 60% de los embriones, y el porcentaje restante muestra el desarrollo de anormalidades (callos, desarrollo de sólo el polo de la raíz o del haustorio) (Engelmann et al., 1995).
En esta investigación se encontró que cuando los embriones tienen longitudes comprendidas entre 0,5 a 1,0 mm estos no se desarrollan bien y comienzan a presentarse anormalidades, tal vez porque el estado de desarrollo no es completo al igual que cuando se presenta daño mecánico en el momento de la extracción del embrión. También como se referencia en la literatura (Engelmann et al.,1995), que la exposición de los embriones a tiempos de deshidratación prolongados causa daños en la radícula principalmente, lo que provoca una ausencia por completo y en un menor grado se presenta en el haustorio.
Desecación en silica gel y congelamiento de embriones desnudos en NL+
Para la variable porcentaje de sobrevivencia se presentaron diferencias significativas en cuanto a la desecación en silica gel, independiente del tiempo de exposición. Los embriones desecados sin congelar presentaron una sobrevivencia del 97,22%, un valor muy cercano al testigo absoluto que presentó el 100% de sobrevivencia, comparado con los embriones que se desecaron y posteriormente se congelaron, los cuales manifestaron un valor del 84,37%, mientras que el testigo absoluto congelado (sin desecar) no sobrevivió. Estos datos indican que los embriones desnudos toleran la desecación y el congelamiento con alta sobrevivencia y viabilidad. Esta tolerancia a la desecación la adquieren los embriones durante su desarrollo en la planta, a partir de los 110 días después de la polinización hasta la fase de la cosecha (Aberlenc et al. 2003). La adquisición de tolerancia a la desecación es asociada con la acumulación de materia seca y la reducción del contenido de agua. La lenta eshidratación de los embriones puede inducir la síntesis de compuestosespecíficos (ABA, LEA) proteínas o antioxidantes involucrados en la tolerancia a la desecación (Vertucci y Farrant, 1995; Pammenter y Berjak, 1999).
Se presentaron problemas de inhibición en el crecimiento de la radícula ya que el proceso de desecación en silica gel afecta la radícula y el haustorio. Por otro lado, el daño mecánico en el momento de la extracción del embrión puede causar malformación o inhibición del crecimiento de las estructuras del embrión, la plúmula, la radícula y el haustorio. En cuanto al tiempo de desecación, se encontraron diferencias del 5%, donde los mejores porcentajes de sobrevivencia estuvieron dados por los tratamientos de desecación por 2,5 horas y congelado con un 100%, desecado de tres horas y congelado 83,33% y desecado de dos horas y congelado 75% (figura 2).
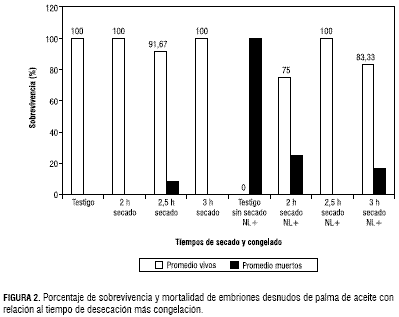
Estos resultados indican que el porcentaje de sobrevivencia es alto, tanto en la etapa de desecación como de congelación. La iferenciación de la sobrevivencia del embrión estuvo dada por el comportamiento de las estructuras del embrión de la siguiente manera: el tratamiento de tres horas, de secado más congelación presentó el promedio más alto de formación de la radícula (80,57%), superando al testigo absoluto que sólo alcanzó un 41,67%, mientras el tratamiento conformado por dos horas de desecación más congelamiento alcanzó el 33,35%. Este mismo tratamiento para formación de la plúmula obtuvo 88,90%, comparado con el testigo absoluto que presentóel 100%; para formación del haustorio este tratamiento presentó resultados satisfactorios, ubicándose en los primeros lugares con un promedio del 66,7% junto con el tratamiento de tres horas de secado más congelamiento con un 69,43% (figuras 3 y 4).
