










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.25 n.2 Bogotá jul./dez. 2007
Fisiología de cultivos
Adriana Tofiño1, Hernán Mauricio Romero2 y Hernán Ceballos3
1 Investigadora Corpoica, Estación Experimental Motilonia, Codazzi, Cesar. atofino@corpoica.org.co
2 Profesor, Departamento de Biología, Universidad Nacional de Colombia, Bogotá. hmromeroa@unal.edu.co
3 Profesor, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Palmira. h.ceballos@cgiar.org
Fecha de recepción: marzo 5 de 2007. Aceptado para publicación: octubre 1 de 2007
RESUMEN
En las plantas superiores la biosíntesis del almidón es sensible al efecto de condiciones ambientales adversas, las cuales actúan sobre el suministro de carbohidratos desde los órganos fuente, la actividad de los vertederos y de las enzimas biosintéticas asociadas. Mediciones de la planta entera in vivo han demostrado que la tasa de flujo de carbono depende además de las propiedades del vertedero y de las características del sistema de transporte. En este documento también se revisan los aspectos del ajuste de esta interacción por el complejo de respuestas celulares inducidas por los estreses abióticos. Además de la problemática de la ruta de señalización del estrés, se elucida su influencia sobre la expresión genética del metabolismo del almidón. Por otro lado, se considera la manera como la planta percibe y responde a las señales ambientales a través de modificaciones en la partición del carbono, mediante el incremento de los niveles de los azúcares solubles y otras estrategias fisiológicas de aclimatación al estrés abiótico.
Palabras clave: expresión genética, señalización del estrés, transporte floemático, biosíntesis de almidón
ABSTRACT
In higher plants, starch biosynthesis is highly sensitive to stress acting on the supply of carbohydrates from source organs, sink activity, and the activities of associated enzymes. Whole plant measurements in vivo have demonstrated that carbon flow rates are additionally dependent on the properties of sink, as well as those of whole transport system. In this article, some issues of the adjustment of this interaction through the complex of cellular responses to abiotic stress are reviewed. The aspects of pathway of stress signaling and its influence on the genetic program of starch expression are also elucidated. Additionally, the pathway, by which a plant senses and responds to environmental cues and endogenous signals through changes in carbon partitioning and increases in soluble sugar cell concentration, and other adaptive physiological strategies are considered.
Key words: gene expression, stress signaling, phloem transport, starch biosynthesis.
Introducción
En las células vegetales las rutas de transducción de señales afectan la trascripción de genes. Las modificaciones en la transcripción pueden llevarse a cabo mediante activación e inhibición de las secuencias de reconocimiento en los promotores por la actividad de factores de trascripción, modificaciones en la cromatina, “empalme” del ARNm y cambios en la estabilidad del ARNm. El proceso de traducción también está influenciado por la dinámica de transducción de señales a través de modificaciones postranscripcionales, tráfico intracelular, degradación de enzimas, alteraciones de la actividad enzimática o la señalización hormonal. La interacción sinérgica entre las rutas de síntesis de ABA y el estrés osmótico, mediante los factores de transcripción DREB y ABREB, ejemplifican este tipo de regulación. Por otro lado, análisis de mutantes individuales de 14 genes aldh de Arabidopsis revelan interacción compleja entre rutas de señalización de respuesta a diversas limitaciones ambientales como sequía, salinidad y frío (Chinnusamy et al., 2005; Buchanan, 1997).
La ruta de biosíntesis del almidón involucra gran número de isoenzimas que interactúan en complejos multiproteínicos y que en casos como el de las enzimas relacionadas con la degradación del almidón, también participan en la“poda” de glucanos altamente ramificados derivados de la síntesis de amilopectina, en el proceso de formación del gránulo (Muñoz et al., 2006; Lloyd et al., 2005). Además del usual proceso refinado de regulación espacio temporal de las enzimas biosintéticas del almidón, el estrés abiótico también afecta la cinética del metabolismo de manera que el balance de los factores ambientales influencia la relación fuente-vertedero y, por tanto, la acumulación final de almidón en los órganos de almacenamiento (Muñoz et al., 2006).
Tradicionalmente, durante el proceso de domesticación de la mayoría de los cultivos no se consideró como criterio de selección la capacidad de los genotipos para soportar las exigencias causadas por las condiciones abióticas adversas, por lo cual, la capacidad de ajuste o tolerancia de las plantas domesticadas a estas condiciones es limitada (Grover et al., 2001). Debido a lo anterior, se han utilizado herramientas genómicas en los programas de mejoramiento en busca de disminuir el efecto de las constricciones ambientales. En arroz, maíz, trigo y cebada se han identificado loci asociados con rasgos cuantitativos (QTL) de características relacionadas con resistencia a estreses abióticos como senescencia prematura, ajuste osmótico, morfología radical, caracteres estomáticos, enrollamiento de las hojas y acumulación de ABA. Así es evidente que las respuestas al estrés tienen un tipo de herencia poligénica que limita las posibilidades de la inserción de genes individuales sobre el metabolismo general de la planta (Humphreys y Humphreys, 2005; Rehman et al., 2005).
La presente revisión se enfoca en las bases moleculares de la modificación de la biosíntesis del almidón como respuesta al estrés en las plantas superiores. Adicionalmente, se consideran los efectos generales de diversos factores ambientales adversos sobre la red de transducción de señales que modifica la relación fuente-vertedero. Por último se ofrece un panorama de las principales estrategias fisiológicas de tolerancia al estrés abiótico en plantas superiores.
Principales factores que afectan la relación fuente-vertedero
Diversos factores ambientales causan grandes diferencias en la producción de biomasa por unidad de área entre especies vegetales y genotipos. El-Sharkawy et al. (1989) encontraron en yuca (Manihot esculenta Crantz) asociación altamente significativa entre tasa fotosintética y productividad. Sin embargo, en la mayoría de los cultivos, los altos rendimientos no suelen estar asociados con alta capacidad fotosintética, debido principalmente a la influencia genética y ambiental multifactorial que determina el rendimiento final. En este sentido los fotoasimilados producidos en la fuente se transportan hacia los posibles vertederos en forma de sacarosa y otros compuestos de la serie rafinosa dependiendo de la especie y el tipo de carga y descarga en el floema vía simplasto/apoplasto. Por tanto, el metabolismo del almidón en el vertedero (semillas, raíces, tubérculos, hojas en desarrollo) está imbricado con la relación entre la síntesis de almidón/sacarosa en la fuente, eficiencia en la exportación de sacarosa, prioridad y potencia del vertedero a través de una compleja y dinámica red autorregulada (Chen y Kim, 2006).
