










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.25 n.2 Bogotá jul./dez. 2007
Paola Cruz1, Kristell Acosta2, José Ricardo Cure3 y Daniel Rodríguez4
1 Bióloga, Universidad Militar Nueva Granada, Bogotá. u0500142@umng.edu.co
2 Bióloga, Universidad Militar Nueva Granada, Bogotá. u0500133@umng.edu.co
3 Docente, Universidad Militar Nueva Granada, Bogotá. jrcure@umng.edu.co
4 Docente, Universidad Militar Nueva Granada, Bogotá. daniel.rodriguez@umng.edu.co
Fecha de recepción: agosto 8 de 2007. Aceptado para publicación: octubre 1 de 2007
RESUMEN
El objetivo de este trabajo fue cuantificar variables asociadas al crecimiento y desarrollo de la planta de lulo bajo polisombra. La fase de campo se llevó a cabo en el municipio de Cajicá, Cundinamarca (2.597 msnm, 13 ºC y 80% HR). Se sembraron 210 plantas en un área de 900 m2; se realizó el seguimiento del cultivo desde la siembra hasta las primeras etapas de floración, y se efectuaron muestreos destructivos quincenales, de tres plantas seleccionadas aleatoriamente durante diez meses. Se determinó el peso seco de las diferentes estructuras de la planta a lo largo del crecimiento, así como la duración de las diferentes etapas fenológicas de dichas estructuras. Se estimaron las tasas de crecimiento exponencial y lineal para hojas (0,002702 g·g-1·dd-1 y 0,006096 g·dd-1), tallo principal (0,0036240 g·g-1·dd-1 y 0,05421 g·dd-1) y estructuras florales del racimo (0,004538 g·g-1·dd-1 y 0,0021270 g·dd-1); la tasa de producción de hojas (0,001119 h·h-1·dd-1) y de botones (0,037006 botones/dd) y, la duración de estados fenológicos como botón (59 dd), botón maduro (44 dd) y flor (44 dd) (dd = grados día, acumulación de unidades térmicas por encima de un umbral de crecimiento de 8 oC).
Palabras clave: estados fenológicos, tiempo fisiológico, parámetros de simulación.
ABSTRACT
The objective of this research was to quantify growth and development parameters of lulo plants under shading. The field experiment was carried out in Cajicá, Cundinamarca (2,597 m a.s.l., 13 ºC and 80% RH), where 210 plants were sowed in a 900 m2 area and sampling was done every two weeks during a ten month period, starting from planting up to first flowering stages. Three plants were taken randomly per sample and the dry weight of each plant organ was measured. Linear and exponential growth rates were obtained for leaves (0.002702 g·g-1·dd-1 y 0.006096 g·dd-1), main stem (0.0036240 g·g-1·dd-1 y 0.05421 g·dd-1) and flowers (0.004538 g·g-1·dd-1 y 0.0021270g·dd-1); production rates for new leaves (0.001119 h·h-1·dd-1) and new flowering structures (0.037006 flower bud/dd) were also estimated. Duration of phenological stages were calculated for flower bud (59 dd), mature flower bud (44 dd) and flowers (44 dd) (dd = degree days, termic units accumulation above a growing threshold of 8 ºC ).
Key words: phenological stages, physiological time, simulation parameters.
Introducción
El lulo (Solanum quitoense var. septentrionale) es un frutal originario de la zona central de Colombia, Panamá y Costa Rica (Heiser, 1972), que ha sido considerado por el Dane (2004) como una fruta agroindustrial promisoria, según el censo realizado entre el segundo semestre de 2003 y el primero de 2004. Ello se debe a su alto valor nutricional, sabor y apariencia, que como el de muchas solanáceas exóticas son muy apetecidas en Europa (Huyskens-Keil et al., 2001). En el país, la producción de la fruta refleja un incremento del 2,6% anual según los registros de 1995 hasta el 2000 (Torres, 2002), en parte porque se está empleando como alternativa a los cultivos ilícitos (Múnera, 2002) y porque su consumo a nivel agroindustrial es cada vez mayor (Morales et al., 2002). En el país se siembran cerca de 5.750 ha (CCI, 2006) en las que los agricultores se enfrentan a problemas agronómicos -como plagas y enfermedades y desarrollo vegetativo y reproductivo limitado- que se traducen en rendimientos bajos de las cosechas o necesidad de eliminar el cultivo antes de terminar el ciclo (Estrada, 1992). Estos problemas se manifiestan en la necesidad de importar la fruta desde Ecuador para suplir las demandas internas del producto. En este aspecto, las importaciones de lulo se han incrementado en 51,6% entre 1996 y 2000 (Torres, 2002). Para los cultivos de mayor importancia económica, muchos de los aspectos fisiológicos, ecológicos, el manejo de plagas y enfermedades han sido estudiados; sin embargo, la integración de esta información muchas veces se dificulta y su utilización es limitada, entre otras cosas, porque es producida de forma independiente y con metodologías no comparables (Cure, 1996).
