










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.25 n.2 Bogotá jul./dez. 2007
Fánor Casierra-Posada1, María Constanza Cardozo2 y Julián F. Cárdenas-Hernández3
1 Profesor asociado, Facultad de Ciencias Agropecuarias, Grupo de Investigación Ecofisiología Vegetal, Universidad Pedagógica y Tecnológica de Colombia (UPTC), Tunja. fanor.casierra@uptc.edu.co
2 Ingeniera agrónoma, Universidad Pedagógica y Tecnológica de Colombia (UPTC), Tunja. connye80@yahoo.com
3 Estudiante de Ingeniería Agronómica, Universidad Pedagógica y Tecnológica de Colombia (UPTC), Tunja. julianc0182@yahoo.es
Fecha de recepción: julio 16 de 2007. Aceptado para publicación: octubre 1 de 2007
RESUMEN
El patrón de crecimiento es general en plantas anuales que se desarrollan en un ambiente óptimo; sin embargo, hay gran variación en la magnitud de los valores de peso seco, en la simetría de la curva en la escala de tiempo en que este sucede. En este artículo se presenta el análisis del crecimiento de tres cultivares de tomate (Lycopersicon esculentum Mill.) bajo condiciones de invernadero. Los cultivares evaluados fueron Sofia, Bravona y Granitio, en la localidad de Sutamarchán (Boyacá, Colombia). Desde el momento de la floración hasta la cosecha, se tomaron semanalmente, veinte frutos de cada cultivar, en los que se determinó el peso seco, el diámetro polar y transversal y los sólidos solubles totales; además, se calculó la tasa de crecimiento absoluto y relativo, y la relación diámetro polar/diámetro transversal. Se realizó un análisis de regresión para obtener los modelos de crecimiento. El peso seco, el diámetro transversal y polar mostraron una curva sigmoidea característica. Los sólidos solubles totales disminuyeron desde el inicio de las mediciones hasta el intervalo entre 29 a 36 días después de floración (ddf); después, los valores de esta variable se mantuvieron en aumento. La tasa de crecimiento relativo disminuyó fuertemente desde 8-22 ddf en los cultivares Sofía y Bravona, pero Granitio presentó una reducción menos drástica a lo largo del desarrollo del fruto. Los modelos de simulación obtenidos mostraron una alta correlación con los respectivos datos observados para el crecimiento del fruto.
Palabras clave: tasa de crecimiento relativo, tasa de crecimiento absoluto, sólidos solubles totales, cv. Granitio, cv. Bravona, cv. Sofía.
ABSTRACT
The growth patterns are quite similar in annual plants grown in an optimal environment; though, there is great variation in the magnitude of the dry weight values, symmetry of the curve, and time scale that it occupies. The growth analysis of three greenhouse-grown tomato cultivars was presented through this paper. Evaluated cultivars were Sofia, Bravona and Granitio that grew in Sutamarchán, Colombia. From bloom to harvest time, twenty fruits of each cultivar were taken weekly in order to determine dry weight, transversal and polar diameter, and total soluble solids; moreover, the absolute and relative growth rate, as well as the polar diameter / transversal diameter ratio were calculated. A regression analysis was done to obtain the growth models. The dry weight, transversal and polar diameter had a characteristic sigmoid shape. The total soluble solids decreased from the start of measurements to 29-36 time interval (days after bloom), subsequently, the values of this variable kept increasing. Relative growth rate is were strongly reduced from 8 to 22 days after bloom in Sofia and Bravona cultivars, but Granitio presented a less drastic reduction along the fruit development time. Obtained simulation outputs showed a good agreement with the observed data of fruit growth.
Key words: relative growth rate, absolute growth rate, total soluble solids, cv. Granitio, cv. Bravona, cv. Sofía.