Azúcares
En la actualidad, los mecanismos que determinan la partición de la biomasa no se han develado totalmente. Sin embargo, recientemente se ha propuesto que la concentración de sacarosa desempeña un papel central en la distribución del CO2 asimilado, en buena medida a través de la regulación de la expresión de genes asociados con la síntesis de almidón o fructano. Por ejemplo, se ha evidenciado que la sacarosa es la principal señal asociada con la inducción de la trascripción de la enzima de ramificación del almidón (sbe). Adicionalmente, se han identificado otros azúcares efectores de la sbe como glucosa, glucosa-1-P y furanosa. Por otro lado, la sacarosa también inhibe la síntesis de amilasa. Mientras que el conjunto de azúcares citosólicos se considera mensajero químico intermediario entre el azúcar suministrado por el floema y el azúcar consumido por las “células receptoras” en el vertedero (Muñoz et al., 2006; Rung et al., 2004). Las diferentes funciones de los azúcares en la ruta de transducción de señales de los limitantes abióticos han sido aprovechadas para inducir cambios en la tolerancia de las plantas mediante la transformación con genes foráneos y expresión alterada de la ruta de la sacarosa, debido principalmente a modificaciones en el balance de la partición del carbono y la fotosíntesis. Adicionalmente, el trabajo con mutantes “Knockout” y la utilización de genómica comparativa paulatinamente develarán los interrogantes remanentes sobre la percepción celular de los azúcares y su papel en la dinámica de la partición de la biomasa (Chinnusamy et al., 2005).
Luz y ritmo circadiano
La luz es un factor determinante en la activación y regulación del metabolismo del almidón. Por tanto, modificaciones drásticas en el ritmo circadiano, intensidad y calidad de la luz, afectan la tasa de acumulación y degradación del almidón, removilización desde los vertederos de los productos hidrolíticos del almidón y la concentración de carbohidratos solubles. Los ritmos circadianos mediados por luz funcionan por medio de señales relacionadas con los sistemas de fitocromo que se transportan al núcleo, luego de la recepción de la luz, y modifican la expresión de genes con secuencias sensibles a diversos componentes de la ruta de transducción del fitocromo en sus promotores. La oscilación diurna de la enzima clave en el control de la producción final de almidón (AGPasa), isoformas cloroplásticas de enzimas asociadas con la biosíntesis del almidón como la fructosa- 1,6 bifosfatasa y sacarosa fosfato sintasa (SPS) y el metabolismo de la maltosa, están bajo la influencia del reloj circadiano (Lu et al., 2005; Geigenberger et al., 2004). A medida que se avanza en el entendimiento de la biosíntesis del almidón in vivo, se relaciona el estímulo luminoso con la regulación de nuevas enzimas y modificaciones en la cinética enzimática por la inducción de cambios conformacionales de diferentes complejos multiproteínicos.
Transporte de fotoasimilados
La sacarosa es el azúcar predominante en la corriente del floema. Un pequeño número de familias traslocan adicionalmente oligosacáridos de la serie rafinosa o azúcares alcohólicos como manitol o sorbitol. Evidencias experimentales indican que las fitohormonas están directamente relacionadas con la transferencia de solutos a cortas distancias. Por ejemplo, se ha sugerido asociación entre concentración de ABA y tasa de crecimiento de los frutos en desarrollo. El ABA también estimula la traslocación de azúcares en plantas intactas de fríjol, traslocación de sacarosa dentro del tejido radical de remolacha azucarera y descarga de sacarosa para la absorción del embrión, a partir del apoplasto de las cubiertas de semillas de soya (Rolland et al., 2006; Holtta et al., 2006; Jung Dae et al., 2006). El modelo que mejor explica la traslocación en el floema fue propuesto originalmente por E. Munch en 1930. El mecanismo de flujo por presión o flujo en masa se basa en la transferencia masiva de los solutos desde la fuente hasta el vertedero, a lo largo de un gradiente de presión hidrostática ligada con el flujo del agua en la corriente transpiratoria y recirculación continua del agua en la planta. En un modelo de transporte propuesto recientemente por Holtta et al. (2006), se complementa el concepto de flujo en masa y se pretende explicar el transporte de fotoasimilados entre vertederos competitivos de acuerdo con la dinámica del comportamiento del sistema floemático intacto. El modelo se basa en la extrapolación del modelo sencillo de una fuente y dos vertederos con rutas separadas, valores de resistencia individual y descripción de la función michaeliana específica saturable, a múltiples vertederos competitivos (Holtta et al., 2006).
El proceso de carga en el floema no es exclusivamente pasivo a través de los plasmodemos pues involucra gran cantidad de proteínas transportadoras de iones, agua y azúcares en la membrana. En la carga del floema participan pasos apoplásticos, simplásticos o su combinación. Por ejemplo, en las células vegetales y en las del floema en particular, existe un mecanismo de absorción de sacarosa que depende del ATP ligado a la toma de protones (cotransportadores azúcar-protón) que pueden hacer parte de familias de genes con expresión similar en la fuente/ vertedero o especificidad tisular y temporal de expresión. Se han identificado y clonado genes que codifican formas heterólogas del transportador sacarosa-H+ (SUT1, SUC2), en muchas especies. Estos genes pueden expresarse en la membrana plasmática de las células acompañantes (SUC2) o en la de los vasos cribosos (SUT1). Recientemente, en caña de azúcar se aisló el transportador ShSUT1, con alta homología con la familia de transportadores de sacarosa SUT/SUC. El gen de este transportador, se expresa en hojas maduras que exportan sacarosa y en los entrenudos de los tallos que la acumulan. ShSUT1 podría actuar en la partición de sacarosa entre el tejido vascular y los puntos de almacenamiento en las células parenquimatosas de los entrenudos de los tallos de caña de azúcar. También existen indicios de transportadores específicos del vertedero como RcSCR1 en Ricinus communis, que se ajusta dinámicamente al tamaño del vertedero pero no actúa en la descarga del floema (Eisenbarth y Weig, 2005; Rae et al., 2005).
