










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.25 n.2 Bogotá jul./dez. 2007
Álvaro Rincón1, Gustavo A. Ligarreto2 y Danny Sanjuanelo3
1 Investigador Programa de Fisiología y Nutrición Animal, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Centro de Investigación La Libertad, Villavicencio, Meta. arincon@corpoica.org.co
2 Profesor asociado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. galigarretom@unal.edu.co
3 Profesor, Universidad de Ciencias Aplicadas y Ambientales (UDCA), Bogotá. dwsanjuaneloc@gmail.com
Fecha de recepción: agosto 9 de 2007. Aceptado para publicación: octubre 1 de 2007
RESUMEN
El experimento se desarrolló en el piedemonte llanero de Colombia, donde el establecimiento y recuperación de praderas con el cultivo del maíz, es una opción productiva para los sistemas ganaderos de la región. En un diseño de bloques completos al azar con cuatro replicaciones, se evaluó el establecimiento de maíz y pastos en monocultivo y asociados (Brachiaria decumbens, Brachiaria cv. Mulato y Toledo (Brachiaria brizantha)), con el fin de determinar la existencia de efectos de competencia sobre los rendimientos de grano de maíz y sobre la producción de forraje de los pastos. El periodo de evaluación comprendió desde los 15 hasta los 90 días después de la siembra (dds), época en la cual el maíz ya se encontraba en proceso de secado. El crecimiento del maíz no fue afectado por la presencia de los pastos, obteniéndose el índice de área foliar de 3, a los 45 dds y la mayor producción de masa seca a los 66 dds con 29 g MS/planta. Por su parte, el crecimiento de los pastos sí fue afectado por la presencia del maíz durante todo el periodo de evaluación. El área foliar y la masa seca en los pastos asociados con maíz fue inferior en un 50% con respecto a los pastos establecidos sin maíz, de forma que a los 75 dds el índice de área foliar de los pastos asociados con maíz fue de 1,0 y en los pastos en monocultivo fue de 2,6. De igual forma, la masa seca fue de 68 g·m-2 y de 140 g·m-2 en el mismo orden. La asociación de maíz con pastos es una buena opción para el establecimiento o renovación de praderas porque los rendimientos de maíz no fueron afectados por la presencia de los pastos. Los pastos asociados con el maíz presentaron un crecimiento más lento en los primeros 75 dds, sin afectar el crecimiento del maíz, sin embargo durante los 15 días siguientes, el índice de área foliar se incrementó en 170% y la masa seca en 53%, por el secado natural de la planta de maíz que permitió la entrada de radiación solar directa.
Palabras clave: asociación, cultivos, índice de área foliar, crecimiento, masa seca.
ABSTRACT
The experiment was carried out in the Piedemont Plains of Colombia, where the establishment and recuperation of grasslands associated with corn culture is a productive option for the livestock farmers of the region. Under a randomized complete block design with four replicates, the establishment of corn in monoculture and association with grasses (Brachiaria decumbens, Brachiaria cv. Mulato and Toledo (Brachiaria brizantha)) was evaluated in order to determine the existence of competition and its effect on the grain yield in corn and forage production in grasses. The period of evaluation was from 15 up to 90 days after sowing (d.a.s.), a time period, in which corn was already in drying process. The growth of corn was not affected by the presence of grasses, being characterized with leaf area index (LAI) equal to 3 at 45 d.a.s. and the highest dry weight production of 29 g DM/plant at 66 d.a.s. At the same time, the presence of corn do affected the growth of grasses during the whole period of evaluation. The leaf area and dry weight of grasses associated with corn were reduced on 50% as compared with those of grasses grown without corn, so that at 75 d.a.s. the LAI was 1.0 and 2.6 in grasses associated with corn or monoculture, respectively. In any case, the dry weight was of 68 g·m-2 and 140 g·m-2 for the same treatments. The cultivation of corn with grasses is a good option for the establishment or renovation of grasslands because the corn yields were not affected by the grass presence. The grasses associated with corn presented a slow growth during the first 75 d.a.s., without affecting the growth of corn, however, during the 15 following days, the LAI and dry weight were increased on 170% and 53%, respectively, due to natural drying of corn plants that allowed the penetration of direct solar radiation.
Key words: association, crops, leaf area index, growth, dry weight.
