










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.26 no.1 Bogotá Jan./June 2008
FISIOLOGÍA DE CULTIVOS
Carlos Carranza1, Octavio Lanchero2, Diego Miranda3, Melba Ruth Salazar4 y Bernardo Chaves3
1 Ingeniero agrónomo, Universidad Nacional de Colombia, Bogotá. cecarranzag@unal.edu.co
2 Ingeniero agrónomo, Universidad Nacional de Colombia, Bogotá. jolancherod@unal.edu.co
3 Profesores asociados, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. dmirandal@unal.edu.co; bchavesc@unal.edu.co
4 Profesora, Facultad de Ingeniería, Universidad de la Sabana, Bogotá. melba.salazar@unisabana.edu.co
Fecha de recepción: mayo 23 de 2007. Aceptado para publicación: abril 9 de 2008
RESU MEN
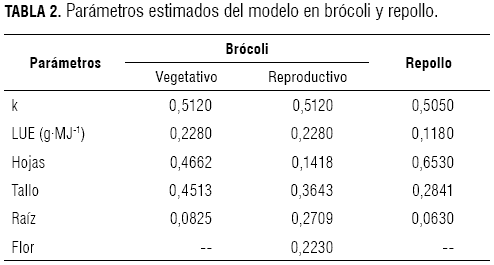
El experimento se realizó en el Centro Agropecuario Marengo de la Universidad Nacional de Colombia, ubicado en el kilómetro 12 vía Bogotá-Mosquera, coordenadas 4° 42’ N 74° 12’ W, a 2.516 msnm, temperatura media 13,7 ºC, precipitación 669,9 mm·año-1 y brillo solar 4,2 h diarias. En tres parcelas de 300 m2 cada una, con 8,3 y 11,1 plantas/m2 para brócoli y repollo respectivamente, regadas con aguas del distrito La Ramada, se realizaron muestreos cada ocho días, se midió el área foliar, el peso seco, la temperatura media y horas de brillo solar. Las horas de brillo solar se transformaron en radiación fotosintéticamente activa (PAR). Se usó un modelo matemático multiplicativo determinístico con los siguientes componentes: PAR, fracción de luz interceptada (FLINT), en función del índice de área foliar, del coeficiente de extinción (k) y uso eficiente de la luz (LUE). La masa seca total y su partición de acuerdo con los órganos de la planta, se simuló con el método de integración de Euler. Se estimó k igual a 0,5120 y 0,5052, LUE igual a 0,2289 y 0,1181 g·MJ-1 para brócoli y repollo, respectivamente. En brócoli, el 32,72% y 30% de la masa seca se distribuyó en el tallo; el 45,28% y el 30%, en las hojas; el 22% y el 17,83%, en la raíz en las etapas vegetativas y reproductiva, respectivamente. En repollo se distribuyó el 28,41% en el tallo; el 65,28%, en las hojas y el 6,31%, en la raíz.
Palabras clave: simulación, coeficiente de extinción, uso eficiente de luz.
ABST RACT
The experiment was carried out in the Agricultural Center Marengo of the National University of Colombia located on the km 12 route Bogotá-Mosquera with coordinates 4° 42’ N 74° 12’ W, at 2,516 m a.s.l. 13.7 ºC average temperature, 669.9 mm/year precipitation, 4.2 h daily light. Three plots with 300 m2 each one, with 8.3 and 11.1 plants/m2 for broccoli and cabbage, respectively, were applied with water of the district The Ramada. The samplings were realized every 8 days with 7 and 8 samplings for broccoli and cabbage, respectively. Leaf area, dry weight, hours of light and average temperature were measured. The light hours were transformed in photosynthetically active radiation (PAR). Deterministic multiplicative mathematical model was used taking into account the following components: Photosynthetic active radiation (PAR), fraction of intercepted light (FLINT), which depends on the leaf area index and, the coefficient of extinction parameter k, and the parameter light use efficiency (LUE). The entire plant dry weight and its partition among the plant organs were simulated by the method of integration of Euler. Parameters k as 0.5120 and 0.5052, and LUE as 0.2289 and 0.1181 g·MJ-1 were estimated for broccoli and cabbage, respectively. The percentage of dry weight distributed during the vegetative and reproductive stages were 32.72 and 30% in the stem, 45.28 and 30% in the leaves, and 22 and 17.83% in the root. In cabbage, the dry weight distribution was 28.41% in the stem, 65.28% in the leaves, and 6.31% in the root.