Adicionalmente se ha identificado mediante herramientas de biología molecular la familia SUT en arroz, compuesta por los transportadores OsSUT1, OsSUT2, OsSUT3, Os- SUT4 y OsSUT5 que presentan expresión diferencial pero traslapada en el transporte de sacarosa a través de membranas en diferentes estados del desarrollo de la planta y tejidos fuente-vertedero (Jung Dae et al., 2006). De manera similar a la carga en el floema, la descarga de los azúcares en el vertedero puede realizarse tanto por la ruta del simplasto como la del apoplasto. La continuidad del flujo pasivo hacia el vertedero se mantiene gracias al gradiente de concentración de sacarosa mediante la hidrólisis de la sacarosa transportada a glucosa y fructosa en las células cercanas a los haces vasculares en el vertedero. Una vez en el apoplasto, la sacarosa se hidroliza mediante la invertasa ácida, la cual está estrechamente ligada a la pared celular (Lloyd et al., 2005; Chen y Kim, 2006).
Factores ambientales como temperatura y fitohormonas influyen el transporte hacia el vertedero pues actúan sobre el crecimiento y la diferenciación histológica del sumidero.La estructura de los componentes del sistema fuente-vertedero también determina el transporte de asimilados, así, en el mutante MIRI66a el nivel de transcripción de ATHB15 (factor de transcripción de expresión exclusiva en tejidos vasculares) disminuye y ocasiona diferenciación celular vascular acelerada a partir de células cambiales/procambiales (Aloni et al., 2006). Las características del sistema fuente-vertedero son dinámicas y varían de acuerdo con la etapa fenológica del cultivo, genotipo y condiciones agroclimáticas. Uno de los objetivos del mejoramiento genético convencional ha sido favorecer el balance del carbono hacia el vertedero. Sin embargo, en cultivos básicos como arroz, se ha llegado al parecer a un tope genético que requiere la implementación de estrategias distintas para incrementar el rendimiento y la eficiencia metabólica del carbono en la fuente y el vertedero. Debido a lo anterior es indispensable el avance en el entendimiento de la fisiología de la interacción fuentevertedero in vivo mediante análisis de flujo metabólico y otros referidos arriba, para establecer los puntos putativos para el mejoramiento dirigido al incremento de la eficiencia de las rutas metabólicas.
Asignación de fotoasimilados en la fuente y el vertedero
Determinadas porciones de carbono se asignan para proveer las necesidades del metabolismo inmediato de la hoja y para satisfacer la demanda metabólica durante el ciclo de 24 horas. Las reservas foliares son en consecuencia accesibles para ser reasignadas y exportadas en las noches o durante los periodos de estrés cuando la fotosíntesis es muy baja. Cerca de la mitad del carbono recién asimilado se exporta inmediatamente de la hoja por la vía del floema; la otra mitad es almacenada transitoriamente como almidón en el cloroplasto (Warren, 2006). En la acumulación de almidón foliar influye el suministro nutricional. La deficiencia de nitrógeno o fósforo en tabaco induce cambios en la actividad de enzimas que afectan la acumulación de carbohidratos como el caso del incremento de la oxidasa alternativa (AOX). En condiciones de estrés, la producción de AOX disminuye el contenido de carbohidratos (Sieger et al., 2005; Knox y Clarke, 2005).
En la tasa de exportación de sacarosa influye el balance entre síntesis de sacarosa y almidón durante el día y la eficiencia degradativa nocturna del almidón. Existe un efecto de competencia entre ambas rutas de síntesis que se regula mediante la actividad de un transportador de triosas en la membrana cloroplástica y un complejo mecanismo de regulación alostérica que responde a la concentración de metabolitos intermediarios (Lloyd et al., 2005; Tofiño et al., 2006). Dado que la regulación metabólica de la biosíntesis de almidón y sacarosa tiene que ver con dos enzimas: la fructosa- 1,6 bifosfatasa y la sacarosa fosfato sintasa (SPS), es razonable esperar que los factores que influyen sobre la asignación del carbono, afecten las actividades de estas enzimas (Weise et al., 2006). La síntesis de sacarosa a partir de glucosa y fructosa fosforiladas se realiza principalmente gracias a la acción enzimática concertada de SPS, sacarosa fosfato fosfatasa y sacarosa sintetasa. Sin embargo, la sobreexpresión de otras enzimas de la ruta de la sacarosa, fosfoglucomutasa (PGM) y pirofosfatasa, tiene efectos pleiotrópicos sobre la fotosíntesis, crecimiento y metabolismo heterotrófico de Arabidopsis, papa y otros cultivos (Rung et al., 2004; Lee et al., 2005).
El reparto del carbono puede alterarse por la acción individual o conjunta del suministro inadecuado de nutrientes, cambios en la actividad de enzimas claves en el control metabólico y por cambios en la fenología de la fuente (Wissuwa et al., 2005). Por ejemplo en el híbrido siempre verde, la velocidad de transporte de asimilados hacia el vertedero y la declinación de la tasa fotosintética fueron más lentas y el periodo de llenado de grano más largo con respecto a cultivares normales de trigo (Gong et al., 2005). Estos aspectos han sido aprovechados por la ingeniería genética en diferentes cultivos como yuca, para mantener la eficiencia de la fuente al aminorar la abscisión de hojas en la planta después de periodos prolongados de sequía, mediante la introducción de un sistema autorregulatorio de inhibición de la senescencia de las hojas asociado con la expresión del gen de isopentenil transferasa, participante en la ruta de síntesis de citoquininas (Zhang y Gruissem, 2005). También se ha trabajado en la modificación enzimática de ADP glucosa pirofosforilasa (AGPasa) para incrementar el rendimiento de almidón y la selección de genotipos con mayor capacidad de absorción de nutrientes bajo estrés hídrico para evitar la caída drástica del rendimiento bajo estrés (Akihiro et al., 2005; Mejía de Tafur, 2002).