El lulo no escapa de esta situación (Nacional Research Council, 1989). En este estudio se pretende obtener información básica sobre el crecimiento, desarrollo y fenología de la planta, para elaborar un modelo de crecimiento y desarrollo que permita simular, de forma integrada, la influencia de diversos factores sobre la productividad de la planta. La base teórica empleada corresponde al modelo de reserva metabólica (metabolic pool, Gutiérrez, 1996) el cual incluye los conceptos de adquisición y asignación de recursos en la planta conforme se describe a continuación. De acuerdo con Gutiérrez (1996), los procesos de crecimiento y desarrollo de plantas y animales pueden describirse y explicarse, en buena medida, en términos de jerarquía de asignación de recursos, que viene determinada genéticamente, y de adquisición de los mismos a partir del medio circundante, que básicamente constituye el efecto de los factores abióticos. La dinámica de este proceso se puede describir mediante el concepto de reserva metabólica. Este asume que la tasa de asimilación A de un individuo en la edad α, en el instante t y temperatura Τ, corresponde a la diferencia entre los recursos adquiridos S(•) y la respiración basal (z(•)), expresada por unidad de masa (respiración/gramo de masa). Así, (z(•)) M(t)) es la respiración del individuo completo, pues M(t) es la masa metabólicamente activa. Corrigiendo por los costos de conversión λ -de radiación a carbohidratos por ejemplo, en nuevos tejidos y órganos- arroja
donde los asimilados deberán ser distribuidos para el crecimiento vegetativo G(t) o reproductivo R(t).
Las demandas de respiración son prioritarias y parten de la base de la regla Q10 para poiquilotermos, en la que la tasa base de respiración (z10, intrínseca a cada especie) es doblada con cada incremento en 10 ºC en la temperatura Τ por encima del umbral de la especie θ
De esta forma, una vez adquiridos, los recursos son repartidos en las subunidades de la planta siguiendo en su orden prioridades de respiración, crecimiento reproductivo y vegetativo (Gutiérrez, 1996, Gutiérrez et al., 1998, 1987, 1975; Hopkins, 1995; Larcher, 2003, Salisbury y Ross, 1994; Wermelinger et al., 1991).
El modelo más simple que describe la asimilación de recursos en términos de acumulación de masa M en un tiempo t corresponde a:
El desarrollo fenológico se encuentra afectado por factores abióticos y bióticos. Dentro de estos, la temperatura en los organismos poiquilotermos tiene una gran incidencia, incrementando su duración cuando esta es baja y aumentando en caso contrario. Las unidades de tiempo fisiológico se denominan en grados día (dd, por sus siglas en inglés) y corresponden a unidades del tiempo fisiológico que se calculan en términos de temperatura acumulada en un tiempo específico sobre un umbral de temperatura que tarda un organismo poiquilotermo, una planta por ejemplo, en completar una etapa fenológica determinada (Gutiérrez, 1996) y se estima por la ecuación
Teniendo en cuenta el objetivo de largo plazo del presente estudio, que es desarrollar un modelo agroecológico para el manejo del lulo, el enfoque propuesto permitirá incluir otros parámetros de importancia como los que se refieren a aspectos fitosanitarios, que disminuyen la estructura vegetativa de la planta en diferentes fases de su crecimiento. De la misma forma, se pretende incluir variables de polinización dirigida, que tienen un efecto sobre el cuajamiento de los frutos y, en consecuencia, sobre la distribución de los asimilados entre las estructuras de la planta.
Materiales y métodos
LocalizaciónLa investigación se llevó a cabo en la Facultad de Biología de la Universidad Militar Nueva Granada, estación experimental Río Grande, ubicada en el municipio de Cajicá, Cundinamarca, a una altura de 2.597 msnm, 4º 56’ 42,8” norte y 74º 00’ 41,3” oeste, desde marzo hasta diciembre de 2005.
Material vegetal y condiciones del terreno
Se trabajó con 210 plantas de lulo S. quitoense var. septentrionale provenientes del municipio de Fusagasugá, Cundinamarca, y con 24 plantas de las mismas características, provenientes del Centro de Investigaciones y Asesorías Agroindustriales (CIAA) de la Universidad Jorge Tadeo Lozano. Ambos grupos de plantas fueron mantenidos en bolsas plásticas negras bajo condiciones de invernadero antes de ser sembradas en campo. El transplante al campo se hizo en un terreno de 30 x 30 m con las siguientes características climáticas: temperatura promedio anual diaria de 13 ºC, humedad relativa en torno a 80%, precipitación acumulada diaria de 2,90 mm bajo un patrón anual bimodal, promedio de radiación instantánea diaria de 135,74 W·cm2·s-1 y velocidad del viento promedio de 0,50 m·s-1.