Key words: simulation, extinction coefficient, light use efficiency.
Introducción
El brócoli (Brassica sp.) pertenece a la familia Brassicaceae y se cultiva principalmente en Cundinamarca, Boyacá, Caldas y, en menor escala, en Antioquia y Nariño (Asohofrucol, 2007). En 2006, se sembraron 134 ha, de las que se cosecharon 106, para completar un volumen cosechado de 1.201 t (CCI, 2006).
El repollo (Brassica oleracea) pertenece a la familia Brassicaceae. Según la Encuesta Nacional Agropecuaria realizada por el Ministerio de Agricultura y Desarrollo Rural-CCI en 2006, en Colombia se sembraron 2.186 ha, de las que se cosecharon 2.150 ha para un volumen total de 50.767 t (CCI, 2006) y, en el 2007, se sembraron en la Sabana de Bogotá 500 ha con una producción anual de 14.457 t (Asohofrucol, 2007). Otras zonas productoras incluyen sectores de los departamentos de Cundinamarca, Boyacá, Antioquia, Nariño, Tolima y Caldas (Semillas Arroyave, 2006). El híbrido Delus es semitardío, se cosecha de 115 a 120 días después del transplante, tiene buena tolerancia a bajas temperaturas y es medianamente resistente a la salinidad (Semillas Arroyave, 2006).
Dentro de la clasificación americana de suelos, Soil Taxonomy, se estableció que, a partir de 2 dS·m-1, el suelo se considera salino y las propiedades morfológicas y fisicoquímicas del perfil quedan fuertemente influenciadas por el carácter salino. Por otra parte, el laboratorio de salinidad de los Estados Unidos estableció como límite 4 dS·m-1 para que la salinidad comience a ser tóxica para las plantas (Greenway y Munns, 1980; Marschner, 2002). Existe también un espectro continuo de tolerancia de plantas a condiciones salinas, que va desde las glicófitas, que son sensibles a la sal, hasta las halófitas, que sobreviven en concentraciones muy altas de sales (Volkmar et al., 1998).
Los modelos matemáticos son herramientas útiles para probar hipótesis, sintetizar el conocimiento, describir, entender sistemas complejos y comparar diferentes escenarios. Se usan como soporte para la toma de decisiones en sistemas, control climático de invernaderos, predicción y planeación de la producción (Lentz, 1998; Marcelis et al., 1998). Un modelo es una representación matemática de un sistema y la modelación es el proceso para desarrollar dicha representación; para que un modelo opere y produzca una imitación del mundo real, es necesario que se dé el proceso de simulación (Goudriaan y Van Laar, 1994). Penning de Vries et al. (1989) clasifican los modelos en descriptivos o explicativos. Descriptivos, estadísticos o empíricos son aquellos que usan regresiones y reflejan poco o nada los mecanismos que causan el comportamiento del sistema, mientras que los explicativos consisten en una descripción cuantitativa de esos mecanismos y procesos.
Así mismo, France y Thornley (1984) clasifican los modelos en: empíricos, cuando sirven para describir; mecanísticos, si describen con cierto grado de comprensión; estáticos, aquellos que no tienen en cuenta el tiempo, y dinámicos, aquellos en los cuales esta variable es explícita. Los modelos determinísticos predicen cantidades sin asociarles ninguna distribución probabilística, en cambio, los modelos estocásticos, que contienen elementos de azar, contemplan una distribución de probabilidad al hacer la estimación. En un modelo de simulación bien fundamentado de un cultivo es posible conocer el comportamiento de este ante variaciones ambientales y prácticas culturales, como respuesta a valores totales diarios u horarios de radiación solar, temperatura, lluvia o riego. También, se puede predecir cómo reacciona este a los manejos culturales, como densidad de siembra y fertilización, entre otros. Dependiendo del objetivo, la estructura y complejidad del modelo, se obtienen diferentes tipos de salidas y usos. Acerca del cultivo se puede obtener información sobre fenología, acumulación de masa seca por diferentes órganos de la planta y contenidos de nitrógeno en cada órgano, entre otros (Jones y Ritchie, 1990).