Estreses abióticos: genética y mecanismos de respuesta
Las plantas como organismos sésiles necesitan monitoreo constante de los cambios ambientales para modificar y ajustar el desarrollo y metabolismo en concordancia con tales cambios. La respuesta a estos estímulos ambientales requiere un mecanismo integrado donde las señales internas y externas se detectan y ocasionan una reacción apropiada en la planta (Stepanova y Alonso, 2005). Los principales mecanismos fisiológicos de respuesta general al estrés están relacionados con respuestas plásticas como modificaciones en la época de senescencia, cambios en el funcionamiento de las células guarda o en el cierre estomático, mantenimiento de la estabilidad de las membranas celulares y de los organelos y cambios en la elasticidad de la pared celular (Chinnusamy et al., 2005; Stepanova y Alonso, 2005). Se ha observado que la expresión de algunos genes está asociada con la respuesta común a varios tipos de estrés. En arroz, por ejemplo, se clonó un gen homólogo a la glicogenina glucosyltransferasa (OsGGT) que puede relacionarse con el estrés a la inmersión y con la respuesta plástica general a varios estreses ambientales (Chinnusamy et al., 2005).
Además de proteínas asociadas con la respuesta a un amplio espectro de factores abióticos, se han identificado proteínas específicas a cada tipo de estrés, como las proteínas anaeróbicas (ANP) que incluyen enzimas de la glicólisis y la fermentación; proteínas del estrés hídrico (WSP), asociadas con las proteínas de los canales de agua y las que forman osmoprotectantes; las proteínas de embriogénesis tardía (LEA), asociadas con la rotación y la translocación de las proteínas como respuesta al estrés osmótico (Busch et al.,2005; Mariano et al., 2005; Grover et al., 2001; Seguí et al., 2005; Buchanan, 1997).
La percepción del estrés involucra componentes específicos de los cuales no se sabe mucho, en el proceso de señalización siguiente a la percepción, se amplifica y transduce el estímulo mediante la maquinaria de transducción que está compuesta por proteínas quinasas, fosfatasas y proteínas de ligamiento al Ca+2. También se ha registrado la interacción entre diferentes fitorreguladores intermediarios y otras moléculas de la ruta de señalización, relacionados con la senescencia como etileno, glucosa y ABA, y entre auxinas y ABA en la cascada de proteínas de genes mitóticos activados por quinasas (MAPK). De manera similar, la detención del crecimiento bajo estrés y la reanudación bajo condicionesóptimas se ha asociado con el efecto de la fluctuación del balance hormonal entre ABA, giberelinas y auxinas sobre la estabilidad de las proteínas DELLA (figura 1) (Bennet et al., 2005).
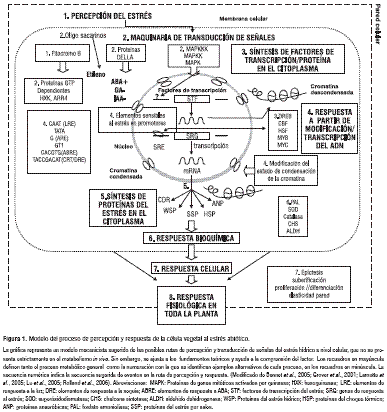
Una vez amplificada en el citoplasma, la señal del estrés se transduce al núcleo, donde se activan genes que codifican factores de transcripción (STF) DREB, MYC, MYB, CBF y HSF, que contienen secuencias complementarias a elementos en los promotores de los genes sensibles al estrés (SRG). La síntesis de factores de transcripción puede involucrar ribosomas citoplásmicos que median la interferencia núcleo-citoplasma; después de la síntesis los factores transactivos reingresan al núcleo donde producen la activación transcripcional de los promotores de genes que contienen elementos ABRE, LTRE, DRE, HSE y ARE, sensibles al estrés (SRE), a los que se ligan factores de transcripción. Los genes de respuesta al estrés (SRG) se traducen en ribosomas citoplásmicos en proteínas del estrés, CDR, WSP, SSP, HSP, ANP, entre otras, responsables de la respuesta bioquímica celular y la consecuente respuesta fisiológica en la planta entera (figura 1) (Lamotte et al., 2005; Chernyad, 2005; Akihiro et al., 2005; Buchanan, 1997).
Respuestas hormonales del metabolismo del almidón al estrés abiótico
Además del papel sobresaliente de las fitohormonas en la regulación de la cascada bioquímica que modifica la expresión genética, ciertos iones como el Ca+2 y algunos metabolitos intermediarios median algunos de los procesos integrados y las reacciones de respuesta de la planta al estrés (Rolland et al., 2006). Se ha demostrado en Arabidopsis y yuca que ABA actúa sinergísticamente con los azúcares en la intensificación de la expresión de los genes de la ruta biosintética del almidón Apl3 y sbe2.2 (Akihiro et al., 2005). El ABA regula también la expresión de otros genes, por ejemplo, aldehído dehidrogenasa (ALDH), OsAPS2 y OsAPL4. El análisis de mutantes de los genes aldh sugiere que las respuestas al estrés se controlan diferencialmente, además de ABA, por las rutas que afectan el metabolismo del azúcar y la composición de los ácidos grasos de las membranas (Kirch et al., 2005). Por otro lado, en arroz la expresión de OsAPS2 y OsAPL4 promovieron la acumulación de almidón de acuerdo con el incremento de ABA y sacarosa (Akihiro et al., 2005). Otras fitohormonas que modifican el metabolismo del almidón bajo estrés son las giberelinas y las citoquininas. Las giberelinas actúan en el desarrollo de las semillas de Arabidopsis mediante la inducción de los genes de α-amilasa.
Algunos investigadores sugieren que las citoquininas también desempeñan papel crucial en la partición de la biomasa pues se ha encontrado relación entre la concentración de varias citoquininas en parte aérea/raíz y acumulación preferencial de azúcar. Este fenómeno afecta la “potencia relativa del vertedero” pues se exporta hacia la raíz cantidades relativamente mayores de carbohidratos (Aloni et al., 2006), además del avance en el entendimiento de los componentes individuales de las bases moleculares de las fitohormonas como las mencionadas en este segmento. En la actualidad es de gran relevancia, el conocimiento de la genética de la integración del balance hormonal y la tasa de proliferación/diferenciación celular, componente fundamental del cambio metabólico en la fuente y el vertedero. Efecto del estrés en el metabolismo del almidón en la fuente La eficiencia del proceso fotosintético depende de factores ambientales como humedad, temperatura e intensidad lumínica. La planta ajusta su eficiencia fotosintética a la máxima intensidad de luz, temperatura, estado de desarrollo, factores genéticos y a la apertura y cierre estomáticos dependientes de la dinámica del continuo sueloplanta-atmósfera. En este sentido, la tasa fotosintética de yuca disminuye notablemente durante periodos de estrés hídrico y la magnitud del efecto depende del genotipo, estado nutricional, intensidad lumínica, y concentración subestomática del CO2 (El-Sharkawy et al., 1989; Mejía de Tafur, 2002).