El suelo incluyó preparación con incorporación de cascarilla de arroz, materia orgánica en forma de compost inicialmente, y antes de la siembra se aplicó una mezcla, en partes iguales, de fertilizante y cal (aproximadamente 30 g por 3 m2), según las recomendaciones del primer análisis de fertilidad del suelo. Después de la primera floración, se realizó un segundo análisis de fertilidad y de acuerdo con este, se realizaron fertilizaciones foliares y en corona. La distancia entre filas e hileras de plantas fue de 2 m y se sembraron a 0,2 m de profundidad El cultivo estuvo cubierto con polisombra negra al 33% (luminosidad 67%) en la parte superior a 2 m de altura sobre el suelo y polisombra blanca al 33% alrededor el cultivo, a fin de generar condiciones de luminosidad similares a las encontradas en el hábitat natural. Las plantas se podaron dejando cuatro tallos principales a partir de la primera ramificación, desde el primer cojín reproductivo. Debajo de este, todos los chupones fueron retirados. De la misma forma, se retiraron las hojas que por su peso u otras razones no se mostraban turgentes, evitando su posterior caída.
Metodología de muestreo
El muestreo se realizó en dos fases a fin de obtener las curvas completas de la fase exponencial y el primer ciclo reproductivo. La primera tuvo en cuenta las plantas provenientes de Fusagasugá y correspondientes a la fase de campo, y la segunda, a las plantas del CIAA, desde la germinación hasta la fase de campo. Inicialmente, 210 plantas de Fusagasugá, Cundinamarca, se sembraron en el lote experimental en marzo de 2005 bajo un diseño de muestreo estratificado que permitió considerar la pendiente existente en el terreno. Los muestreos se realizaron tomando al azar tres plantas quincenalmente, procurando que éstas estuvieran en competencia completa, es decir, con plantas que se encontraban al interior y no en el borde del terreno.
Se tomó una muestra aleatoria de tres plantas, una por estrato, quincenalmente, durante los ocho primeros muestreos. Con las plantas procedentes del CIAA se realizaron nueve muestreos destructivos durante cuatro meses, uno cada quince días, con tres repeticiones por fecha de muestreo. Los datos obtenidos de estas plantas fueron acoplados a los ya existentes de las plantas procedentes de Fusagasugá, de acuerdo con la masa asimilada y número de estructuras por planta de una fecha de muestreo a la siguiente, teniendo en cuenta que fueran consecutivas. Este proceso fue facilitado por la medición del tiempo en unidades fisiológicas de grados día. Así, el muestreo nueve de las plantas del CIAA fue reemplazado por el primero de las plantas de Fusagasugá ya que las masas y, aún más importante, el tiempo fisiológico, fueron equivalentes en este punto. A cada una de las plantas extraídas para los muestreos se les registraron las siguientes variables: altura de la planta, longitud de la raíz principal, número de hojas, número de ramas (incluyendo el tallo principal y los tallos secundarios), número de inflorescencias, número de flores, número de frutos, peso fresco y seco de las estructuras individuales de la planta, raíz, tallos, hojas, cojines florales, botones, flores y frutos. Igualmente, se estimó el área foliar a partir de la materia seca, la cual se transformó en área mediante el método tradicional del sacabocados. Adicionalmente, se estableció el patrón de ramificación y la ubicación de estructuras dentro de la planta mediante observación directa de las mismas en las diferentes fechas de muestreo.
Parámetros de crecimiento y desarrollo
A partir de los datos de materia seca, se obtuvieron parámetros de crecimiento como la acumulación de materia seca para hojas, tallos y estructuras reproductivas en forma de tasas de crecimiento. Estas tasas de crecimiento se estimaron con base en la metodología descrita por Gutiérrez (1996), la cual separa el crecimiento en dos etapas, la primera exponencial y la segunda lineal. La primera fase se estimó ajustando el siguiente modelo exponencial:
Para la fase lineal se empleó el modelo de regresión lineal:
donde M(t), es la masa de la estructura en un momento dado a, es la masa inicial γ, es la tasa de incremento de masa, y es la edad en tiempo fisiológico. Debido a que el crecimiento exponencial implica que la masa acumulada cada día depende a su vez de la masa acumulada en días previos, esta se expresa en g·g-1·dd-1. Por el contrario, en la fase de crecimiento lineal, la acumulación de masa es constante a través del tiempo y no es afectada por la biomasa acumulada previamente. Por esta razón, la tasa de crecimiento en la fase lineal se expresa en g·dd-1. Los parámetros calculados a partir de estos modelos fueron obtenidos empleando el programa R: A language and environment for statistical computing, desarrollado por R. Development Core Team (2006).