En Brassicas se han realizado trabajos en coliflor por Olesen y Grevsen (1997), quienes presentaron un modelo simple para el efecto de la temperatura y la radiación solar sobre la expansión del área foliar y la acumulación de masa seca. Con el uso de modelos de simulación dinámicos, también se describen los efectos del clima sobre la adaptación de las plantas y, por ejemplo, se puede determinar duración, en términos de la acumulación de grados día, el periodo de poscosecha en coliflor (Olesen y Grevsen, 2000).
Un modelo que incorpora efectos de ambiente tanto en crecimiento como en desarrollo del cultivo y que incluye la aclimatación de la planta se puede usar para predecir cosechas para diferentes condiciones ambientales (Olesen y Grevsen, 2000). Algunos modelos usan un parámetro de conversión de radiación solar a masa seca acumulada, los cuales no incluyen específicamente los procesos de fotosíntesis y respiración, ya que diversos estudios han mostrado que la relación de respiración a fotosíntesis es constante en muchos cultivos (Gifford, 1995; Dewar, 1996). Como lo demuestran los experimentos de Olesen y Grevsen (1997), para coliflor, y por Wheeler et al. (1993), para lechuga, el valor del coeficiente de conversión decrece con el incremento en la intensidad de la luz.
La función de demanda de asimilados se presenta no solo para regular su asimilación, sino que también sirve como mecanismo para su distribución en varios órganos. Muchos modelos usan un esquema simple en el que el coeficiente de partición depende de la edad del cultivo (Weir et al., 1984; Spitters et al., 1989). La partición de la masa seca es el resultado final del flujo de asimilados transportados desde los órganos fuente hasta los órganos de demanda (Marcelis, 1996).
Los modelos de simulación disponibles actualmente son específicos para las especies; la aproximación a la simulación depende del tipo de cultivo estudiado y del objetivo del modelo. El genotipo, el estado de desarrollo de la planta, diferentes condiciones de crecimiento y la regulación interna de la planta pueden afectar la partición de masa seca (Marcelis, 1996). La partición de la masa seca dentro de los órganos cosechables contribuye al rendimiento del cultivo, por lo que es importante hacer correctamente la simulación de la partición de la masa seca (Marcelis et al., 1998).
El objetivo de este estudio fue establecer un modelo simple de simulación de distribución de masa seca en brócoli (Brassica sp .) y repollo (Brassica oleracea) cultivados en la Sabana de Bogotá.
Materiales y métodos
Ubicación y material vegetal
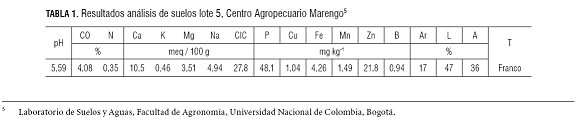
El ensayo se realizó en el Centro Agropecuario Marengo de la Universidad Nacional de Colombia, sede Bogotá, ubicado en el kilómetro 12 vía Bogotá-Mosquera, coordenadas 4° 42’ N 74° 12’ W, a 2.543 msnm, temperatura anual 12,6 ºC, precipitación 669,9 mm·año-1 y brillo solar 4,2 h diarias. Para cada especie se establecieron tres parcelas de 300 m2, con la siembra de plántulas de brócoli variedad Coronado a una distancia de 40 x 30 cm para una densidad de 8,3 plantas/m2 y para repollo híbrido Delus de 30 cm x 30 cm, para una densidad de 11,1 plantas/m2. La fertilización se realizó a partir del análisis de suelo (tabla 1), los nutrientes faltantes se suplementaron con fertilizantes químicos edáficos. El suelo presentó una conductividad eléctrica de 2,16 dS·m-1 y el agua de riego, 0,95 dS·m-1. El manejo agronómico de malezas, plagas y enfermedades se efectuó oportunamente, dependiendo de las poblaciones, niveles y síntomas. Por último, no hubo problemas relevantes que afectaran el ciclo de desarrollo de las plantas.