Introducción
La asociación del cultivo de maíz con los pastos se presenta como una opción de alta viabilidad para el establecimiento y la renovación de praderas en los Llanos Orientales de Colombia. Este sistema mejora la fertilidad del suelo, se obtienen mayores producciones de biomasa de pasto de mejor calidad, y se beneficia el productor porque con la cosecha del cultivo cubre en gran medida los costos de la renovación de la pradera y tiene disponibilidad de mejores pastos. Este sistema presenta una productividad animal mayor en un tiempo más corto (Kluthcouski y Aidar, 2001; Pereira y Pacheco, 2001). El establecimiento de maíz y pastos con sistema fotosintético C4 y alta capacidad de aprovechamiento de la radiación solar del trópico, lleva a la necesidad de conocer aspectos relacionados con el crecimiento de las especies establecidas en forma simultánea en el mismo espacio. Los aspectos más notables de estas especies son las altas tasas de fotosíntesis y su respuesta a niveles crecientes de intensidad de luz, hasta llegar a la plena iluminación del sol. Otra diferencia importante es la carencia de fotorrespiración durante el proceso fotosintético (Montaldi, 1995; Medrano y Flexas, 2001).
La mayor tasa fotosintética en pastos tropicales comparada con las leguminosas, no se debe simplemente a la menor fotorrespiración en las C4, que pueden explicar menos del 50% de la diferencia. Un efecto principal parece estar relacionado con las grandes diferencias en las resistencias intracelulares a la transferencia de CO2 (Black, 1976). Cuando varias especies se siembran en mezcla, su respuesta a los factores ambientales es diferente que cuando las mismas especies son sembradas separadamente (Clavijo, 1992). Un rápido crecimiento y una mayor expansión de hojas y raíces se señalan como características importantes de plantas competidoras. Una planta que crece más rápido que su vecina utilizará una mayor cantidad de un determinado recurso disponible e incrementará su masa seca de manera que se convierte en ventaja para competir (Roush y Radosevich, 1985, citado por Clavijo, 1992). En este sentido, una medida como la tasa de crecimiento relativo (TCR) podría servir como indicador de la habilidad competitiva potencial entre cultivos y malezas. Clavijo (1992), estudiando las interacciones entre arroz rojo y dos variedades de arroz, concluyó que el arroz rojo considerada como una de las principales malezas en el cultivo de arroz, competía con las variedades de arroz produciendo mayor número de macollas y hojas y mayor cantidad de masa seca, que cuando se sembraba en monocultivo. También se han realizado evaluaciones de competencia entre arroz y pasto establecidos en forma simultánea, encontrándose efectos sobre los rendimientos de arroz especialmente cuando la menor fertilidad del suelo o la alta densidad de Brachiaria, favorece el desarrollo del pasto (Silveira et al., 1999; Sans et al., 1999).
Un método de estudio capaz de abrir amplias perspectivas sobre las posibilidades de cuantificar y posteriormente aumentar la producción vegetal es el conocido con el nombre de análisis de crecimiento y del desarrollo. El crecimiento se define como el aumento de la masa de un organismo y es el resultado de la formación de nuevas células y del aumento de peso de la masa seca del mismo. En la práctica, el crecimiento se obtiene midiendo el aumento del peso de la masa seca de la planta entera. El desarrollo, por el contrario, implica diferenciación, es decir, una modificación más profunda que comparta organización y especialización anatómica y fisiológica (Hunt, 1982). La producción de masa seca en la plantas es el resultado neto del balance entre los procesos metabólicos básicos. La masa seca es producida en el proceso fotosintético y los productos se distribuyen y acumulan en diferentes órganos de la planta (Duarte et al., 2005).
Los métodos de análisis de crecimiento han sido particularmente elaborados para estudiar este proceso durante la fase vegetativa. A partir de los datos de área foliar y masa seca, se pueden calcular los índices fisiológicos, dentro de los cuales los más importantes son: índice de área foliar (IAF), cuyo incremento durante las primeras etapas del crecimiento está asociado con la fase de macollamiento y su disminución, al final del ciclo del cultivo, se debe a la senescencia y muerte de las hojas dada en forma secuencial desde la base de la planta hasta la zona apical. La tasa de crecimiento relativo (TCR) representa la capacidad de la planta para producir material nuevo y depende de la fotosíntesis y la respiración (Hunt, 1982). La TCR presenta inicialmente valores altos los cuales disminuyen a medida que transcurre la edad de la planta con un comportamiento similar al del área foliar. La tasa de asimilación neta (TAN) es un índice importante de la capacidad fotosintética del aparato asimilador; esta no mide la fotosíntesis real sino la eficiencia neta en la producción de masa seca por fotosíntesis menos la pérdida de asimilados en los procesos de respiración.