Se determinó la tasa y el tiempo que tardan en aparecer las hojas, tallos, flores y frutos en las plantas en base al modelo exponencial para hojas y lineal para las demás. Para las estructuras reproductivas, se emplearon cinco estadios de desarrollo, los cuales fueron descritos y de cada uno de ellos se tuvo en cuenta el tiempo de desarrollo. El umbral de temperatura empleado (8 ºC) fue una aproximación basada en las temperaturas presentes en el rango altitudinal en las que crece el lulo. A partir del umbral y la temperatura se obtuvo el tiempo fisiológico acumulado por la planta diariamente para cada una de las fechas de muestreo.
Factores abióticos
Se registraron diariamente las siguientes variables: temperatura máxima, mínima y promedio, humedad relativa, precipitación diaria acumulada, radiación diaria promedio y velocidad del viento con la estación meteorológica Metos compact/98 disponible en la estación experimental para las plantas cuando estas fueron transplantadas al campo. Durante la fase de invernadero, se registró únicamente la temperatura diaria con un termómetro de máximas y mínimas.
Resultados y discusión
Aspectos generales de crecimiento y desarrollo
La biomasa acumulada por la planta es distribuida entre las estructuras vegetativas y reproductivas según las prioridades (Gutiérrez et al., 1998, 1994, 1987, 1985; Gutiérrez, 1996; Wermelinger et al., 1991 y Villacorta et al., 1985). Como se presenta en la figura 1, la acumulación de masa en las diferentes estructuras o subunidades fue graficada en términos de tiempo fisiológico (grados día, dd), obteniendo el comportamiento esperado de las curvas para la fase evaluada, de acuerdo con el patrón estándar del crecimiento vegetal y que semeja la acumulación de materia seca obtenida por Medina (2003). Para analizar la tendencia del crecimiento adecuadamente, se tuvieron en cuenta factores circunstanciales abióticos y bióticos presentes en el ciclo del cultivo, que tuvieron un efecto marcado sobre su proceso. Así, los principales eventos ocurridos en su desarrollo se emplearon como patrón para dividirlo en diferentes periodos.
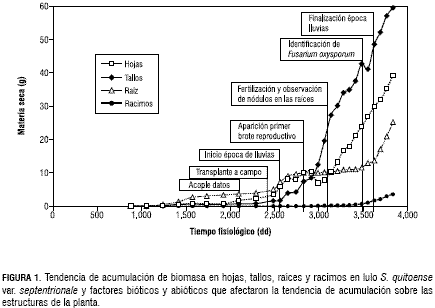
El primero de ellos tiene lugar desde el inicio de la toma de datos hasta alrededor de los 2.700 dd cuando inicia la época de lluvias (figura 1). Se caracteriza porque se producen el acople de los datos de las plantas provenientes del CIAA y los de las plantas procedentes de Fusagasugá a los 2.095 dd y posterior transplante a campo a los 2.300 dd. A continuación de este evento, se produce el transplante a campo y se observa crecimiento lento de las estructuras mientras las plantas se adecuan a las nuevas condiciones ambientales. Se debe anotar que el sustrato de las plantas durante la estancia en el invernadero no fue modificado y ello pudo ser un motivo para que las raíces no acumularan masa en este periodo (figura 1). El siguiente corresponde al periodo de inundación comprendido entre 2.700 dd y 3.700 dd, cuando finaliza la época de lluvias. Se denominó de esta manera debido a la alta precipitación que se presentó y a la textura arcillosa del suelo que en conjunto, provocaron inundación sobre el terreno. Durante este periodo es evidente la reducción en la acumulación de masa en las raíces y que coincide con la producción del primer brote reproductivo a los 2.828 dd (figura 1).
Después de este suceso, las hojas presentan una leve caída en la acumulación de materia seca pero se recupera rápidamente. La reducción se atribuye en términos generales, al efecto que tuvo la intensa temporada de lluvias y a la propagación de nemátodos. Ello es corroborado con los datos de precipitación, descartando así la posibilidad de un cambio en la distribución de los recursos como ocurre en otras especies cuando inician la etapa reproductiva (Gutiérrez, 1996; Gutiérrez et al., 1994, 1988, 1975 y Villacorta et al., 1985). A los 3.100 dd, de acuerdo con las recomendaciones de un análisis de fertilidad de suelos, se inició un programa de fertilización que incluyó la corrección de la concentración de manganeso, excesiva para ese entonces. A partir de ese momento, hojas, tallos y en su momento, estructuras reproductivas, incrementan la masa asimilada hasta finalizada la recolección de datos en campo. Las raíces sólo se recuperan luego de la culminación de la época de lluvias, cuando retoman el crecimiento. La identificación de Fusarium oxysporum y otros microorganismos fitopatógenos en el suelo cerca de los 3.500 dd, es un factor biótico que afectó al cultivo en épocas posteriores. Las estructuras vegetativas en la última etapa continúan en fase de crecimiento, por lo que para el periodo evaluado no se encontró un efecto evidente de la etapa reproductiva sobre la asignación de recursos.