Muestreos material vegetal
Los muestreos se realizaron cada ocho días en tres plantas por parcela, para un total de siete en brócoli y ocho en repollo. El área foliar se determinó mediante un acumulador LI-COR® 3000a (LI-COR, Estados Unidos), se registraron datos de número de hojas por planta, peso fresco y seco de hojas, tallo y de raíz, mediante su secado en un horno a 60 °C durante 72 h, además se registraron variables de clima, como horas de brillo solar y temperatura media, en la estación meteorológica de C.I. Corpoica Tibaitatá entre los meses de septiembre y noviembre de 2006.
Componentes del modelo
Para la generación del modelo, las horas de brillo solar se transformaron en radiación fotosintéticamente activa (PAR en MJ), según metodología propuesta por Angstrom (1924). Se usó un modelo matemático multiplicativo determinístico con estos componentes: radiación fotosintéticamente activa (PAR), fracción de luz interceptada (FLINT), en función del índice de área foliar y de los parámetros coeficiente de extinción de luz (k) y uso eficiente de luz (LUE).
Tasa de crecimiento diario del cultivo (g·m-2·d-1)
Se determinó, con la siguiente ecuación propuesta por Monteith (1996), Gosse et al. (1986) y Kooman (1995):
donde:
Wt: es el incremento de la masa seca total de la planta (g·m-2) en el tiempo t.
FLINTt =(1-e-k·LAI), es la fracción de luz interceptada en el tiempo t; k es el coeficiente de extinción de la luz y LAI es el índice de área foliar del cultivo.
PARt: es la radiación fotosintéticamente activa en el tiempo t (MJ·m-2·d-1).
LUE: es el uso eficiente de la luz (g·MJ-1).
Simulación
La simulación es la integración numérica de la tasa de crecimiento diario del cultivo. Para obtener la masa acumulada en el tiempo t (Wt), se usó el método de Euler con Δt=1 (día).
La estimación de los parámetros k y LUE se llevó a cabo a través del método de optimización no lineal usando SOLVER de Excel. Este método iterativo minimiza la suma de cuadrados de la diferencia entre el valor observado y el valor simulado.
Modelo de distribución de masa seca
Los coeficientes de distribución αo son las proporciones de la masa seca total que se asignan a hojas, tallos y frutos, dependiendo del estado de desarrollo vegetativo o reproductivo (Marcelis, 1994; Kooman, 1995). En repollo, la distribución de la masa seca total se llevó a cabo en los órganos: hojas, tallo, raíz. En brócoli, se tuvieron en cuenta los estados vegetativo y reproductivo y se modeló la masa seca de hojas, tallo, raíz y flor, respectivamente.
La masa seca de cada órgano se simuló con la ecuación de Euler, así:
donde:
Wot: es masa seca total de un órgano: hojas (Wlt), tallos (Wst), flores (Wft) o raíz (Wrt) en el día t (g·m-2·d-1).
Wo (t-1): es la masa seca de hojas (Wlt-1), tallos (Wst-1), flores (Wft-1) o raíz (Wrt-1) en el día t-1 (g·m-2·d-1).
αo: es el coeficiente de distribución de hojas (αl), tallos (αs), flores (αf) o raíz (αr) y Σαo=1.
δWt: es la tasa de crecimiento diario de masa seca total (g·m-2·d-1).
∆t: es el incremento a través del tiempo (1 día).
Resultados y discusión
Radiación fotosintéticamente activa y fracción de luz interceptada
La radiación fotosintéticamente activa (RFA) durante los meses de septiembre-noviembre de 2006 en el Centro Agropecuario Marengo (figura 1) presentó una fluctuación entre 9,3 y 22,8 MJ·m-2·día-1.
En modelos basados en fotosíntesis, primero se calcula la intercepción de la luz por el área foliar para simular la producción de fotosintatos; posteriormente, se calculan los fotosintatos usados para la respiración, la conversión a masa seca estructural, la partición de asimilados o masa seca de los diferentes órganos de la planta y, por último, el peso fresco se puede estimar a partir del peso seco (Marcelis et al., 1998).