Introducción
El cultivo de tomate a nivel nacional ocupó 0,6% del área total sembrada en cultivos transitorios, en 2006. Por su parte, en Boyacá se sembró, en el mismo año, el 10,4% del área total cultivada en tomate a nivel nacional, con un rendimiento de 45,7&m t∙ha-1, superado sólo por Caldas, cuyo rendimiento fue 47,5 t∙ha-1, lo cual es un índice de la importancia del cultivo para los agricultores boyacenses (Corporación Colombia Internacional, 2006). El rendimiento de un cultivo está determinado por la capacidad de acumular materia seca en los órganos destinados a la cosecha. Las hortalizas de fruto, cultivadas bajo condiciones de invernadero, se caracterizan, en su mayoría, por un crecimiento indeterminado, en donde los frutos son los principales órganos sumidero, los cuales compiten entre ellos y con los órganos vegetativos, por los asimilados disponibles (Peil y Gálvez, 2005). La distribución de materia seca puede cambiar durante el desarrollo de un cultivo, debido a cambios en la potencia de sumidero de un órgano individual y a alteraciones del número de sumideros que crecen en la planta (Marcelis, 1994). Asimismo, el crecimiento potencial de un órgano sumidero podría ser determinado durante el periodo de división celular, en la fase inicial de desarrollo de este órgano (Patrick, 1988).
El crecimiento es una aproximación holística, explicativa e integral, para comprender la forma y funciones de los vegetales (Hunt, 2003; Hunt et al., 2002). Este se manifiesta como un aumento irreversible de la masa de un organismo vivo, órgano o célula. El incremento en masa debe ser permanente, con lo cual se elimina la variación de volumen debida a fenómenos osmóticos. Existe, por tanto, el concepto de que el crecimiento puede definirse como un aumento en tamaño, sin embargo, esto no es siempre cierto, dado que puede darse el crecimiento sin que aumente el tamaño, pero sí, el número de células; por otro lado, también puede haber crecimiento con aumento de tamaño pero disminución del peso seco (Barceló et al., 1987; Salisbury y Ross, 2000).
Las curvas de crecimiento en los vegetales son un reflejo del comportamiento de una planta en un ecosistema particular con respecto al tiempo. Su elaboración es indispensable para la aplicación racional de las labores culturales en el momento adecuado, para garantizar una respuesta óptima del vegetal de acuerdo con nuestras necesidades y exigencias (Casierra-Posada et al., 2003; Hunt, 2003; Casierra-Posada et al., 2004). La elaboración de curvas de crecimiento permite conocer el modo en que crece la fruta con respecto al tiempo. La curva que representa el crecimiento de un fruto varía con la especie evaluada; así los frutos de las pomoideas son del tipo sigmoide simple (Casierra-Posada et al., 2003); en cambio la mayoría de las especies de carozo son del tipo doble sigmoidea (Coombe, 1976; Casierra-Posada et al., 2004).
En el caso de los frutos cítricos la forma dependerá de su origen, si es partenocárpico o sexual, pero en general es una sigmoidea (Agustí, 2000). Por su parte, el diámetro de los frutos del tomate crece también describiendo una curva sigmoide simple (Bertin, 2005). Las curvas de crecimiento del diámetro transversal en función del tiempo son las más empleadas por muchos autores. Tienen la ventaja de ser de fácil determinación, no destructivas y permiten el seguimiento a lo largo de la temporada. Estas curvas de crecimiento sirven para identificar no solamente la evolución del crecimiento en la temporada en las condiciones climáticas de la zona bajo estudio, sino también para estimar el peso que tendrá el fruto a cosecha (Coombe, 1976; Hunt, 1979). El análisis de crecimiento de plantas puede basarse directamente en la evolución cronológica de medidas, como peso seco, longitud de tallos, número de hojas y de ramas, etc. (Manrique, 1990). Con estas medidas es posible calcular la tasa de crecimiento relativo (TCR), la razón de peso foliar (RPF), la tasa de asimilación neta (TAN) y otras variables de importancia en la cuantificación del crecimiento (Ascencio y Fargas, 1973; Leopold, 1974; Hunt, 1990; Hunt et al., 2002). Mientras los primeros, tienen que ver con el desarrollo absoluto de la planta, los segundos explican su eficiencia en acumular materia seca como producto de sus procesos metabólicos (Geraud et al., 1995). A pesar de que los estudios básicos de crecimiento de órganos son fundamentales para el conocimiento de la fisiología del tomate, estos estudios son escasos en nuestro país, la información está dispersa o no es de fácil acceso. El objetivo del presente trabajo consistió en la cuantificación y establecimiento de modelos del crecimiento de frutos de tres cultivares de tomate, en función del tiempo, en plantas cultivadas bajo condiciones de invernadero.