Arquitectura de la planta
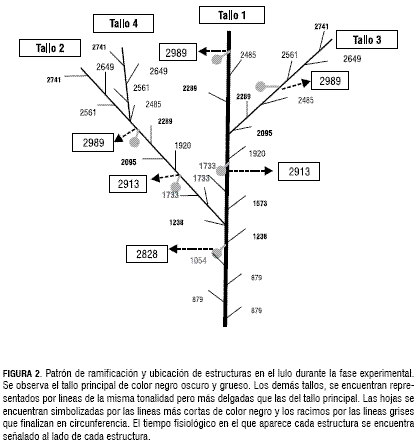
La arquitectura de la planta de lulo se explica según la ramificación y localización de estructuras en el tiempo. En términos generales, el primer brote reproductivo de la planta, el racimo 1 del tallo principal, se presenta a los 2.828 dd, una vez se han formado y desarrollado las cinco primeras hojas. Las siguientes estructuras reproductivas aparecen después de la primera una vez se han formado y desarrollado tres hojas más. El desarrollo de nuevos tallos tiene el siguiente comportamiento: se genera una nueva estructura cada seis hojas (o dos racimos) como se observa en la figura 2. Este patrón de crecimiento se repite de igual manera sobre los demás tallos. Esta secuencia de aparición de estructuras vegetativas y reproductivas, asegura una oferta de frutos constante en la planta ratificando su condición de planta perenne (Franco et al., 2002).
Parámetros de crecimiento y desarrollo
Los parámetros obtenidos hacen referencia a los tiempos de desarrollo, duración de los estadios y tiempo fisiológico de aparición de cada una de las estructuras, así como a las tasas de crecimiento, coeficiente de conversión de masa foliar a área foliar durante la etapa vegetativa y a los primeros estadios reproductivos del cultivo.

Parámetros vegetativos
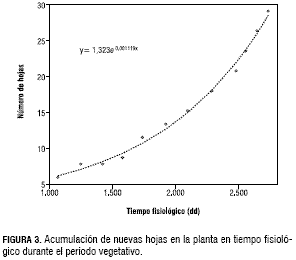
Se estimaron las tasas de acumulación de materia seca de hojas y tallos así como la tasa de aparición de hojas, los cuales se presentan en la tabla 1. La producción de hojas se ajusta apropiadamente a una curva de tipo exponencial (figura 3); así se obtiene una tasa de 0,001119 h·h-1·dd-1, indicando que no se alcanzó a incluir la fase de estabilización en el periodo evaluado. Después de la aparición de las primeras estructuras reproductivas (a los 2.828 dd), la tasa de aparición varía ampliamente entre plantas y entre muestreos, siendo evidencia del estrés al que las plantas fueron sometidas y que impidió la continuación del seguimiento de la aparición de hojas. Este comportamiento se asocia posiblemente a los eventos abióticos ya nombrados, en especial la época de inundación (desde finales de abril hasta mediados de octubre) que impactó sobre ciertas zonas del cultivo en función de la pendiente del terreno y la baja capacidad de drenaje del suelo. En relación al número de hojas obtenido por Medina (2003), se observa que existen grandes diferencias, ya que comparando el periodo evaluado hasta la época de reproducción, el acumulado es cercano a 100 hojas y el del presente estudio no supera las 40 hojas. Esta diferencia puede estar asociada a las diferentes características climáticas de las dos zonas, incluyendo las características de sombra del presente estudio.
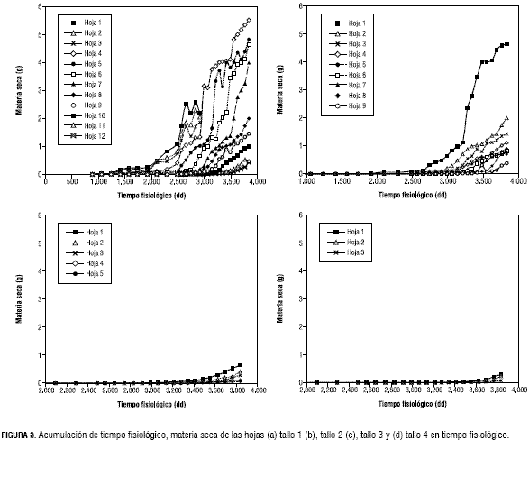
De acuerdo con la estructura establecida de arquitectura y como se ilustra en la figura 4, la acumulación de masa a través del tiempo en las hojas es muy similar en los diferentes ramos, por lo que se optó por hallar una sola tasa promedio de acumulación de masa de las hojas (tabla 1), para la fase exponencial 0,002702 g·g-1·dd-1 y lineal 0,006096 g·dd-1. En esta figura, es posible observar que las hojas 1, 2 y 3 del tallo principal, después de los 2.900 dd no siguen acumulando masa, ello se debe a que estas hojas fueron retiradas como parte de la poda de formación. Como se observa en la figura 4, la acumulación de materia seca en las hojas no se estabiliza y por las observaciones realizadas, el crecimiento se mantuvo hasta la terminación de los muestreos, motivo por el cual no fue posible determinar la longevidad de las estructuras vegetativas. La aparición de tallos es escalonada (figura 5). Este patrón implica que deben desarrollarse seis hojas antes de la formación de un nuevo tallo. La acumulación de materia seca en esta estructura, asume el comportamiento esperado, obteniéndose crecimiento exponencial en la primera fase para todos los tallos y una lineal en la segunda, para los tallos 1 y 2. Sin embargo, se observaron diferencias en las curvas de crecimiento entre estos, motivo por el que se asumieron dos curvas diferentes, una para el tallo principal y otra para los demás. Así, los valores de estas tasas fueron 0,0036240 g·g-1·dd-1 para la fase exponencial y 0,05421 g·dd-1 para la fase lineal para el tallo principal. Para los demás tallos, la tasa de crecimiento exponencial fue hallada a partir del promedio de los datos de acumulación de masa de los tallos 2, 3 y 4 siendo 0,003639 g·g-1·dd-1 y la lineal se obtuvo de la masa acumulada por el tallo 2, único tallo del que se obtuvieron suficientes datos para este segmento y cuyo valor es 0,025884 g·dd-1.