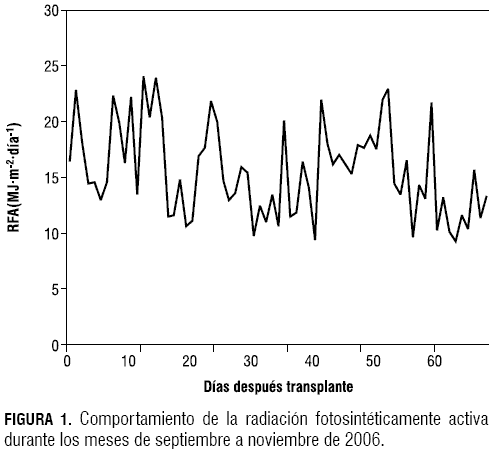
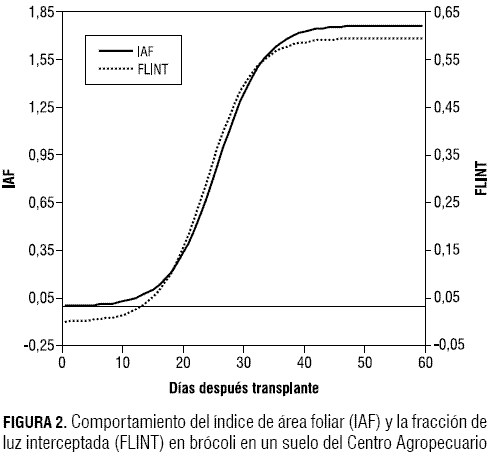
En brócoli, 50 días después del transplante (ddt) se presentó el máximo índice de área foliar (IAF) igual a 1,76, menor 2,3 veces al máximo encontrado en estudios realizados en diferentes densidades de brócoli por Francescangeli et al. (2006), donde a una densidad de 8 plantas/m2 el IAF fue igual 4,04; la FLINT fue de 0,7 a los 84 ddt, superior a la máxima encontrada en esta investigación, que fue de 0,59, constante desde los 44 días hasta el final del ensayo (figura 2). En repollo, a los 68 ddt, se alcanzó un IAF de 5,17 y la FLINT fue igual a 0,92 (figura 3).
A pesar de la importancia del desarrollo del área foliar sobre el crecimiento del cultivo, su estimación es imprecisa, aspecto que se considera débil en el estudio de modelos de crecimiento, así mismo, no se ha entendido realmente qué controla la tasa de desarrollo foliar. En varios estudios no se ha simulado el área foliar o IAF; sin embargo, contribuye mucho a la estructuración del modelo. Salazar et al. (2008), en un estudio de modelación de la biomasa de uchuva (Physalis peruviana L.), simuló el IAF a partir del área foliar específica y la masa seca foliar. El desarrollo del área foliar se puede ver como la combinación de cuatro procesos: tasa de emisión foliar, tasa y duración de expansión de hojas individuales y duración de hojas individuales (Marcelis et al., 1998).
En cultivos hortícolas se presentan valores promedio del coeficiente de extinción de la luz (k) en un rango de 0,5 a 0,8. El coeficiente de extinción depende de la inclinación de los rayos de luz incidente, la posición y la orientación de las hojas. De Wit (1965) y Ross (1981) han desarrollado modelos más detallados en los que se incluyen otros aspectos que no se consideran usualmente en la estructuración de modelos. En este trabajo en brócoli y repollo se estimó k igual a 0,5120 y 0,5052, respectivamente (tabla 2).
El coeficiente de extinción para brócoli es mayor que el obtenido en estudios realizados por Olesen y Grevsen (1997), en coliflor y brócoli encontraron valores de 0,55 y 0,45, respectivamente. Se asume que el coeficiente de extinción usualmente depende del tipo de planta, según los estudios realizados por Weir et al. (1984), en trigo, Graf et al. (1990), en arroz, y Steer et al. (1993), en girasol; y presenta pocas variaciones durante el ciclo del cultivo. Sin embargo, Yunusa et al. (1993), en trigo, Imai et al. (1994), en Canna edulis, y Kubota et al. (1994), en Pennisetum purpureum, evidenciaron que el coeficiente de extinción cambió con la edad de la planta en los cultivos evaluados.