La fotosíntesis es particularmente sensible al estrés por temperatura. El calor moderado puede ocasionar daños en el fotosistema II, degradación de los tilacoides, desactivación de rubisco y reducción en la tasa fotosintética pues se estimula la producción de formas reactivas de oxígeno, en gran parte por el incremento de la actividad oxigenasa de rubisco. La tolerancia al estrés por temperatura elevada está relacionada, entre otros aspectos, con la capacidad de las plantas de modificar la composición de los lípidos de los tilacoides o por el suministro de isopreno. También es de relevancia el papel de proteínas de choque térmico (HSP), mecanismos de disipación de calor, capacidad transpiratoria, etc. (Sharkey, 2005). Además de daños al proceso fotosintético por la sobreproducción de radicales libres de oxígeno (ROS) (Rey et al., 2005), el estrés ocasiona alteraciones en la biosíntesis de carbohidratos no estructurales. En tejidos foliares de mango y manzana se forman sorbitol y sacarosa a expensas del almidón; mientras que en respuesta al estrés hídrico la actividad de AGPasa decrece y aumenta la de amilasa. Por otro lado, modificaciones en la actividad de las enzimas relacionadas con la síntesis de sacarosa, afectan el contenido y calidad de almidón formado en el vertedero (Rolland et al., 2006; Cheng et al., 2005).
El incremento en la acumulación de azúcares solubles también promueve la tolerancia al estrés por frío en plantas superiores. De tal modo que la degradación del almidón desempeña un papel fundamental en la acumulación de azúcares inducidos por baja temperatura y está asociada con la función de α-glucano/H2O diquinasa, que regula putativamente la degradación del almidón en los plastidios. Otra molécula involucrada en la tolerancia al frío y otros estreses es la trehalosa que actúa como protector de la estabilidad de las estructuras biológicas. Las plantas transgénicas de tabaco y papa con sobreexpresión de trehalosa desarrollan tolerancia a la sequía y a la salinidad (Zhang et al., 2005; Chinussamy et al., 2005). De acuerdo con lo anterior, el mejoramiento genético para la estabilidad de la eficiencia genética de la fuente bajo estrés abiótico se fundamenta en la selección de genotipos con mecanismos eficientes de disipación del calor, eficiencia en la fermentación etanólica, velocidad de síntesis de solutos compatibles y moléculas detoxificantes del H2O2 y radicales libres, pues en esencia el efecto general del estrés abiótico en el metabolismo del almidón en la fuente se basa en la disminución de la eficiencia enzimática y la disrupción de los complejos proteínicos asociados con la fotosíntesis y la ruta biosintética del almidón. La selección asistida por marcadores podría indicar en cada especie los genes candidato putativos de mayor relevancia en la tolerancia al estrés abiótico.
Efecto del estrés abiótico en el metabolismo del almidón en el vertedero
El metabolismo del almidón en el vertedero se ajusta a las fluctuaciones del suplemento de fotosintatos a partir de la fuente y a la dinámica de la interacción de los factores ambientales, pues los cambios en la concentración de azúcares y removilización de reservas constituyen simultáneamente mecanismo de aclimatación y componente de la ruta de señalización ante diferentes tipos de estrés. La fisiología de los carbohidratos en las flores por ejemplo, se afecta fuertemente por el estrés abiótico, que ocasiona infertilidad parcial o total (Lebon et al., 2005). El llenado de los granos del arroz también depende del suplemento de carbohidratos en la fuente conjuntamente con la actividad del vertedero, lo cual incluye la actividad de transportadores de sacarosa, enzimas relacionadas con la síntesis del almidón y diversos factores de regulación transcripcional. Un fenómeno asociado con la cinéticaórgano-específica de las enzimas ligadas al metabolismo de la sacarosa es el desarrollo no uniforme de los cariópsides, de acuerdo con la ubicación en las panículas del arroz, que se explica por diferencias entre cariópsides inferiores y superiores en la relación hexosa/sacarosa, actividad de invertasa ácida y expresión de los genes del metabolismo de la sacarosa (Ishimaru et al., 2005). De modo similar, en fríjol mungo se registraron variaciones cualitativas y cuantitativas del almidón durante el desarrollo de las vainas; también se observó acción sinérgica de SuSy, UGPasa y AGPasa en el control de la partición de sacarosa en ADP glucosa y, por consiguiente, en la regulación de la potencia del vertedero (Muñoz et al., 2006). La temperatura elevada constituye otro factor que ocasiona alteraciones en la acumulación de almidón y amilosa, estructura fina de la amilopectina y actividad de algunas enzimas relacionadas con la síntesis del almidón. Los cambios en la relación amilosa/amilopectina debidos al calor en gran parte se explican por la variación en la actividad de GBSS (Cheng et al., 2005). De manera similar, la temperatura afecta el llenado del grano de trigo y ocasiona la removilización de reservas de carbohidratos no estructura les y de nitrógeno desde el tallo, lo cual sugiere asociación entre senescencia foliar y la eficiencia de la removilización (Tahir y Nakata, 2005).