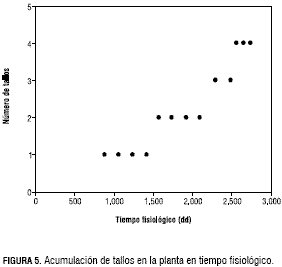
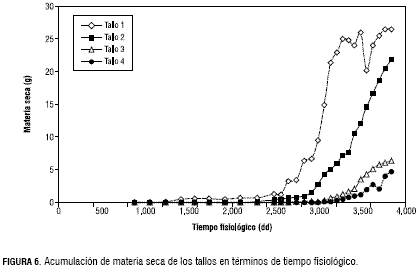
Como se observa en la figura 4, después de los 3.300 dd, la materia seca no se continúa acumulando a la misma tasa, esto se atribuye a que el tallo empieza a lignificarse. Parámetros reproductivos La estimación de los parámetros reproductivos (tabla 2), consiste en determinar los tiempos de aparición (figura 2) y duración de los diferentes estadios hasta el cuajado del fruto (fructificación). El periodo del experimento no permitió determinar la tasa de aparición de racimos ni la duración de los frutos.

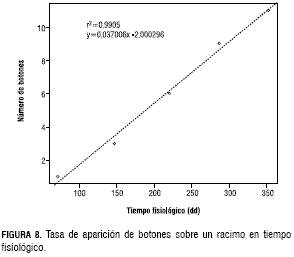
El primer cojín floral de la planta emerge a los 2.828 dd. Este estadio de “cojín floral” se caracteriza por ser una estructura uniforme, en la cual se observa únicamente una masa de tejido de coloración morada sobresaliendo del tallo. La duración de este estadio aumenta en forma constante según la aparición de los botones -en grupos de tres botones-, siendo el botón 1 el que permanece menos tiempo en forma de “cojín floral” y el botón 11 (según los datos obtenidos) el que tarda más tiempo en desarrollarse a este estadio; por ello, se estimó la tasa de aparición de botones en la planta que tiene un valor de 0,037006 botones/dd (figura 8). A esta ecuación se adiciona el tiempo fisiológico en el que aparece el cojín (tabla 2) y así se obtuvo la duración en el estadio de cojín para el botón deseado.
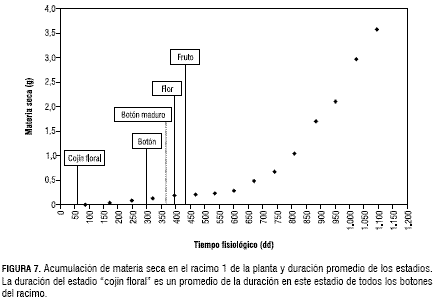
El estadio de “botón” tiene una duración promedio de 58 dd. En el estadio de “botón”, el cojín ya se ha diferenciado, y se pueden apreciar estructuras independientes de la misma coloración, la flor se encuentra totalmente cubierta por los sépalos que están rodeando los pétalos. La duración de este estadio en todos los botones del racimo es similar y ocurre desfasada en el tiempo. Una vez las estructuras reproductivas completan el estadio de “botón”, la maduración de estos se hace evidente, los sépalos inician su apertura gradualmente y se pueden distinguir con facilidad los pétalos de coloración blanca que están recubriendo las estructuras femeninas y masculinas de la flor. Este periodo se denominó “botón maduro” y tiene una duración promedio de 44 dd. Una vez el botón maduro se abre completamente, es evidente el paso al estadio de “flor”, en el cual los sépalos se abren y sostienen los pétalos de color morado en el envés y blanco en el haz. Se puede distinguir igualmente, el ovario de coloración amarilla, el estigma verde con el filamento amarillo y las anteras de color amarillo. La flor permanece en este estadio alrededor de 44 dd hasta que la fertilización se lleva a cabo y el fruto cuaja.