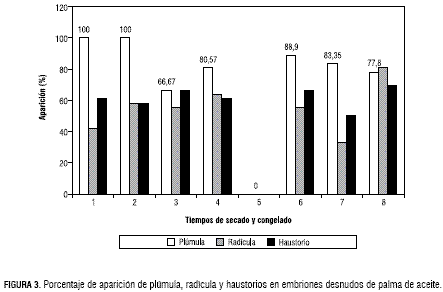

Para la variable altura, el tratamiento dos horas de desecación más congelación se mantuvo en un promedio de 10,88%, muy cerca al testigo absoluto (figura 3). En el caso de los tratamientos que entraron al proceso de congelación, independiente del tiempo de desecación, presentaron el 84,37% de sobrevivencia y la diferenciación de esta sobrevivencia estuvo dada por un 59,39% de formación de radícula, 83,35% de plúmula y 63,55% de haustorio, y una altura de planta promedio de 9,55 cm (figuras 3 y 5).

La desecación parcial de embriones cigóticos es necesaria para obtener obrevivencia después de la crioconservación; los tratamientos de desecación reducen el contenido de agua del tejido a criopreservar, aumentando de esta manera la posibilidad de sobrevivencia antes y después del congelamiento, en vista de que disminuye la formación de cristales de hielo a nivel intracelular que puede llevar a un proceso de plasmolisis o muerte celular. Esta es una ventaja en comparación con los embriones que no son desecados; sin embargo, en algunos estudios se ha encontrado que estos procesos en algunas especies como Veitchia y Howea presentan modificaciones en el patrón de reestablecimiento de los embriones crioconservados, como el no desarrollo de haustorio (Chin et al., 1988). Los resultados muestran una relación con experimentos desarrollados con embriones cigóticos de palma de aceite extraídos de semillas hidratadas los cuales revelan que el daño podría ser más severo que los observados en embriones de Vitchia, Howea o Cocos (Engelmann et al., 1995). En este estudio, el efecto de la desecación influye directamente en la formación de la radícula y del haustorio, ya que estos se encuentran en cada uno de los polos del embrión y, por tanto, se exponen directamente a la desecación.
Engelmann et al. (1995) reportan que cuando los embriones se desecan por debajo de 0,3 g H2O/g de peso seco, y son congelados en nitrógeno líquido, estos muestran un promedio alto de obrevivencia (90%). Sin embargo, la desecación induce daños irreversibles para algunos embriones y tan sólo el 60% podría desarrollarse en plántulas completas, mientras que el porcentaje restante resenta anormalidades en el desarrollo (formación de callo, sólo desarrollo en el embrión del polo de la raíz o del haustorio); además, Engelmann et al. (1995) reiteran que el buen estado fisiológico de los embriones cigóticos antes de la crioconservación, ofrece resultados satisfactorios.
Durante esta investigación se encontró que es necesario tener en cuenta el tamaño del embrión (2 a 5 mm) el cual se relaciona con el estado de desarrollo del embrión, así como el estado del mismo en razón a que en algunos casos pueden tener la longitud adecuada, pero su estado fisiológico se puede afectar, pues es un material recalcitrante. Esto hace que las condiciones medio ambientales temperatura superiores a 30 ºC) y el proceso de beneficio de la semilla (cosecha y despulpe) afecten la viabilidad de los embriones. Como consecuencia, los embriones de las semillas sufren daños severos e irreversibles (daño a nivel de radícula y haustorio por la excesiva deshidración), por consiguiente disminuye la respuesta de germinación en el medio M&S.
Prueba de viabilidad de Tetrazolio
Los resultados de esta prueba confirmaron la sobrevivencia de los embriones desnudos, ya que con 1,5 h de desecación se obtuvo el 65% de tinción de plúmula, radícula y haustorio, valor 1 de la escala (tabla 1); pero a medida que se incrementó el tiempo de desecación a dos horas, la tinción disminuyó hasta el 40% para el mismo valor 1 de la escala (tabla 1). Con 2,15 h de desecación se obtuvo un valor del 10% y con este mismo tiempo de desecación más el congelamiento en nitrógeno, la tinción fue del 2% para la misma escala; pero el 61% de los embriones procedentes de NL+ se ubicaron en el valor 3 de la escala que hace referencia a una tonalidad rosada en las estructuras del embrión (plúmula, radícula y haustorio) (figura 6).