Materiales y métodos
En el presente ensayo se evaluó el crecimiento de frutos en tres cultivares de tomate, mantenidos bajo condiciones de invernadero. Los cultivares estudiados fueron: Bravona, Sofía y Granitio, por ser materiales ampliamente cultivados en la región. El estudio se desarrolló en un cultivo comercial, en la zona rural del municipio de Sutamarchán (Boyacá), ubicado a 2.400 msnm. Para cada cultivar se marcaron 300 flores en igual estado de desarrollo, y luego del cuajado, se tomaron 20 frutos por cultivar para las respectivas evaluaciones. Las mediciones se hicieron hasta la madurez de los frutos. En los frutos, colectados semanalmente, se determinaron los diámetros polar y transversal, por medición con un nonio, así como la relación entre los dos diámetros, como el cociente de las dos mediciones; el peso seco del fruto, por secado al horno a 105 ºC, hasta peso seco constante; los sólidos solubles totales (SST), por lectura directa con un refractómetro manual. Con la información obtenida semanalmente, se calcularon las tasas de crecimiento absoluto y relativo, de acuerdo con la metodología propuesta por Hunt (1990). La información obtenida se analizó mediante el programa SPSS versión 11.5 con el que se establecieron los modelos de crecimiento y las ecuaciones de regresión para cada una de las variables. Para la selección del modelo, se tuvo en cuenta el coeficiente de regresión más cercano a la unidad.
Resultados y discusión
En todas las variables evaluadas se encontró diferencia altamente significativa (P < 0,01). En la mayoría de los casos se encontró que el modelo cúbico ofrecía el mayor coeficiente de regresión, con excepción de la tasa de crecimiento relativo en el cultivar Bravona, en la cual, el modelo exponencial fue el más acertado (tabla 1). El alto valor de los coeficientes de regresión es un índice de que los modelos encontrados estadísticamente se ajustan a los datos observados, con un mínimo de error. De esta forma es posible calcular el valor instantáneo de las variables, teniendo como factor variable (x) el tiempo después de floración.
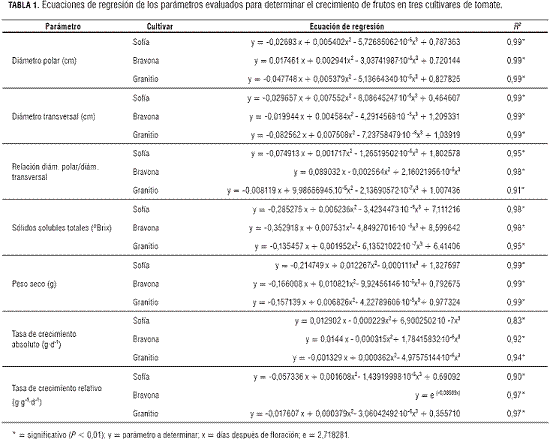
Los resultados mostraron que el incremento del diámetro transversal de los frutos de los tres cultivares describe curvas sigmoidales simples (figura 1), del mismo modo que el incremento del diámetro polar. Se presentó un incremento lento aproximadamente hasta 22 días después de floración (ddf); posteriormente, el crecimiento del diámetro de los frutos es más rápido hasta 50 ddf; luego, la intensidad del incremento disminuye hasta la maduración de los frutos. Por lo regular, los frutos, durante su desarrollo temprano, presentan tres fases: desarrollo del ovario, división celular y expansión celular (Gillaspy et al., 1993), lo cual se observa claramente en la figura 1, que representa el crecimiento en diámetro de los frutos. Inicialmente se tiene un crecimiento lento; después, el crecimiento se incrementa hasta disminuir su ritmo después de 50 ddf, hasta la cosecha, momento en el cual suceden los cambios fisicoquímicos que conlleva la maduración. A pesar de esto, no se observó una reducción en los valores registrados para el diámetro hacia el final, en el periodo de crecimiento de los frutos, como reportan Casierra-Posada et al. (2007) para un cultivar de tomate establecido a campo abierto. Esta diferencia radica en que a campo abierto los frutos están sometidos a una mayor deshidratación generada por los factores medioambientales como el viento, la radiación solar directa y los cambios en la humedad relativa; mientras que en cultivos mantenidos bajo condiciones de invernadero, el ambiente presenta una mayor estabilidad para los frutos y, por tanto, se previene la deshidratación de los mismos durante las fases tardías de su desarrollo.