Dada la importancia que está adquiriendo los sistemas agropastoriles para el desarrollo de una ganadería competitiva en la región, se estableció este experimento con el fin de determinar posibles efectos de competencia que pudieran afectar los rendimientos de los componentes de la asociación maíz + pastos y así aportar en el conocimiento sobre el análisis de crecimiento de esta asociación, en la cual son escasos los reportes de literatura en estudios bajo las condiciones ambientales del piedemonte llanero colombiano.
Materiales y métodos
El experimento se desarrolló en el primer semestre de 2005 en un Oxisol de terraza media del Centro de Investigaciones La Libertad, Corpoica, ubicado en el municipio de Villavicencio (Meta, Colombia) a 9º 6` de latitud norte y 73º 34` de longitud oeste, a 330 msnm, la precipitación anual promedia de los últimos 30 años ha sido de 2.900 mm, el promedio de temperatura es de 26 °C y una humedad relativa de 85% en la época lluviosa y 65% en la época seca. Los suelos son muy ácidos con una saturación de aluminio de 71% mientras que la saturación de bases fue 25%. Los nutrientes más deficientes fueron fósforo, calcio y magnesio con 1 ppm, 0,44 y 0,12 meq por 100 g de suelo, respectivamente. Respecto a las características físicas de los suelos se encontró que el contenido promedio de arena, limo y arcilla fue de 59,8, 24,8 y 15,4% respectivamente, valores que permiten clasificarlo como un suelo franco arenoso, la densidad aparente presentó un valor de 1,36 g·cc-1, en tanto la densidad real fue de 2,53 g·cc-1 y la porosidad de 46%.
Tratamientos y diseño experimental
Se establecieron siete tratamientos: maíz solo, B. decumbens en asocio con maíz, B. decumbens solo, pasto híbrido de Brachiaria cv. Mulato en asocio con maíz, pasto Mulato solo, pasto Toledo (Brachiaria brizantha) en asocio con maíz y pasto Toledo solo. Estos tratamientos se distribuyeron en un diseño de bloques completos al azar con cuatro repeticiones. El área de cada unidad experimental fue de 30 m2.
Establecimiento del experimento
La labranza del suelo se realizó mediante un pase de cincel rígido, dos pases de rastra y un pase de pulidor; posteriormente se hizo la siembra en surcos de maíz separados a 0,8 m y en medio se establecieron los surcos de pasto. La densidad de siembra fue de 22 kg·ha-1 de maíz híbrido “Master” y de 4 kg·ha-1 de pasto (Rincón et al., 2002). Antes de la siembra se aplicaron los correctivos necesarios para disminuir la saturación de aluminio al 50% para lo cual fue necesario aplicar 1,5 t·ha-1 de cal dolomítica, además se aplicó 400 kg·ha-1 de yeso agrícola y 300 kg·ha-1 de roca fosfórica, por los bajos contenidos de azufre y fósforo en este suelo. Al momento de la siembra se aplicó en el surco del maíz 150 kg·ha-1 de fosfato diamónico junto con 20 kg de borozinco y 50 kg de cloruro de potasio. Posteriormente se aplicaron 150 kg de nitrógeno y 100 kg de cloruro de potasio, fraccionándolos en partes iguales a los 15 y 35 días después de la siembra. El control de gusano cogollero (Spodoptera sp.) se realizó cuando el maíz tenía una edad de 30 días, con 200 cm3·ha-1 del inhibidor de quitina a base de Clorfluazuron mezclados con 30 kg de melaza y 300 L·ha-1 de agua. La cosecha del maíz se realizó en forma manual a la edad del cultivo de 120 días.
Evaluaciones
Las evaluaciones sobre altura, materia seca y área foliar en el maíz, se realizaron a la edad de 15, 30, 45, 55, 66, 77 y 90 días, mientras que las evaluaciones sobre masa seca y área foliar de los pastos solos y asociados con el maíz, se realizaron a la edad de 37, 60, 75 y 94 días. Para la estimación de masa seca las muestras fueron sometidas a una temperatura de 70 °C en un horno durante 72 h. Para medir el área foliar se utilizó un equipo LI 3100 Licor. De acuerdo con Radford (1967) y Hunt (1990) los índices de crecimiento comúnmente utilizados se presentan en la tabla 1, y son: el IAF, entendido como el área de las hojas del cultivo presente en el área de un metro cuadrado de suelo, por tanto, no tiene unidades; la TCA mide el incremento de la masa seca por unidad de tiempo (g·día-1); la TCR expresa el incremento en masa seca en un intervalo con relación al peso inicial (g·g-1 ·día-1) y la TAN que mide la acumulación de masa seca en función del área foliar por unidad de tiempo. Para su cálculo se utilizó la información obtenida de masa seca y del área foliar. Dicha información fue analizada mediante el paquete estadístico Statistical Analysis System (SAS) versión 9.1. Los resultados se sometieron a análisis de varianza para determinar la significancia, y la comparación de medias se determinó por la prueba de comparación de Tukey.