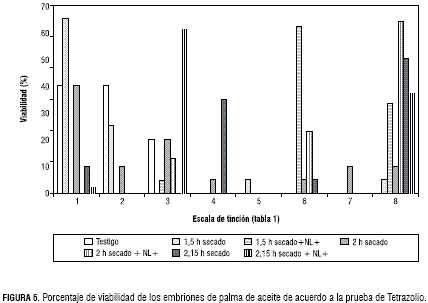
No obstante, los embriones procedentes de NL+, al colocarlos en un medio de cultivo M&S, se lograron recuperar y se obtuvieron plántulas completas, las cuales pasaron a la fase ex vitro o proceso de endurecimiento en condiciones de invernadero. Estos resultados sugieren que a pesar de que la prueba de viabilidad determinó que los embriones se afectan en el proceso de desecación y congelación, al colocarse en medio de cultivo M&S se logran recuperar, lo que implica que la técnica de cultivo de tejidos puede ser una alternativa para embriones de semillas que están perdiendo la viabilidad por estrés, longevidad o almacenamiento. En consecuencia, la prueba solo se empleó como control del material procedente de campo, ya que es un indicativo real del estado del embrión después de los procesos de cosecha y despulpe. En condiciones de campo, las semillas que pierden humedad debido a la naturaleza de la misma (recalcitrante-intermedia “sensibles a temperaturas altas”) disminuyen la viabilidad del embrión a nivel del haustorio y la radícula, para el caso del haustorio se podría pensar que estos embriones no tienen la posibilidad de recuperarse, en razón a que pierden la conexión con la almendra la cual le aporta los nutrientes necesarios para el proceso de germinación; así mismo, cuando la radícula se ve afectada, la semilla no geminará porque esta última es la que le permite anclarse para tomar del sustrato los nutrientes necesarios y así completar el desarrollo de la planta en condiciones de campo.
Conclusiones
La selección de las semillas de la periferia del racimo de palma africana (semillas maduras) permitió obtener embriones de una longitud mayor o igual a 2 mm, los cuales presentan un desarrollo adecuado y pueden soportar el proceso de desecación y congelación, dependiendo del tiempo de exposición al secado; lo que induce a un mínimo de lesiones en las de estructuras del embrión. Aunque el proceso de secado afecta especialmente a la radícula, esta puede recuperarse cuando el material es adaptado a condiciones ex vitro y la planta puede desarrollarse normalmente. Este estudio permitió establecer un protocolo de crioconservación a partir de embriones cigóticos desnudos de palma (Elaeis guineensis Jacq.), el cual consiste en colocar los embriones durante de 2 h en silica gel (20 g/frasco), llevándose luego a inmersión rápida en nitrógeno líquido (NL) durante 1 h; se descongelan al baño de María a 37ºC durante 3 min y se restablecen en un medio de cultivo M&S con una temperatura de 28 ºC, 16 h luz y 8 h de oscuridad. Este protocolo permitió obtener plántulas de embriones cigóticos congelados en nitrógeno líquido (-196 ºC); alcanzando una sobrevivencia del 84,37% para embriones desecados y congelados. En cuanto a la diferenciación de estructuras, el tratamiento de 2 h de secado más congelación directa en nitrógeno líquido presentó valores para plúmula del 88,9%, radícula del 55,57% y haustorio del 66,7%.
Agradecimientos
Los autores expresan sus agradecimientos al Ministerio de Agricultura y Desarrollo Rural por la financiación, así como a Silvio Bastidas, investigador del Centro Experimental El Mira, Corpoica Tumaco (Nariño), por el suministro tanto de semilla de palma como por la literatura de la especie, elementos necesarios para el desarrollo de esta investigación.