Entre los factores que contribuyen significativamente al incremento en volumen de los frutos se encuentran los reguladores de crecimiento como las giberelinas, responsables de la expansión celular (García-Matínez y Hedden, 1997). De igual forma, Kojima (2005) encontró altas concentraciones de ácido giberélico en todos los constituyentes de los frutos de tomate durante la fase de expansión celular (frutos de 6 cm de diámetro), pero la concentración de GA1 fue menor cuando los frutos alcanzaban 8 cm de diámetro, lo que justifica la importancia de las giberelinas en la expansión celular en los frutos y, por consiguiente, en la ganancia en diámetro de los mismos. Por otro lado, el fruto presenta alto poder vertedero, lo cual significa que toma de la planta una serie de sustancias que transforma y convierte en sus componentes, siendo estos, moléculas de estructuras complejas y en principio muy diferentes a las que la planta toma del medio. Sin embargo, estas últimas son utilizadas, entre otros, para el crecimiento volumétrico del fruto (Barceló et al., 1987). Así, el crecimiento en diámetro de los frutos es un aumento irreversible como consecuencia del incremento en masa y número de las células.
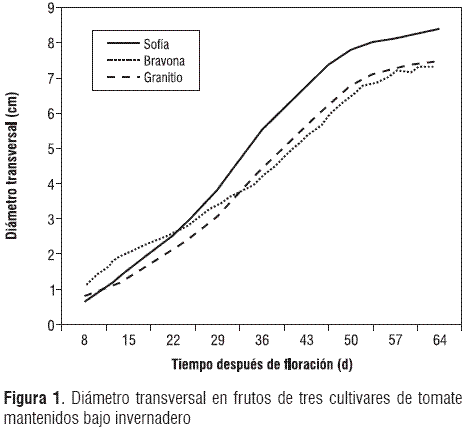
La relación diámetro polar-diámetro transversal mostró una reducción muy marcada en Sofía, hasta 15 ddf, mientras que el comportamiento de esta variable fue lo contrario para Bravona, durante el mismo periodo. Esto implica que en Sofía, hasta 15 ddf, los frutos son oblongos, pero en Bravona, los frutos son achatados; a partir de este momento, los frutos casi adquieren la forma característica del cultivar, hasta la maduración; mientras que en Granitio, la forma característica, en cuanto a longitud y ancho, está casi determinada desde el momento del cuajado del fruto (figura 2). La forma que adquiere el fruto durante los primeros estados de su crecimiento es un efecto de los reguladores de crecimiento producidos por las semillas inmaduras, de los cuales, el ácido giberélico cumple un papel relevante, como se mencionó anteriormente. Además, se ha encontrado que el ácido abscísico (ABA) está presente en altas cantidades en el pericarpo, en las axilas y en el tejido locular, en los estados tempranos de crecimiento de los frutos; sin embargo, posteriormente se encuentran grandes cantidades de ácido indolacético (AIA) que determinan tanto el ritmo del crecimiento como la forma del fruto (Kojima, 2005). Estos resultados sugieren que la expansión del tejido del pericarpo no es causado directamente por el AIA, sino por la actividad vertedero del fruto, ocasionada por las semillas en desarrollo (Varga y Bruinsma, 1976). Por otro lado, el ABA puede estimular la descarga de asimilados, transportados a través del floema, logrando así una promoción del poder vertedero de los frutos, durante sus fases tempranas de crecimiento (Brenner, 1989; Kojima, 2005).
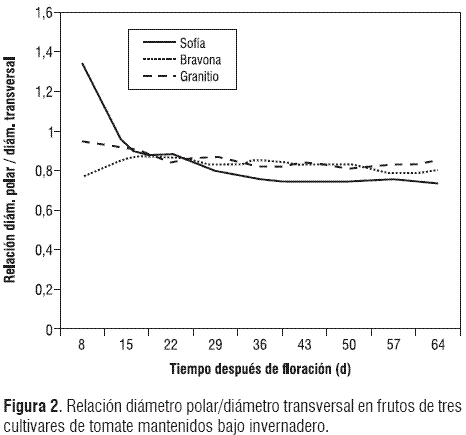
El contenido de sólidos solubles totales (sst) se redujo hasta 29 ddf en Sofía, mientras que el menor valor de esta variable se registró a los 36 ddf en Bravona. Luego, en los dos cultivares, se incrementan los sst hasta la maduración. Por su parte, Granitio no mostró una reducción tan drástica como los demás cultivares, sin embargo, mostró una reducción de los sst hasta 29 ddf, al igual que Sofía (figura 3). Las reacciones de hidrólisis que predominan en las macromoléculas presentes en frutos verdes, las rompen en monómeros que van a ser constituyentes importantes del sabor (Martínez-Barajas, 2003). El almidón se hidroliza para producir azúcares, responsables de la variación en el contenido de sólidos solubles durante todo el periodo de crecimiento y desarrollo de los frutos. El azúcar libre proviene principalmente de la glucosa y la fructosa, las cuales se encuentran en concentraciones similares en los frutos de tomate. Además, se discute que el contenido de azúcares en los frutos es altamente dependiente de la intensidad luminosa; por tanto, los tomates cultivados en campo abierto presentan mayores contenidos en sólidos solubles que aquellos que crecen bajo condiciones de invernadero (Herrmann, 2001).