Resultados y discusión
Índices fisiológicos en la planta de maíz establecida en monocultivo y asociada con pastos
Durante los cuatro meses de desarrollo de la asociación (abril a julio), la precipitación fue de 1.424 mm, la cual correspondió a un 51% de la precipitación total del año 2005; por consiguiente, no se presentó limitantes por agua, considerando que el maíz requiere de al menos 500 mm de agua bien distribuidos (Lafitte, 2002). En todos los tratamientos en los que se estableció el maíz en monocultivo (sin pasto) y maíz asociado con B. decumbens, Mulato y Toledo, no se presentaron diferencias significativas (P > 0,05) en altura, masa seca y área foliar en la planta de maíz, durante el periodo de 90 días de evaluación de crecimiento (tabla 2).
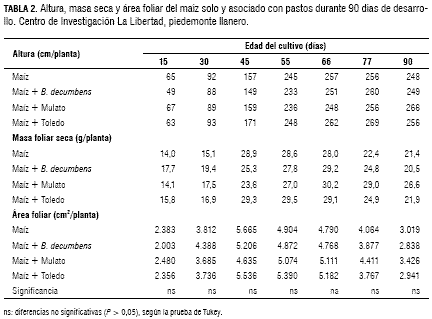
La altura promedio a los 15 días fue de 0,58 m y llegó hasta 2,60 m a la edad de 77 días. El mayor incremento de altura de la planta de maíz se presentó en los primeros 55 días de crecimiento, ya que durante este tiempo la planta alcanzó una altura de 2,40 m, luego en los siguientes 45 días sólo aumento 0,20 m. La masa seca foliar promedia en los primeros 15 días de desarrollo de la planta de maíz fue de 15,4 g/planta y la máxima producción de biomasa foliar seca se produjo a los 66 días después de la siembra (dds) con 29,1 g/planta coincidiendo con el momento de llenado de grano. Posteriormente, la masa seca de las hojas disminuyó por senescencia de estas, de forma que a los 90 dds el promedio alcanzado fue de 22,6 g hasta planta. El área foliar de la planta de maíz a los 15 dds presentó un promedio de 2.256 cm2, la cual aumentó hasta llegar a su máximo valor a la edad de 45 días con 5.261 cm2. Se presentó una relación directa entre la producción de masa seca y el área foliar de la planta de maíz, sin embargo, los valores máximos de estos parámetros, se presentaron en diferentes edades del cultivo, es así como la mayor área foliar coincidió con la máxima floración de la planta de maíz (45 dds) y la máxima producción de masa foliar seca coincidió con el llenado del grano, que fue a los 66 dds (figura 1).
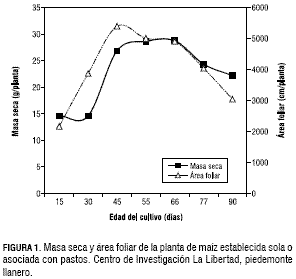
En la figura 2 aparece el IAF de la planta de maíz durante el periodo de evaluación que comprendió desde los 15 dds hasta los 90 dds. Se puede apreciar la misma tendencia en el maíz establecido en monocultivo y asociado con pastos, con excepción del maíz establecido en monocultivo y en el maíz asociado con Toledo, porque a los 45 dds, el IAF fue de 3,3, resultando ser más alto con respecto a las demás asociaciones que presentaron un IAF promedio de 2,9. Sin embargo, después de los 55 dds, este parámetro estuvo alrededor de 3 en todos los tratamientos y después de 66 dds, disminuyó a 2,9, hasta llegar a un IAF de 1,8 a los 90 días, fecha de la última evaluación de crecimiento, en todos los tratamientos. El mayor IAF encontrado a la edad de 45 días coincidió con el inicio del desarrollo reproductivo de la planta, momento en el cual necesita de su máxima capacidad fotosintética para acumular carbohidratos y masa seca, para ser utilizados posteriormente en la formación de la mazorca (Lafitte, 2002).