La biosíntesis del almidón también es sensible al efecto nutricional pues los diferentes elementos actúan como efectores, sustratos, inactivadores o componentes estructurales de las enzimas asociadas con la ruta. En las plantas de garbanzo sometidas al frío, por ejemplo, la concentración de almidón, azúcares reductores y no reductores varió en las semillas de acuerdo con las concentraciones de molibdeno (Nautiyal et al., 2005; Nayyar et al., 2005). De manera similar, las concentraciones inadecuadas de boro afectan la calidad de las semillas e incrementan la actividad de enzimas de la senescencia (Chatterjee et al, 2005). Mientras que diferentes proporciones de hierro, cobalto y fósforo afectan la calidad del fruto en tomate, al disminuir la concentración de clorofila a y b, licopeno, azúcares reductores y almidón (Chatterjee y Chatterjee, 2005). En cebada diferentes niveles de fósforo bajo ambientes saturados con CO2 afectaron la calidad del grano pues incrementaron adenilatos, glutamina y carbohidratos no estructurales a expensas del almidón. También, se registran cambios en la morfología y la expresión genética en las raíces de lupino blanco sometidas a estrés por déficit/exceso de fósforo pues la fotosíntesis y los azúcares están asociados con la transducción de señales que modifican la expresión genética bajo deficiencia de fósforo (Junqi Liu et al., 2005).
Las lesiones resultantes del estrés hídrico se relacionan con efectos dañinos de la desecación sobre el protoplasma. La proporción de almidón total en el grano de cebada y trigo tendió a aumentar en condiciones de estrés hídrico, lo cual puede indicar plasticidad para contrarrestar la desecación del protoplasma (Pessarakli et al., 2005). Otro mecanismo para contrarrestar la desecación del protoplasma de las células del vertedero se identificó en variedades comerciales de yuca bajo estrés por deficiencia hídrica prolongada. La producción de materia seca del cultivo dependió más de la fertilización que del estrés hídrico y el contenido total de nutrientes fue mayor en las plantas bajo estrés hídrico (Mejía de Tafur, 2002). Adicionalmente se evidenció que la acumulación de azúcares solubles y otro tipo de moléculas en las raíces, como norefinetrina en papa y prolina en garbanzo constituye una respuesta fisiológica de aclimatación de las plantas al estrés osmótico (Kukreja et al., 2005).
La concentración interna del oxígeno en el vertedero afecta la partición de asimilados en las plantas y la potencia de vertedero expresada en la raíz (Herschbach et al., 2005). Por ejemplo, en el grano de maíz se altera la síntesis de almidón y la distribución del ATP debido a la fluctuación de la concentración de oxígeno. En condiciones anóxicas la fermentación etanólica es la vía de suministro energético pues debido a las altas tasas de glicólisis y fermentación el agotamiento rápido de los azúcares almacenados incrementa las probabilidades de supervivencia (Mancuso y Marras, 2006). Por ejemplo en Medicago truncatula la actividad de AlaAT incrementó la concentración de alanina que limita la acumulación del compuesto tóxico acetaldehído, producto de la fermentación (Ricoult et al., 2005). Por otro lado, la modificación de la actividad de algunas enzimas relacionadas con la ruta de la síntesis de sacarosa como sacarosa fosfato sintasa (SPS) y sacarosa sintetasa (SUSY), se correlacionaron con la tolerancia a las condiciones anóxicas en potomagetón, pues se observó que el nivel de cDNA del gen susy fue proporcional al alargamiento anaerobio del tallo (Harada et al., 2005).
El efecto del estrés abiótico al igual que los mecanismos de tolerancia al estrés abiótico son similares en la fuente y el vertedero. Sin embargo, aunque el efecto del estrés actúa simultáneamente limitando tanto la fotosíntesis como la absorción de nutrientes, el principal problema para el rendimiento de los cultivos es la removilización y deterioro en la calidad de las reservas durante el periodo de estrés y durante la restauración de la normalidad metabólica posterior a la desaparición de la condición limitante. Algunos cultivos presentan tolerancia natural frente a ciertas constricciones ambientales como la tolerancia a sequía de la yuca. Sin embargo el cultivo es extremadamente susceptible a la inundación. Cuando no existe variabilidad genética en las características de tolerancia al estrés dentro del germoplasma de la especie, el mejoramiento genético debe combinarse con estrategias productoras de variabilidad como la ingeniería genética y la mutagénesis inducida. Las estrategias genómicas deben apuntar hacia la estabilidad de las enzimas del almidón en la fuente y el vertedero, promoción de la estabilidad de la fotosíntesis neta y la absorción de nutrientes y la modificación de la cinética michaeliana de las enzimas degradadoras del almidón bajo estrés, para promover la removilización de almidón de órganos distintos al del interés comercial. Por ejemplo, la movilización rápida de reservas del tallo y hojas maduras de la planta de yuca en vez del almidón de las raíces. Estos avances solo serán posibles a través del descubrimiento de promotores organoespecíficos y el desarrollo de minicromosomas para la inserción de genes múltiples (Chynnusamy et al., 2005).