Mientras Casierra-Posada et al. (2007) encontraron un incremento progresivo en el contenido de sólidos solubles totales en los frutos de tomate de plantas cultivadas a campo abierto, en el presente ensayo se observó un leve descenso en los valores registrados para esta variable hasta el intervalo entre 29 y 36 ddf, según el cultivar, diferencia que está relacionada con el sistema de producción en cuestión. Es así como a campo abierto las condiciones de luminosidad y transpiración favorecen un incremento constante de la concentración de sólidos solubles en los frutos (Herrmann, 2001), mientras que en invernadero, esta variable es altamente dependiente, no solo de la migración de fotosintatos a los frutos, acorde con su poder vertedero, sino también de la toma de agua realizada por los frutos, la cual diluye o concentra los SST. El comportamiento de la variable peso seco, mostró igual tendencia en los tres cultivares, una curva sigmoide simple; sin embargo, difirió en la magnitud de los valores, en cada lectura. Los cultivares Sofía y Bravona mostraron gran similitud en el patrón de acumulación de peso seco en los frutos, mientras que Granitio presentó valores considerablemente más bajos que los demás (figura 4).

Aunque la fotosíntesis en los frutos inmaduros de tomate no es despreciable (Czarnowski y Starzecki, 1992), sólo es responsable de una parte pequeña (menos de 10%) de la materia seca presente en la fruta (Ho y Grimbly, 1990). La mayoría de la materia seca de los frutos del tomate proviene de los fotoasimilados producidos en las hojas y transportados a los frutos, acorde con la relación fuente-vertedero. Los frutos representan vertederos de alto poder que, en sus estados iniciales de crecimiento, acumulan cantidades escasas de materia seca, cuando se tiene en cuenta el valor acumulado, pero este valor es alto, cuando se observa la tasa de crecimiento relativo. En los cultivares en estudio, los frutos empiezan a acumular grandes cantidades de materia seca a partir de 22 ddf, en forma sigmoide hasta el momento de la cosecha; por tanto, se debe tener en cuenta que dado que los frutos en desarrollo se encuentran en diferentes grados de madurez en la planta, la demanda por asimilados será constante, así que las labores orientadas hacia el suministro de nutrientes, agua y luz, especialmente, se deben realizar con gran prontitud y exactitud. Los tres cultivares evaluados mostraron diferentes patrones en cuanto a la acumulación instantánea de materia seca en g∙d-1 (figura 5).