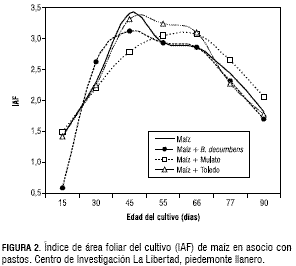
La TCA del maíz solo y asociado con el pasto Toledo, fue similar durante todo el desarrollo del cultivo, tal como sucedió con el IAF, destacándose una mayor TCA a los 30 dds con un promedio de 0,86 g·día-1 de MS. En las asociaciones del maíz con el pasto Mulato y el B. decumbens, la TCA alcanzó el máximo valor de 0,4 g·día-1 a los 30 días, pero a diferencia del TCA del maíz solo y el maíz asociado con Toledo, el incremento de masa seca permaneció entre 0,3 y 0,2 g·día-1 durante 25 días (desde los 45 dds hasta cuando el maíz cumplió una edad de 55 días).En tanto, en el maíz establecido en monocultivo y en asociación con Toledo, el periodo de mayor incremento de masa seca fue a la edad de 30 días para descender a una TCA cercana a cero a los 45 dds (figura 3).
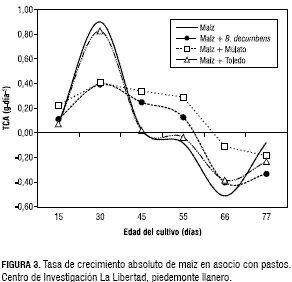
El comportamiento y la tendencia de la TCR fueron similares a la TCA y disminuyeron de manera progresiva (figura 4). Las curvas observadas en la TCR en el maíz asociado con el B. decumbens y el Mulato fue mayor a los 30 dds con un valor de 0,02 g·g-1·día-1 y luego disminuyó levemente hasta los 55 dds con una TCR de 0,01 g·g-1·día-1, posteriormente, la TCR fue negativa. En el maíz solo y asociado con Toledo la TCR alcanzó un valor de 0,04 g·g-1·día-1, a los 30 dds, sin embargo después de los 45 dds la TCR fue negativa. A medida que aumenta el IAF, se aumenta el sombreamiento y disminuye la TAN. La TAN de maíz en monocultivo y asociado con el pasto Toledo, presentó un comportamiento semejante en todo el periodo de evaluación, y llegó a su máximo valor de 0,32 g·m-2·día-1 a la edad de 30 días, lo que coincidió con un IAF de 2, para luego decrecer a valores negativos después de los 45 dds.
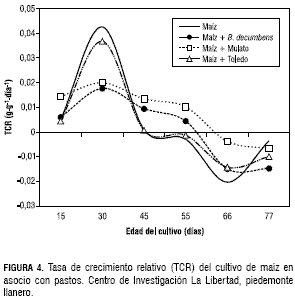
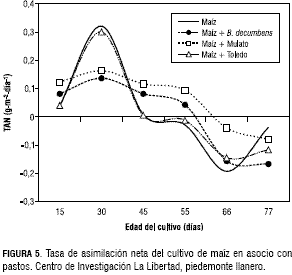
El maíz asociado con pasto Mulato y con el B. decumbens presentaron la misma tendencia, con una TAN a los 30 dds que estuvo entre 0,16 y 0,13 g·m-2·día-1, respectivamente. Después de los 66 dds, la TAN fue negativa con valores de -0,038 y -0,15 g·m-2· día-1 en el mismo orden (figura 5).
Índices fisiológicos en los pastos establecidos en monocultivo y asociados con maíz
Contrario a lo presentado en el crecimiento del maíz, los pastos sí mostraron diferencias significativas cuando estos se establecieron solos y asociados con maíz. El área foliar y la masa seca de los pastos asociados con maíz fue menor con respecto a los pastos establecidos sin maíz. El pasto Mulato sobresalió de los demás, por tener un establecimiento significativamente más rápido (P < 0,05) después de la siembra; en los primeros 37 días, este pasto presentó un área foliar de 1.908 cm2 y una masa seca de 9,4 g·m-2, mientras que cuando se sembró en asocio con maíz, el área foliar fue de 934 cm2 y la masa seca de 5,2 g·m-2. El otro pasto que también presentó un rápido establecimiento fue el Toledo, seguido por el B. decumbens; sin embargo, su establecimiento también fue más lento cuando se asoció con el maíz. En los primeros 37 días, los pastos sin maíz superaron en 1,6 veces el área foliar y en 2 veces la masa seca, a los pastos establecidos con maíz.