Las hojas de brócoli son más anchas en el ápice que en la base, esto incrementa la captura de la radiación fotosintéticamente activa debido a que se presenta menor agrupación de hojas cerca al tallo y el área foliar es uniforme (Francescangeli et al., 2006). Los pecíolos de las hojas también incrementan la intercepción de la radiación solar, debido a la reducción de los aglomerados de las hojas alrededor del tallo (Teh, 2004). En cualquier posición dentro del dosel de la planta, la radiación fotosintéticamente activa está conformada de contribuciones desde todas las direcciones, que se agrupan de acuerdo a su dirección zonal y acimut (Marcelis et al., 1998). El ángulo de incidencia de la radiación en una hoja depende de la orientación de la hoja y la dirección de la radiación.
En repollo, al inicio del cultivo, las hojas tienen IAF menores a 2. En esta etapa las hojas son más erectófilas y se presenta mayor penetración de la radiación fotosintéticamente activa a través del dosel de la planta. A medida que el IAF alcanza su máximo (5,17) a los 68 ddt, las hojas del tercio superior e intermedio se compactan formando la cabeza, que finalmente se cosecha; las hojas del tercio inferior captan parte de la radiación directa y los tercios superiores captan radiación difusa. En valores bajos de IAF, las hojas forman ángulos adecuados para interceptar la luz, con lo que se incrementa el valor del coeficiente de extinción hasta cerca de la unidad. Por otra parte, en doseles escasos, las hojas se pueden presentar formando racimos, con lo que se reduce el coeficiente de extinción. No obstante, para estas especies hortícolas el valor del coeficiente de extinción se encuentra por debajo de 0,7 según Marcelis et al. (1998).
De acuerdo con trabajos realizados por Gijzen (1995) a nivel del dosel, la captación de la radiación es mayor debido a su dispersión, que hace que las hojas puedan tener múltiples oportunidades para captarla. Para un IAF superior a tres unidades, aproximadamente el 90% de la PAR la absorbe el dosel.
Uso eficiente de la luz (LUE )
A través de modelos empíricos es posible estimar la producción de masa seca como resultado de la radiación interceptada y empíricamente se derivan valores de LUE (Monteith, 1977; Gallagher y Biscoe, 1978; Jones y Kiniry, 1986; Williams et al., 1989). En modelos de crecimiento de cultivo debido básicamente a su simplicidad y a la evidencia experimental disponible, el LUE es una aproximación para calcular la conversión de MJ (Mega Joule) a gramos de masa seca total. Monteith (1977), Gallagher y Biscoe (1978) y García et al. (1988) aseguran que la relación entre el tiempo integral de intercepción de la radiación y la producción de masa seca es constante. Análisis teóricos realizados por Hammer y Wright (1994), Dewar (1996), Haxeltine y Prentice (1996), Dewar et al. (1998), Medlyn (1998) y Kage et al. (2001a) presentan al LUE, en un amplio rango de días, como equivalente a la suma de la radiación fotosintéticamente activa, en un efecto combinado de la adaptación del aparato fotosintético a la radiación ambiental dentro del dosel y en el tiempo (Kage et al., 2001b). Para las condiciones de este experimento, en brócoli se estimó un LUE de 0,2289 g·MJ-1 y en repollo, de 0,1181 g·MJ-1 (tabla 2). Estos valores son inferiores comparados con otros cultivos hortícolas como coliflor donde Olesen y Grevsen (1995, 1997) encontraron valores de 3,2 y 4,1. Para otras especies, como el pasto ryegrass, se obtuvo un LUE de 2,01 g·MJ-1 (Akmal y Janssens, 2004); en cereales, un rango de 1 a 4 g·MJ-1 (Curt et al., 1998); en maíz, 1,6 g·MJ-1 y sorgo, 1,25 g·MJ-1 (Muchow y Davies, 1988); en cebada, 1,4 g·MJ-1 (Monteith, 1969); sin embargo, pueden ocurrir pequeñas variaciones de LUE en plantas de la misma especie, establecidas en un mismo sitio de plantación, como consecuencia de variaciones microclimáticas (Kage et al., 2001a).