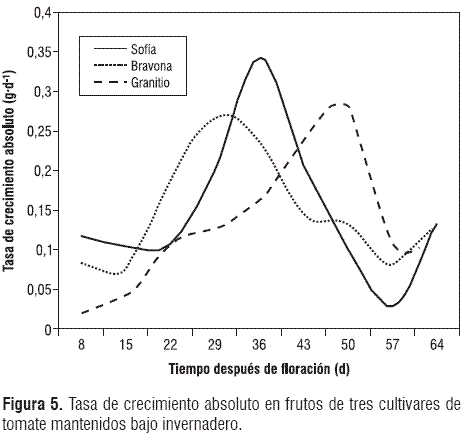
El máximo valor registrado de la tasa de crecimiento absoluto se presentó a los 29, 36 y 50 ddf, para Bravona, Sofía y Granitio, respectivamente. A partir de 57 ddf, los tres cultivares mostraron de nuevo, incremento de esta variable, hasta el momento de la cosecha. La tasa de crecimiento absoluto ofrece una visión acertada del poder vertedero de los frutos, en términos de acumulación de materia seca, medido de manera instantánea (Hunt, 1990; 2003; Casierra-Posada et al., 2007); por tanto, es importante destacar que las labores agrícolas orientadas a incrementar la acumulación de materia seca en los frutos, se deben llevar a cabo teniendo en cuenta los máximos valores registrados para la tasa de crecimiento absoluto, de acuerdo con cada uno de los cultivares en estudio, dado que en el momento en que se presenta un alto valor de esta variable es el periodo en que el fruto es sensible de influenciarse por actividades como la fertilización foliar, el suministro de agua e incluso por un mayor suministro de luz. En teoría, este planteamiento tendría gran validez, no obstante, se debe tener en cuenta que al tiempo se presentan en la planta frutos en diferentes grados de desarrollo a lo largo del tallo e incluso en un mismo racimo, por lo que las labores tendientes a incrementar el peso seco de los frutos deben realizarse de manera constante, una vez el primer racimo inicie su crecimiento y desarrollo (Casierra-Posada et al., 2007). El comportamiento de la tasa de crecimiento relativo mostró un descenso drástico desde el inicio de la toma de datos, hasta el intervalo 15-22 ddf en Sofía y Bravona; luego, continuó descendiendo paulatinamente hasta el momento de la cosecha. En el caso de Granitio, la reducción en los valores de esta variable no fue tan drástica como en los demás cultivares; de esta manera se presentó un descenso lento y casi uniforme hasta el momento de la cosecha.
Los altos valores encontrados en los primeros días de crecimiento de los frutos pueden relacionarse con el hecho de que el pedúnculo del fruto del tomate es rico en haces vasculares, con floema, a través del cual, los asimilados pueden migrar hacia el fruto en crecimiento. El contenido de ácido abscísico (ABA) en el pericarpo y lóbulos del fruto es muy alto, hasta cerca de 19 ddf, así que el ABA estimularía la descarga de asimilados en el fruto, lo que promueve la actividad vertedero del fruto de tomate en los primeros días de su crecimiento (Brenner, 1989; Kojima et al., 1993; Kojima, 2005), como ya se había anotado. Las diferencias encontradas entre los materiales evaluados en cuanto a cada una de las variables registradas, son la consecuencia de la expresión genética de los mismos. Si bien el fenotipo está influenciado por el sistema de producción, el componente genético es determinante en el comportamiento de los cultivares, como pudo evidenciarse en los resultados.
Agradecimientos
Este estudio se desarrolló con el apoyo de la Dirección de Investigaciones (DIN) de la Universidad Pedagógica y Tecnológica de Colombia, en el marco del plan de trabajo del grupo de investigación Ecofisiología Vegetal, adscrito al programa de Ingeniería Agronómica de la Facultad de Ciencias Agropecuarias.
Literatura citada
Agustí, M. 2000. Citricultura. Primera edición. Editorial Mundi- Prensa, Madrid. 161 p. [ Links ]
Ascencio, J. y J.F. Fargas. 1973. Análisis del crecimiento del fríjol (Phaseolus vulgaris L. var. Turrialba-4) cultivado en solución nutritiva. Turrialba 23(4), 420-428. [ Links ]
Barceló, C.J., G. Nicolás, B. Sabater y R. Sánchez. 1987. Fisiología vegetal. Ediciones Pirámide, Madrid. pp. 429-441. [ Links ]
Bertin, N. 2005. Analysis of the tomato fruit growth response to temperature and plant fruit load in relation to cell division, cell expansion and DNA endoreduplication. Ann. Bot. 95, 439-447. [ Links ]
Brenner, M.L. 1989. Hormonal control of assimilate portioning: regulation in the sink. Acta Hort. 239, 141-148. [ Links ]
Casierra-Posada, F., D.I. Hernández, P. Lüdders y G. Ebert. 2003. Crecimiento de frutos y ramas de manzano Anna (Malus domestica Borkh.) cultivado en los altiplanos colombianos. Agron. Colomb. 21(1-2), 69-74. [ Links ]
Casierra-Posada, F.; V.E. Barreto y O.L. Fonseca. 2004. Crecimiento de frutos y ramas de duraznero (Prunus persica L. Batsch, cv. Conservero) en los altiplanos colombianos. Agron. Colomb. 22(1), 40-45. [ Links ]
Casierra-Posada, F., M.C. Cardozo y J.F. Cárdenas-Hernández. 2007. Análisis básico del crecimiento en frutos de tomate (Lycopersicon esculentum Mill. cv. Quindío) cultivado a campo abierto. Documento en preparación.