A los dos meses después de la siembra, los pastos Mulato y Toledo sin maíz presentaron un área foliar y masa seca superior (P < 0,05), con un promedio de 15.375 cm2 y 77,5 g MS por planta, respectivamente. Por su parte el B. decumbens desarrolló un área foliar de 4.350 cm2 y una masa seca de 20,3 g·m-2. En esta época el pasto Toledo en monocultivo presentó 3,4 veces más área foliar y 4 veces más masa seca, respecto al pasto Toledo + maíz. A los 75 dds el pasto Mulato aumentó su área foliar a 41.740 cm2, cifra superior a los demás tratamientos, incluyendo al pasto Toledo, que presentó un área foliar de 26.558 cm2, en tanto, la masa seca fue mayor en los pastos Mulato y Toledo con un promedio de 179 g·m-2. El crecimiento del pasto Mulato en monocultivo fue superior en los primeros 75 dds, mientras que los otros pastos en monocultivo y asociados con maíz se encontró que el área foliar a los 94 dds aumentó en más de dos veces respecto a la de los 75 dds. En los últimos 19 días de evaluación, se encontró que los pastos asociados con maíz presentaron un mayor crecimiento a lo cual contribuyó la senescencia del maíz que permitió mayor entrada de los rayos solares para ser aprovechados por los pastos (tabla 2).
La producción de masa seca de los pastos establecidos en monocultivo en los primeros 75 dds duplicó a la masa seca obtenida en los mismos pastos establecidos en asociación con el maíz. Aunque estas diferencias se redujeron a los 94 dds, la producción promedia de los pastos sin maíz y con maíz fue de 214 y 131 g·m-2 de MS, respectivamente (figura 6).En los tres pastos solos y asociados con maíz, el IAF fue en aumento hasta el día de la última evaluación, contrario a lo observado en el maíz en monocultivo y asociado con los pastos que por ser una especie semestral, después de los 55 dds, este parámetro empezó a descender. Este comportamiento del IAF el cual está estrechamente relacionado con la producción de biomasa favorece y justifica el establecimiento de la asociación de maíz-pastos, porque el cultivo de maíz no tiene ninguna competencia de los pastos asociados durante los primeros 30 días de crecimiento ni en los 20 días siguientes, importantes para la floración y formación de la mazorca (figura 7).
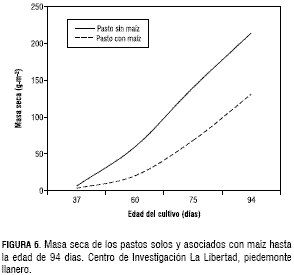
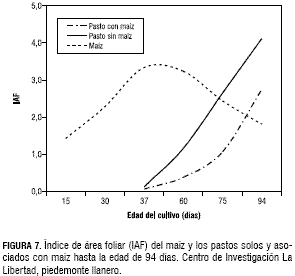
El maíz es una gramínea de rápida germinación y alto crecimiento en edades tempranas de la planta, mientras que los pastos en su mayoría germinan después de los 15 días y su desarrollo inicial es más lento, lo cual se confirmó con un IAF promedio de 0,071 y masa seca de 3,2 g·m-2 a la edad de 37 días. El maíz a los 45 dds presentó el mayor IAF con un valor de 3,5 y una masa seca de 29 g planta, cuando el pasto solo tenía un IAF de 0,4 y una masa seca de 20,3 g·m-2. Un comportamiento similar del IAF en la asociación de maíz con pastos, se presentó en condiciones de los Cerrados Brasileros, donde el maíz en sus estados de crecimiento iniciales no fue afectado por competencia con el pasto asociado. El mayor IAF del maíz fue a la edad de 50 días con un valor de 2, mientras que en esta misma fecha el IAF del pasto fue de 0,5; el cual aumento hasta 1,5 a la edad de 90 días, mientras que en este mismo momento el IAF de maíz también fue de 1,5 (Portes et al., 2001). El IAF de la asociación de maíz-pastos en las condiciones del piedemonte llanero, a los 90 dds fue de 1,8 para el maíz y de 2,7 para los pastos.
En esta misma época, los pastos establecidos sin maíz presentaron un IAF promedio de 3,5. La TCA del pasto Mulato en monocultivo y asociado con maíz presentó su mayor valor a los 75 dds con 8 g·día-1 y 5,2 g·día-1 de MS respectivamente. B. decumbens en monocultivo presentó el mismo comportamiento pero con una TCA más baja (2,8 g·día-1 de MS). El pasto Toledo solo y asociado con maíz presentó un TCA diferente a los otros dos pastos, porque a mayor edad se incrementó en orden ascendente, de forma que la TCA del pasto Toledo sin maíz fue de 3,3; 5,4 y 7,7 g·día-1 de MS a los 60, 75 y 94 días, respectivamente. Este mismo pasto asociado con maíz presentó una TCA de 0,7; 2,9 y 4,6 g·día-1 de MS en el mismo orden. En general, los pastos establecidos en monocultivo presentaron una mayor TCA que cuando estos se establecieron con el maíz (figura 8). La TCR de los pastos Toledo y Mulato establecidos sin maíz, presentaron un descenso al aumentar la edad desde los 60 hasta los 94 días (figura 9).