Distribución de la masa seca
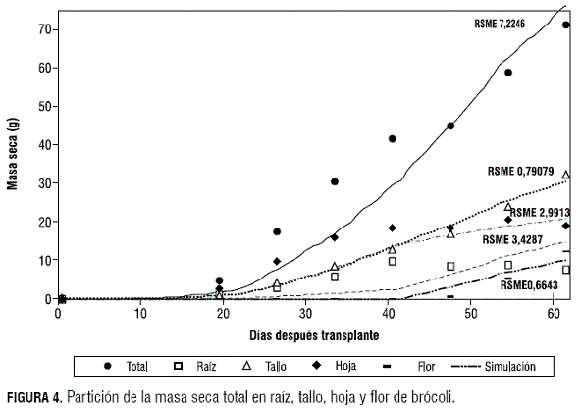
Durante las etapas vegetativa y reproductiva, la masa seca se distribuyó respectivamente así: en brócoli, el 45,13% y 36,43%, en el tallo; el 46,62% y el 14,18%, en las hojas; el 8,25% y el 27,10%, en la raíz. En repollo se distribuyó el 28,41% en el tallo, el 65,28% en las hojas y el 6,31% en la raíz (figura 4).
Algunos autores dicen que existe bastante información sobre los progresos en la cuantificación y modelación de la partición de masa seca en frutas (Wermelinger et al., 1991; Grossman y DeJong, 1994) y hortalizas (Dayan et al., 1993a,b; Marcelis, 1994; De Koning, 1994; Heuvelink, 1996). El genotipo, la etapa de desarrollo de la planta, las condiciones ambientales para el crecimiento y la regulación interna de la planta también pueden afectar la partición de la masa seca (Marcelis, 1996).
La masa seca en los órganos cosechables contribuye al rendimiento del cultivo, esto indica la importancia de simular correctamente la partición de la masa seca. Sin embargo, para plantas pequeñas (jóvenes) de crecimiento rápido, la partición de la masa seca dentro de las hojas es importante, porque una fracción grande de luz todavía no es interceptada por las hojas. Además, en muchos cultivos, como hortalizas y frutas, donde los órganos cosechables se producen durante un periodo extenso, se debe mantener un balance óptimo entre la partición en los órganos cosechables, la productividad a corto plazo y la otra parte de la planta (partes vegetativas: la futura capacidad de producción) (Marcelis et al., 1998).
Conclusiones
• A pesar de las diferencias encontradas con otros autores, los valores de la raíz cuadrada del cuadrado medio del error (RSME), tanto para la biomasa total como para las particiones en los diferentes órganos, demuestran el buen ajuste de los modelos generados.
• Para brócoli, durante el cambio de la fase vegetativa a la reproductiva, algunas de las estructuras que actúan como demandas (tallos y hojas) redistribuyen sus asimilados hacia los órganos reproductivos en formación y, posteriormente, para llenado. Es así como la planta redistribuye el 32% de los fotoasimilados de la hoja, el 9% del tallo (estas estructuras actúan como fuentes) y estos asimilados a su vez son distribuidos a la raíz (19%) y la flor (22%).
• En repollo, más del 65,3% de la biomasa se acumuló en las hojas -que es la parte comercializable-, un 28,4%, en el tallo y el 6,3%, en la raíz.
• No es posible extrapolar este modelo a otras condiciones, debido a las particulares características tanto del suelo como del riego, sin embargo, es posible usarlo para las características ambientales del área cubierta por el distrito de riego La Ramada -zona más representativa de la producción hortícola en la Sabana de Bogotá-.
• Futuros trabajos deben incluir otras condiciones ambientales, el análisis de sensibilidad de los parámetros estimados y la validación de estos modelos. A largo plazo, se deben modelar los aspectos relacionados con el suministro de agua y nutrientes y con el manejo de plagas y enfermedades.