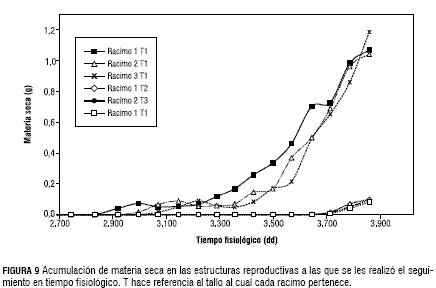
La acumulación de materia seca sobre los racimos es muy homogénea (figura 9), hecho que posibilita la observación de la fase exponencial claramente y una mayor uniformidad de la acumulación de biomasa en los racimos con respecto a la de biomasa en las hojas y demás estructuras vegetativas, ello se explica por la prioridad que tienen las primeras sobre las segundas (Gutiérrez, 1996, Villacorta et al., 1985).
Literatura citada
Angulo, R. 2003. Frutale s exóticos de clima frío. Bayer Crop Science, Bogotá. 136 p. [ Links ]
Azcón-Bieto, J. y M. Talón. 2001. Fundamentos de fisiología vegetal. Editorial McGraw Hill, Madrid. 536 p. [ Links ]
Bernal, J., O. Córdoba, G. Franco, M. Londoño, J. Rodríguez y N. Guevara. 1996. Cultivo del lulo (Solanum quitoense Lam.). pp. 61-80. En: Botero, R., G. Franco, M.J. Giraldo y D. Abad. (eds.). Memorias I Seminario de Frutales de Clima Frío Moderado. Centro de Desarrollo Tecnológico de Frutales, Manizales. [ Links ]
Cabezas, M. y D.L. Novoa. 2000. Efecto de la remoción de hojas y frutos en la relación fuente demanda en lulo (Solanum quitoense Lam.). pp. 69-75. En: Giraldo, M.J. (ed.). Memorias III Seminario de Frutales de Clima Frío Moderado, Centro de Desarrollo Tecnológico en Frutales, Manizales. [ Links ]
Corporación Colombia Internacional (CCI). 2006. Perfil del producto, inteligencia de mercados: lulo. En: http://www.cci.org. co, 15 p.; consulta: octubre de 2006. [ Links ]
Cure, J.R. 1996. Modelos de simulación en ecología aplicada. En: Memorias Seminario Internacional Modelos de Simulación: Su aplicación al estudio de los agroecosistemas y otras áreas de las ciencias básicas. Corpoica, Bogotá. [ Links ]
Davis, F. y C. Castaño. 1968. Crecimiento: Introducción a la fisiología de cultivos tropicales. Publicación miscelánea No. 9. pp. 1-18. [ Links ]
Departamento Administrativo Nacional de Estadística (Dane). 2004. I Censo Nacional de 10 Frutas Agroindustriales y Promisorias. Bogotá. 305 p. [ Links ]
Estrada I. 1992. Potencial genético del lulo (Solanum quitoense) y factores que limitan su expresión. Acta Hort. 310, 171-182. [ Links ]
Franco, G., J. Bernal, M. Giraldo, P. Tamayo, O. Cataño, A. Tamayo, J. Gallego, M. Botero, J. Rodríguez, N. Guevara, J. Morales, M. Londoño, G. Ríos, J. Rodríguez, J. Cardona, J. Zuleta, J. Castaño y M. Ramírez. 2002. El cultivo del lulo: manual técnico. Corpoica regionales 9 y 4, Colombia. 103 p. [ Links ]
García, J. 2003. Evaluación del crecimiento de dos ecotipos de lulo amazónico (Solanum sessiliflorum Dunal) bajo tres ambientes en el piedemonte amazónico del Caquetá. Tesis de maestría. Universidad Nacional de Colombia, Bogotá. 63 p. [ Links ]
Gómez, C., G. Franco y J.L. Gallego. 2003. Análisis de crecimiento de lulo La Selva (Solanum quitoense Lam.), en condiciones del departamento de Caldas. En: Memorias: Congreso Sociedad Colombiana de Fitomejoramiento y Producción de Cultivos. Universidad Nacional de Colombia, Bogotá. 57 p. [ Links ]
Gurevith, J., S. Scheiner y G. Fox. 2002. The ecology of plants. Sinauer Associates, Sunderland. 523 p. [ Links ]
Gutiérrez, A.P. 1996. Applied population ecology: A supply-demand approach. Jhon Wiley & Sons, Nueva York. 300 p. [ Links ]
Gutiérrez, A.P., L.A. Falcon, W. Loew, P.A. Leipzig y R. Van den Bosch. 1975. An analysis of cotton production in California: A model for alcala cotton and the effects of defoliators on its yields. Environ. Entomol. 4(1), 125-136. [ Links ]
Gutiérrrez, A.P., E.J. Mariot, J.R. Cure, C.S. Wagner Riddle, C.K. Ellis y A.M. Villacorta. 1994. A model of bean (Phaseolus vulgaris L.) growth types I-III: factors affecting yield. Agr. Systems 44, 35-63. [ Links ]