Literatura citada
Akihiro, T., K. Mizuno y T. Fujimura. 2005. Gene expression of ADP-glucose pyrophosphorylase and starch contents in rice cultured cells are cooperatively regulated by sucrose and ABA. Plant Cell Physiol. 46(6), 937-946. [ Links ]
Aloni, R., E. Aloni, M. Langhans y C. Ullrich. 2006. Role of cytokinin and auxin in shaping root architecture: Regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. AOB 97(5), 883-893. [ Links ]
Bennet, M., C. Bellini y D. Van Der Straten. 2005. Integrative biology: dissecting cross-talk between plant signaling pathways. Physiol. Plant. 123, 109. [ Links ]
Buchanan-Wollaston,V. 1997. The molecular biology of leaf senescence. J. Expl. Bot. 48(307), 181-199. [ Links ]
Busch, W., M. Wunderlich y F. Schoffl. 2005. Identification of novel heat shock factor-dependent genes and biochemical pathways in Arabidopsis thaliana. Plant. J. 41, 1-14. [ Links ]
Chatterjee, C., Sinha Pratima y B. Dube. 2005. Biochemical changes, yield, and quality of gram under boron stress. Comm. Soil Sci. Plant Anal. 36, 13-14. [ Links ]
Chatterjee, J. y C. Chatterjee. 2005. Deterioration of fruit quality of tomato by excess cobalt and its amelioration. Comm. Soil Sci. Plant Anal. 36 (13-14), 1931-1945. [ Links ]
Chen, X. y J. Kim. 2006. Transport of macromolecules through plasmodesmata and the phloem. Physiol. Plant. 126(4), 560-567. [ Links ]
Cheng, F., L. Zhong, N. Zhao, Y. Liu y G. Zhang. 2005. Temperature induced changes in the starch components and biosynthetic enzymes of two rice varieties. Plant Growth Regulat. 46(1), 87-95. [ Links ]
Chernyad, I. 2005. Effect of water stress on the photosynthetic apparatus of plants and the protective role of cytokinins: A review. Prikladnaya Biokh. Mikrobiol. 41(2), 133-147. [ Links ]
Chinnusamy, V., Liming Xiong y Jian- Kang Zhu. 2005. Chapter two: Use of Genetic Engineering and Molecular Biology Approaches for Crop Inprovement for Strees Environments. pp. 47- 95. En: Ashraf, M. y P. Harris (eds.). Abiotic stresses plant resistance through breeding and molecular approaches. Food Products Press, Binghamton. [ Links ]
Eisenbarth, D. y A. Weig. 2005. Sucrose carrier RcSCR1 is involved in sucrose retrieval, but not in sucrose unloading in growing hypocotyls of Ricinus communis L. Plant Biol. 7, 98-103. [ Links ]
El-Sharkawy, M., J. Cock, E. Marcio y M. Porto. 1989. Características fotossintéticas da mandioca(Manihot esculenta Crantz). Rev. Bras. Fisiol. Veg. 1(2), 143-154. [ Links ]
Geigenberger, P., A. Kolbe y A. Tiessen. 2004. Redox regulation of carbon storage and partitioning in response to light and sugars. J. Expl. Bot. 55(406), 2131-2145. [ Links ]
Gong, Y., J. Zhang, J. Gao, J. Lu y J. Wang. 2005. Slow Export of Photoassimilate from Stay-green Leaves during Late Grain- Filling Stage in Hybrid Winter Wheat (Triticum aestivum L.). J. Agron. Crop. Sci. 4(191), 292. [ Links ]
Grover, A., A. Kapoor, O. Satya, S. Agarwal, Ch. Sahi, S. Katiyar, M. Agarwal y H. Dubey. 2001. Understanding molecular alphabets of the plant abiotic stress responses. Curr Sci. 80(2), 206- 216. [ Links ]
Harada T., S. Satoh, T. Yoshioka y K. Ishizawa. 2005. Expression of sucrose synthase genes involved in enhanced elongation of pondweed (Potamogeton distinctus) turions under anoxia. Ann. Bot. 96(4), 683-692. [ Links ]
Herschbach, C., S. Mult, J. Kreuzwieser y S. Kopriva. 2005. Influence of anoxia on whole plant sulphur nutrition of flooding-tolerant poplar (Populus tremula × P. alba). Plant Cell Environ. 28(2), 167-175. [ Links ]
Hölttä, T., T. Vesala, S. Sevanto, M. Perämäki y E. Nikinmaa. 2006. Modeling xylem and phloem water flows in trees according to cohesion theory and Münch hypothesis. Trees- struct. Func. 20(1), 67-78. [ Links ]
Humphreys, M. y M.W. Humpreys. 2005. Breeding for Stress Resistance: General principles. pp. 19-14. En: Arshraf M. y P. Harris. (eds.). Abiotic stresses plant resistance through breeding and molecular approaches. Food Products Press, Binghamton. [ Links ]
Ishimaru, T., T. Hirose, T. Matsuda, A. Goto, T. Kazunari, H. Sasaki, O. Tera, R. Ishii, R. Ohsugi y T. Yamagishi. 2005. Expression patterns of genes encoding carbohydrate-metabolizing enzymes and their relationship to grain filling in rice (Oryza sativa L.): Comparison of caryopses located at different positions in a panicle. Plant Cell Physiol. 46(4), 620-628. [ Links ]
Jung Dae Lim, Cho Jung-Il, Park Youn-Il, Hahn Tae-Ryong, Choi Sang-Bong, Jeon Jong-Seong. 2006. Sucrose transport from source to sink seeds in rice. Physiol. Plant. 126(4), 572-581. [ Links ]
Junqi Liu, D. Samac, B. Bucciarelli, D. Allan y C. Vance. 2005. Signaling of phosphorus deficiency-induced gene expression in white lupin requires sugar and phloem transport. Plant J. 41, 257-268. [ Links ]
Kirch, H., S. Schlingensiepen, S. Kotchoni, R. Sunkar y Bartels. 2005. Detailed expression analysis of selected genes of the aldehyde dehydrogenase (ALDH) gene superfamily in Arabidopsis thaliana. Plant. Mol. Biol. 3(57), 315-332. [ Links ]
Knox, K. P. Clarke; 2005. Nutrient availability induces contrasting allocation and starch formation in resprouting and obligate seeding shrubs. Funct. Ecol. 4(19), 690-670. [ Links ]
Kukreja, S., A. Nandwal, N. Kumar, S. Sharma, S. Sharma, V. Unvi y P. Sharma. 2005. Plant water status, H2O2 scavenging enzymes, ethylene evolution and membrane integrity of Cicer arietinum roots as affected by salinity. Biol. Plant. 49(2), 305-308. [ Links ]
Lamotte, O., C. Courtois, L. Barnavon, A. Pugin y D. Wendehenne. 2005. Nitric oxide in plants: the biosynthesis and cell signalling properties of a fascinating molecule. Planta 221, 1-4. [ Links ]
Lebon, G., E. Duchene, Brun,O. y C. Clement. 2005. Phenology of Flowering and Starch Accumulation in Grape (Vitis vinifera L.) Cuttings and Vines. Ann. Bot. 95, 943-948. [ Links ]