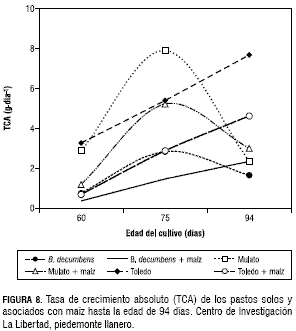
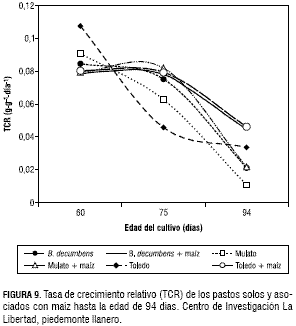
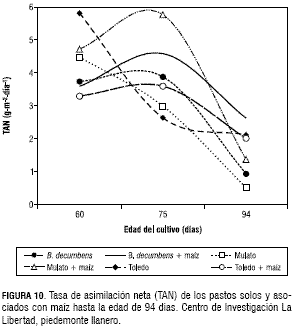
El pasto Toledo fue el de mayor TCR de MS a los 60 dds con 0,12 g·g-1·día-1 y a los 75 dds descendió a 0,045 g·g-1·día-1. Los pastos asociados con maíz y el pasto B. decumbens en monocultivo presentaron una TCR de MS similar, en 60 a 75 dds, con un valor promedio de 0,08 g·g-1·día-1. La TCR inicialmente presentó valores altos, los cuales fueron disminuyendo con la edad la cual se debe en parte al aumento gradual de tejidos no asimilatorios (Maldonado, 1993; Jarma et al., 1994). La TAN como indicador de la eficiencia fotosintética, mostró al pasto Toledo con el mayor valor a los 60 dds con 5,8 g·m-2·día-1, mientras que a la edad de 75 dds el máximo valor se obtuvo con el pasto Mulato asociado con maíz, cuyo valor fue de 5,7 g·m-2·día-1. Los pastos Toledo y Mulato establecidos en monocultivo disminuyeron la TAN desde la edad de los 60 días considerando que estos pastos cubrieron más rápidamente el suelo y la competencia entre plantas fue más temprano. El B. decumbens en monocultivo y los tres pastos asociados con maíz, presentaron un comportamiento similar de la TAN con un ligero incremento desde los 60 hasta los 75 dds, por efecto de la sombra del maíz y por el establecimiento más lento del B. decumbens (figura 10).
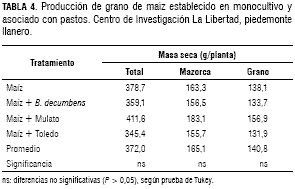
Producción de grano de maíz establecido en monocultivo y asociado con pastos
La asociación de maíz con pasto Mulato se caracterizó por presentar valores más altos de masa seca total y de grano, sin embargo, las diferencias observadas no fueron significativas (P > 0,05) cuando el cultivo se estableció solo o asociado con los tres pastos, de la que se obtuvo una producción de masa seca total promedia de 372 g/planta, 165 g/planta de mazorca y 140,8 g/planta de grano de maíz (tabla 4). Esta producción de grano era de esperarse, considerando que no se presentaron diferencias en los índices fisiológicos (IAF, TCC, TAN) en el cultivo de maíz establecido en monocultivo y en asociación con los pastos.
Conclusiónes
El crecimiento del maíz y la producción de grano no fueron afectados por el establecimiento en asociación con los pastos B. decumbens, Mulato y Toledo. El crecimiento de estos fue afectado cuando estos se establecieron en asociación con maíz. La producción de masa seca y el área foliar de los pastos establecidos en monocultivo en los primeros 75 dds duplicó la masa seca y área foliar obtenida en los mismos pastos establecidos en asociación con el maíz; por tanto, la asociación de maíz con pastos se constituye en una alternativa importante para el establecimiento o renovación de praderas en el piedemonte llanero, porque el lento crecimiento inicial de los pastos hace que el maíz se desarrolle sin competencia y no afecta el rendimiento de grano.
Agradecimientos
Se presenta un especial agradecimiento a Colciencias por la financiación de este trabajo de investigación, en el marco del proyecto “Potencial productivo y aspectos fisiológicos de los pastos tropicales bajo condiciones de manejo intensivo, como alternativa para recuperar praderas en suelos ácidos del piedemonte llanero”.