Coombe, B.G. 1976. The development of fleshy fruits. Annu. Rev. Plant Physiol. 27, 207-228. [ Links ]
Corporación Colombia Internacional. 2006. Oferta agropecuaria ENA. Cifras 2006. Ministerio de Agricultura y Desarrollo Rural. Bogotá. pp. 14-62. [ Links ]
Czarnowski, M. y W. Starzecki. 1992. Spectral properties and CO2 exchange of tomato fruits. Photosynthetica 27, 513-519. [ Links ]
García-Martínez, J.L. y P. Hedden. 1997. Gibberellins and fruit development. pp. 263-285. En: Tomas-Barberan, F.A. y R.J. Robins. (eds.). Phytochemistry of fruit and vegetables. Oxford Sci. Publications, Heidelberg. [ Links ]
Geraud, F., D. Chirinos, M. Marín y D. Chirinos. 1995. Desarrollo de la planta de tomate, Lycopersicon esculentum Miller, cv. Río Grande en la zona del río Limón del Estado Zulia, Venezuela. II. Índice de crecimiento relativo, razón de peso foliar y gamma. Rev. Fac. Agron. 12, 15-23. [ Links ]
Gillaspy, G., H. Ben-David y W. Gruissem. 1993. Fruits: a developmental perspective. Plant Cell 5, 1439-1451. [ Links ]
Herrmann, K. 2001. Inhaltsstoffe von Obst und Gemüse. Ulmer Verlag, Stuttgart. pp. 73-89. [ Links ]
Ho, L.C. y P. Grimbly. 1990. The physiological basis for tomato quality. Grower 22, 33-36. [ Links ]
Hunt, R. 1979. Plant growth analysis: The rationale behind the use of the fitted mathematical function. Ann. Bot. 43, 245-249. [ Links ]
Hunt, R. 1990. Basic growth analysis. Plant growth analysis for beginners. Unwin Hyman, Boston. 110 p. [ Links ]
Hunt, R. 2003. Growth analysis, individual plants. pp. 579-588. En: Thomas, B., D.J. Murphy y D. Murray (eds). Encyclopaedia of applied plant sciences. Academic Press, Londres. [ Links ]
Hunt, R., D.R. Causton, B. Shipley y A.P. Askew. 2002. A modern tool for classical plant growth analysis. Ann. Bot. 90, 485-488. [ Links ]
Kojima, K. 2005. Phytohormones in shoots and fruits of tomato; Apoplast solution and seedless fruit. JARQ 39(2), 77-81. [ Links ]
Kojima, K., S. Kuraishi, N. Sakurai y K. Fusao. 1993. Distribution of abscisic acid in different parts of the reproductive organs of tomato. Sci. Hort. 56, 23-30. [ Links ]
Leopold, C. 1974. Plant growth analisis and development. McGraw- Hill Book Company, Nueva York. 466 p. [ Links ]
Manrique, L.A. 1990. Plant morphology of cassava during summer and winter. Agron. J. 82(5), 881-886. [ Links ]
Marcelis, L.F.M. 1994. Fruit growth and dry matter partitioning in cucumber. Ph.D. thesis. Wageningen Agricultural University, Wageningen. 173 p. [ Links ]
Martínez-Barajas, E. 2003. Análisis de la acumulación de azúcares en pericarpios de dos genotipos silvestres de jitomate (Lycopersicon esculentum). Agrociencia 37, 363-370. [ Links ]
Patrick, J.W. 1988. Assimilate partitioning in relation to crop productivity. HortScience 23, 33-40. [ Links ]
Peil, R. y J.R. Gálvez. 2005. Reparto de materia seca como factor determinante de la producción de las hortalizas de fruto cultivadas en invernadero. Rev. Bras. Agrociência 11(1), 5-11. [ Links ]
Salisbury, F.B. y C.W. Ross. 2000. Fisiología de las plantas. Desarrollo de las plantas y fisiología ambiental. Thompson Ediciones, Madrid. pp. 557-564. [ Links ]
Varga, A. y J. Bruinsma. 1976. Roles of seeds and auxins in tomato fruit growth. Z. Pflanzenphysiol. 80, 95-104. [ Links ]