Lee J., D.S. Lee, S. Bhoo, J.S. Jeon, Y.H. Lee y T.R. Hahn. 2005. Transgenic Arabidopsis plants expressing Escherichia coli pyrophosphatase display both altered carbon partitioning in their source leaves and reduced photosynthetic activity. Plant Cell Rep. 24(6), 374-382. [ Links ]
Lloyd, J., J. Kossmann y G. Ritte. 2005. Leaf starch degradation comes out of the shadows. Trends Plant Sci. 3(10), 130-137. [ Links ]
Lu, Y., J.P. Gehan y T. Sharkey. 2005. Daylength and circadian effects on starch degradation and maltose metabolism. Plant Physiol. 138(4), 2280-2291. [ Links ]
Mancuso, S. A. Marras. 2006. Adaptative response of Vitis root to anoxia. Plant Cell Physiol. 47(3), 401-409. [ Links ]
Mariano, E., R. Jorge; G. Keltjens y M. Menossi. 2005. Metabolism and root exudation of organic acid anions under aluminium stress. Braz. J. Plant. Physiol. 17(1), 157-172. [ Links ]
Mejía de Tafur, M. 2002. Fisiología de la yuca Manihot esculenta Crantz. Pp. 34-45. En: Ospina, B. y H. Ceballos. (eds.). La yuca en el tercer milenio: sistemas modernos de producción, procesamiento, utilización y comercialización. CIAT, Calí. [ Links ]
Muñoz, F., M. Morán Zorzano, N. Alonso-Casajús, E. Baroja- Fernández, E. Etxeberria y J. Pozueta-Romero. 2006. New enzymes, new pathways an alternative view on starch biosynthesis in both photosynthetic and heterotrophic tissues of plants. Biocatalysis and Biotransformation 24(1-2), 63-76. [ Links ]
Nayyar, H., T. Bains, S. Kumar y G. Kaur. 2005. Chilling effects during seed filling on accumulation of seed reserves and yield of chickpea. J. Sci Food Agric. 85, 1925-1930. [ Links ]
Nautiyal, N., S. Singh y C.H. Chatterjee, 2005. Seed reserves of chickpea in relation to molybdenum supply. J. Sci Food Agric. 85, 860-864. [ Links ]
Pessarakli, M., P. Morgan y J. Gilbert. 2005. Dry-matter yield, protein synthesis, starch, and fiber content of barley and wheat plants under two irrigation regimes. J. Plant Nutr. 7(28), 1227-1241. [ Links ]
Rae, A., J. Perroux C.H. Grof. 2005. Sucrose partitioning between vascular bundles and storage parenchyma in the sugarcane stem: a potential role for the ShSUT1 sucrose transporter. Planta 220(6), 817-825. [ Links ]
Rehman, S., P. Harris y M. Ashraf. 2005. Stress Environments and their impact on crop production. pp. 3-18. En: Ashraf, M. y P. Harris. (eds.). Abiotic stresses plant resistance through breeding and molecular approaches. Food Products Press, Binghamton. [ Links ]
Rey, P., S. Cuiné, F. Eymery, J. Garin; M. Court, J. Jacquot, N. Rouhier y M. Broin. 2005. Analysis of the proteins targeted by CDSP32, a plastidic thioredoxin participating in oxidative stress responses. Plant J. 1(41), 31-42. [ Links ]
Ricoult, C., J. Cliquet y A. Limami. 2005. Stimulation of alanine amino transferase (AlaAT) gene expression and alanine accumulation in embryo axis of the model legume Medicago truncatula contribute to anoxia stress tolerance. Physiol. Plant. 1(123), 30. [ Links ]
Rolland, F., E. Baena-González y J. Sheen. 2006. Sugar sensing and signaling in plants: Conserved and Novel Mechanisms. Annu. Rev. Plant Biol. 57, 675-709. [ Links ]
Rung, J., H. Draborg, K. Jorgensen, y T. Nielsen. 2004. Carbon partitioning in leaves and tubers of transgenic potato plants with reduced activity of fructose-6-phosphate,2-kinase/fructose-2,6-bisphosphatase. Physiol Plant. 121(2), 204-212. [ Links ]
Seguí, J., P. Testillano, S. Jouannic, Y. Henry y M. Risueño. 2005. Mitogen- activated protein kinases are developmentally regulated during stress-induced microspore embryogenesis in Brassica napus L. Histochem. Cell Biol. 4-5(123), 541-551. [ Links ]
Sharkey, T. 2005. Effects of moderate heat stress on photosynthesis: importance of thylakoid reactions, rubisco deactivation, reactive oxygen species, and thermotolerance provided by isoprene. Plant Cell Environ. 3(28), 269. [ Links ]
Sieger, S., B. Kristensen, C. Robson, S. Amirsadeghi, E. Eng, A. Abdel-Mesih, Møller, I. Y G. Vanlerberghe. 2005. The role of alternative oxidase in modulating carbon use efficiency and growth during macronutrient stress in tobacco cells. J. Exp. Bot. 56(416), 1499-1515. [ Links ]
Stepanova, A. y J. Alonso. 2005. Ethylene signalling and response pathway: a unique signaling cascade with a multiple of inputs and outputs. Physiol. Plant. 123, 195-206. [ Links ]
Tahir, I. y N. Nakata. 2005. Remobilization of nitrogen and carbohydrate from stems of bread wheat in response to heat stress during grain filling. J. Agron. Crop. Sci. 2(191), 106. [ Links ]
Tofiño, A., M. Fregene, H. Ceballos y D. Cabal. 2006. Regulación del almidón en plantas terrestres: perspectivas de modificación. Acta Agron. 55(1), 1-13. [ Links ]
Warren, CH. 2006. Why does Photosynthesis decrease with needle age in Pinus pinaster? Trees Structure and Function 20(2),157-164 [ Links ]
Weise, S., S. Schrader, K. Kleinbeck y T. Sharkey. 2006. Carbon Balance and Circadian Regulation of Hydrolytic and Phosphorolytic Breakdown of Transitory Starch. Plant Physiol. 141, 879-886. [ Links ]
Wissuwa, M., G. Gamat y A. Ismail. 2005. Is root growth under phosphorus deficiency affected by source or sink limitations? J. Expl. Bot. 56(417),1943-1950. [ Links ]
Zhang, S., B. Yang, C. Feng y H. Tang. 2005. Genetic transformation of tobacco with the trehalose synthase gene from grifola frondosa Fr. enhances the resistance to drought and salt in tobacco. J. Integ. Plant Biol. 5(47), 579. [ Links ]
Zhang, P. y W. Gruissem. 2005. Leaf senescence-Inducible expression of isopentenyl transferase in cassava rendering it resistant to drought stress. The global food and product chain: Dynamics, innovations, conflicts, strategies, Deutscher Tropentag 2005. October 11-13. University of Hohenheim, Stuttgart. [ Links ]
Zhang, P. y W. Gruissem. 2005. Leaf senescence-Inducible expression of isopentenyl transferase in cassava rendering it resistant to drought stress. 2nd International Conference on Integrated Approaches to Sustain and Improve Plant Production Under Drought Stress. September 24-28. Rome. [ Links ]