Literatura citada
Aberlenc-Bertossi, F., N. Chabrillange, F. Corbineau y Y. Duval 2003. Acquisition of desiccation tolerance in developing oil palm (Elaeis guineensis Jacq.) embryos in plant and in vitro in relation to sugar content. Seed Sci. Res. 13, 179-186. [ Links ]
Abdelnour, A., V. Villalobos y F. Engelmann. 1992. Cryopreservation of zygotic embryos of Coffea spp. Cryo-Letters 13, 297-302. [ Links ]
Assy-Bah, B. 1992. Utilization de la culture in vitro d´embryons zygotiques pour la collecte et la conservation des resources genétiques du cocotier (Cocos nucifera L.). Thése d´Unversité. Paris 6. 146 p. [ Links ]
Assy-Bah, B. y F. Engelmann. 1992a. Cryopreservation of immature embryos of coconut (Cocos nucifera L.). Cryo-Letters 13, 67-74. [ Links ]
Assy-Bah, B. y F. Engelmann. 1992b. Cryopreservation of mature embryos of coconut (Cocos nucifera L.) and subsequent regeneration of plantlets. Cryo-Letters 13, 117-126. [ Links ]
Chin, H.F., B. Krishnapillay y Z. Alang. 1988. Cryopreservation of Veitchia and Howea palms embryos: non-development of the haustorium. Cryo-Letters 9, 372-379. [ Links ]
Corley, R.H.V. 1982. Chapter 3: Germination and seedling growth. pp. 23-36. En: Developments in Crop Science (1): Oil Palm Research. Second edition. Scientific Publishing Company, Amsterdam. [ Links ]
Dumet, D., F. Engelmann, N. Chabrillange y Y. Duval. 1993. Cryopreservation of oil palm (Elaeis guineensis Jacq.) somatic embryos involving a desiccation step. Plant Cell Rpt. 12, 352-355. [ Links ]
Ellis, R.H., T.D. Hong, E.H. Roberts y U. Soetisna, 1991. Seed storage behaviour in Elaeis guineensis. CAB International, Wallingford. pp. 99-104. [ Links ]
Ellis, R.H., T.D. Hong y E.H. Roberts. 1991. Effect of storage temperature and moisture on the germination of papaya seeds. Seed Sci. Res. 1, 69-72. [ Links ]
Engelmann, F., D. Dumet, N. Chabrillange, A. Abdelnour-Esquivel, B. Assy-Bah, J. Dereuddre y Y. Duval. 1995. Factors affecting the cryopreservation of coffee, coconut and oil palm embryos. Plant Genet. Resources Nwsl. 103, 27-31. [ Links ]
Grout, B.W.W., K. Shelthon y H.W. Pritchard. 1983. Orthodox behavior of oil palm seed and cryopreservation or excised embryo for genetic conservation. Ann. Bot. 53, 381-384. [ Links ]
International Seed Testing Association (Ista). 1992. Regras para análise de sementes. Coodenacao de laboratório vegetal-clav. Brasilia. [ Links ]
King, M.W. y E.H. Roberts. 1979. The storage of recalcitrant seeds: achievements and possible approaches. International Board for Plant Genetic Resources, Roma. [ Links ]
Murashige, T. y F. Skoog, 1962. A revised medium for rapid grown and bioassays whit tobacco tissue cultures. Physiol. Plant 15, 473-497. [ Links ]
Pammenter, N.W. y P. Berjak. 1999. A review of recalcitrant seed physiology in relation to desiccation tolerance mechanisms. Seed Sci. Res. 9, 13-37. [ Links ]
Roberts, E.H. 1973. Predicting the storage life of seeds. Seed Sci. Technol. 1, 499-514. [ Links ]
Vertucci, C.W. y J.M. Farrant. 1995. Acquisition and loss of desiccation tolerance. pp. 237-271. En: Kigel, J. y G. Galili (eds.). Seed development and germination. Marcel Dekker, Nueva York. [ Links ]