Literatura citada
Black, C.C. 1976. An assessment of C4 photosynthesis an productivity. En: Burris, R.H. y C.C. Black. (eds.). CO2 metabolism and plant productivity. University Park Press. Baltimore. [ Links ]
Clavijo, J.P. 1992. Análisis de crecimiento en malezas. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 15 p. [ Links ]
Duarte, M.J., E. Alexandrino y J.A. Comide. 2005. Duração do periodo de descanso e crescimento do dossel de Panicum maximum cv. Mombaça sob lotação intermitente. Rev. Bras. Zootec. 34(2), 398-405. [ Links ]
Hunt, R. 1982. Plant growth curves. The funtional approach to plant growth analysis. Edward Arnold, Londres. 48 p. [ Links ]
Jarma, A.O., C. Buitrago y S. Gutiérrez. 1994. Respuesta del crecimiento de la habichuela (Phaseolus vulgaris L. var. Blue Lake) a tres niveles de radiación incidente. Revista Comalfi 26(1-3), 62-67. [ Links ]
Kluthcouski, J. y H. Aidar. 2001. Implantaçao, conduçao e resultados obtidos com o Sistema Santa Fé. pp. 409- 441. En: Kluthcouski, J., L.F. Stone y H. Aidar(eds.). Integraçao lavoura-pecuaria arroz e Feijo. Embrapa, San Antonio de Goias. [ Links ]
Laffite, H.R. 2002. Fisiología del maíz tropical. Programa de maíz CIMMT. Tecnifenalce 2(7), 4-5. [ Links ]
Maldonado, G. 1993. Análisis de crecimiento de dos variedades de fríjol (Phaseolus vulgaris L.) cv. Cerinza cv. Tundama en condiciones de campo. Tesis de maestría. Universidad Nacional de Colombia, Bogotá. 453 p. [ Links ]
Medrano, H. y J. Flexas 2001. Fijación del dióxido de carbono y biosíntesis de fotoasimilados. pp. 173-201. En: Azcon-Bieto, J. y M. Talón (eds.). Fundamentos de fisiología vegetal. McGraw- Hill/Interamericana, Madrid. [ Links ]
Montaldi, E.R. 1995. Principios de fisiología vegetal. Ediciones Sur, La Plata, Argentina. 298 p. [ Links ]
Pereira, I. de O. y L.Y. Pacheco. 2001. Implantación y conducción del sistema Barreirao. pp. 265-303. En: Kluthcouski, J., L.F. Stone. y H. Aidar. (eds.). Integraçao lavoura-pecuaria. Embrapa arroz e Feijao, San Antonio de Goias. [ Links ]
Portes, T. de A., SI.C. Carvalho de y J. Kluthcouski. 2001. Aspectos fisiológicos das plantas cultivadas e analise de crescimento da Brachiaria consorciada com cereais. pp.305-329. En: Kluthcouski, J., L.F. Stone y H. Aidar (eds.). Integraçao Lavoura- Pecuaria. Embrapa arroz e Feijao, San Antonio de Goias. [ Links ]
Radford, P.J. 1967. Growth analysis formulae. Their use and abuse. Crop Sci. 7, 171-175. [ Links ]
Rincón, A., P.A. Cuesta, R.A. Pérez, G. Bueno, O. Pardo y J.E. Gómez. 2002. Producción y utilización de los recursos forrajeros en sistemas de producción bovina de la Orinoquía y el piedemonte caqueteño. Manual técnico. Plan de Modernización Tecnológica de la Ganadería Bovina Colombiana, Corpoica. 76 p. [ Links ]
Sans, J.I., R.S. Zeiger, S. Sarkarung, D.L. Molina y M. Rivera. 1999. Sistemas mejorados arroz-pasturas para sabana nativa y pasturas degradadas en suelos ácidos de América del Sur. pp. 232-244. En: Guimaraes, E.P., J.I. Sanz, I. Rao, M.C. Amézquita y E. Amézquita (eds.). Sistemas agropastoriles en Sabanas Tropicales de América Latina. Centro Internacional de Agricultura Tropical (CIAT), Cali y Empresa Brasilera de Pesquisa Agropecuaria (Embrapa). [ Links ]
Silveira, P.B., Honrad, M.L. y Pereira, M. 1999. Características morfológicas y fisiológicas relacionadas con el desempeño de arroz de secano asociado con Brachiaria brizantha. pp. 175- 195. En: Guimaraes, E.P., J.I. Sanz, I. Rao, M.C. Amézquita, y E. Amézquita (eds.). Sistemas agropastoriles en Sabanas Tropicales de América Latina. Centro Internacional de Agricultura Tropical (CIAT), Cali y Empresa Brasilera de Pesquisa Agropecuaria (Embrapa). [ Links ]