Gutiérrez, A.P., F. Schulthess, L.T. Wilson, A.M. Villacorta, C.K. Ellis y J.U. Baumgätner. 1987. Energy acquisition and allocation in plants and insects: a hypothesis for the possible role of hormones in insect feeding patterns. Can. Entomologist 119, 109-129. [ Links ]
Gutiérrez, A.P., A.M. Villacorta, J.R. Cure y C.K. Ellis. 1998. Tritrophic analysis of the coffee (Coffea arabica)- coffee berry borer [Hypothenemus hampei (Ferrari)]-parasitoid system. Anais da Sociedade Entomológica do Brasil 27, 357-386. [ Links ]
Gutiérrez, A.P., B. Wermelinger, F. Schulthess, J.U. Baumgätner, H.R. Herren, C.K. Ellis y J.S. Yaninek. 1988. Analysis of biological control of Cassava pests in Africa. I. Simulation of carbon, Nitrogen and water dynamics in Cassava. J. Appl. Ecol. 25, 901-920. [ Links ]
Heiser, C.B. 1972. The relationships of the naranjilla, Solanunm quitoense. Biotropica 4(2), 77-84. [ Links ]
Hernández M. y O. Martínez. 1993. Modelos de crecimiento para el fruto del lulo (Solanum quitoense Lam). Agricultura Tropical 30(3), 85-97. [ Links ]
Hopkins, W. 1995. Introduction to plant physiology. John Wiley & Sons, Nueva York. [ Links ]
Huyskens-Keil, S., H. Prono-Widayart, M. Schreiner y P. Ludders. 2001. Effect of surface coating and film packing on the keeping quality of solanaceous crops (Solanum muricatum Ait, Solanum quitoense Lam). Acta Hort. 553, 621-625. [ Links ]
Hunt, R. 1990. Basic growth analysis. Unwin Hyman, London. 112 p. [ Links ] Larcher, W. 2003. Physiological plant ecology. 4th edition. Springer, Berlín. 513 p. [ Links ]
Medina, C.I. 2003. Estudio de algunos aspectos fisiológicos del lulo (Solanum quitoense Lam.) en el bosque húmedo montano bajo del oriente antioqueño. Tesis de maestría. Universidad Nacional de Colombia, Medellín. 249 p. [ Links ]
Morales, J.E., F.J. López, J.C. Pérez, G. Ríos, D.I. Echeverri y M.A. Murillo. 2002. Evaluación agroeconómica del cultivo del lulo (Solanum quitoense Lam.) en la región central cafetera de Colombia. pp. 319-325. En: Giraldo, M.J. y J.P. Higuera. (eds.). Memorias IV Seminario de Frutales de Clima Frío Moderado. Corpoica, Universidad Pontificia Bolivariana, Centro de Desarrollo Tecnológico de Frutales, Medellín. [ Links ]
Múnera, G.E. 2002. Nemátodos asociados con el cultivo del lulo. pp. 135-141. En: Giraldo, M.J. y J.P. Higuera. (eds.). Memorias IV Seminario de Frutales de Clima Frío Moderado. Corpoica, Universidad Pontificia Bolivariana, Centro de Desarrollo Tecnológico de Frutales, Medellín. [ Links ]
National Research Council. 1989. Lost crops of the Incas: littleknown plants of the Andes with promise for worldwide cultivation. National Academy Press, Washington. pp. 267-275. [ Links ]
R. Development Core Team. 2006. A language and environment for statistical computing. En: http://www.r-project.org; consulta: octubre de 2007. [ Links ]
Salisbury, F.B. y C.W. Ross. 1994. Fisiología vegetal. Grupo Editorial Iberoamericana, México D.F. 759 p. [ Links ]
Torres, A. 2002. Inteligencia de mercados para lulo, mora y uchuva. pp. 326-330. En: Giraldo, M.J. y J.P. Higuera. (eds.). Memorias IV Seminario de frutales de clima frío moderado. Corpoica. Universidad Pontificia Bolivariana, Centro de Desarrollo Tecnológico de Frutales, Medellín. [ Links ]
Villacorta, A., A.P. Gutiérrez, W.J. Dos Santos y M.A. Pizzamiglo. 1985. Análise do crescimento e desenvolvimento do algodoneirono Paraná: um modelo de simulação para a variedade IAC-17. Pesquisa Agropecuaria Brasileira, Brasilia 20(1), 115-128. [ Links ]
Wermelinger, B., J. Baumgärtner y A.P. Gutiérrez. 1991. A demographic model of assimilation and allocation of carbon and nitrogen in grapevines. Ecol. Modeling 53, 1-26. [ Links ]