Literatura citada
Akmal, A. y M.J.J. Janssens. 2004. Productivity and light use efficiency of perennial ryegrass with contrasting water and nitrogen supplies. Field Crops Res. 88, 143-155. [ Links ]
Angstrom, A. 1924. Solar and terrestrial radiation. Quart. J. Royal Meteorol. Soc. 50, 121-125. [ Links ]
Asohofrucol, 2007. Repollo. En: http://frutasyhortalizas.com.co; consulta: diciembre de 2007. [ Links ]
Corporación Colombia Internacional (CCI). 2006. Entorno nacional. pp. 41-44. En: Plan Hortícola Nacional. Corporación Colombia Internacional, Bogotá. 511 p [ Links ]
Curt, M.D., J. Fernández y M. Martínez. 1998. Productivity and radiation use efficiency of sweet sorghum (Sorghum bicolor L. Moench cv. Keller) in central Spain. Biomass Bioenergy 14, 169-178. [ Links ]
Dayan, E., H. Van Keulen, J.W. Jones, I. Zipori, D. Shmuel y H. Challa. 1993a. Development, calibration and validation of a greenhouse tomato growth model: I. Description of the model. Agric. Syst. 43, 145-163. [ Links ]
Dayan, E., H. Van Keulen, J.W. Jones, I. Zipori, D. Shmuel y H. Challa. 1993b. Development, calibration and validation of a greenhouse tomato growth model: II. Field calibration and validation. Agric. Syst. 43, 165-183. [ Links ]
DeJong, T.M. y Y.L. Grossman. 1994. A supply demand approach to modelling annual reproductive and vegetative growth of deciduous fruit trees. HortScience 29, 1435-1442. [ Links ]
Dewar, R.C., B.E. Medlyn y R.E. McMurtrie. 1998. A mechanistic analysis of light and carbon use efficiencies. Plant Cell Environ. 21, 573-588. [ Links ]
Dewar, R.C., 1996. The correlation between plant growth and intercepted radiation: an interpretation in terms of optimal nitrogen content. Ann. Bot. 78, 125-136. [ Links ]
De Wit, C.T. 1965. Photosynthesis of Leaf Canopies. PUDOC, Wageningen. [ Links ]
De Koning, A.N.M. 1994. Development and dry matter distribution in tomato: a quantitative approach. Dissertation. Wageningen Agric. Univ., Wageningen, 240 p. [ Links ]
France, J. y J.H.M. Thornley. 1984. Mathematical models in agriculture. A quantitative approach to problems in agriculture and related sciences. Butterworths, Londres. 335 p. [ Links ]
Francescangeli, N., M.A. Sangiacomo y H. Marti. 2006. Effects of plant density in broccoli on yield and radiation use efficiency. Scientia Hort. 110, 135-143. [ Links ]
Gallagher, J.N. y P.V. Biscoe. 1978. Radiation absorption, growth and yield of cereals. J. Agric. Sci. 91, 47-60. [ Links ]
Garcia, R., E.T. Kanemasu, B.L. Blad, A. Bauer, J.L. Hatfield, D.J. Major, R.J. Reginato y K.G. Hubbard. 1988. Interception and use efficiency of light in winter wheat under different nitrogen regimes. Agric. For. Meteorol. 44, 175-186. [ Links ]
Gifford, R.M. 1995. Whole plant respiration and photosynthesis of wheat under increased CO2 concentration and temperature: long-term vs. short-term distinctions for modelling. Glob. Change Biol. 1, 385-396. [ Links ]
Gijzen, H. 1995. Short term crop responses. En: Bakker, J.C., G.P.A. Bot, H. Challa y N.J. van de Braak (eds.) Greenhouse climate control, an integrated approach. Wageningen Pers, Wageningen. pp. 16-35. [ Links ]
Gosse, G., C. Varlet-Grancher, R. Bonhomme, M. Chartier, J. Allirand y G. Lemaire. 1986. Production maximale de matiére seche et rayonnement solaire intercepté par un couvert vegetal. Agronomie 6, 47-56. [ Links ]
Goudrian, J. y van Laar, H.H. 1994. Current sigues in production ecology. Modelling potential growth processes. Text book with exercises. Kluwer Academic Publishers, Londres. [ Links ]
Graf, B., O. Rakotobe, P. Zahner, V. Delucchi y A.P. Gutierrez. 1990. A simulation model for the dynamics of rice growth and development: Part I - The carbon balance. Agric. Syst. 32, 341-365. [ Links ]
Greenway, H. y R. Munns. 1980. Mechanisms of salt tolerance in non-halophytes, Annu. Rev. Plant Physiol. 31, 149-190. [ Links ]
Grossman, Y.L. y T.M. DeJong. 1994. Peach: a simulation model of reproductive and vegetative growth in peach trees. Tree Physiology 14(4), 329-345. [ Links ]
Hammer, G.L. y G.C. Wright. 1994. A theoretical analysis of nitrogen and radiation effects on radiation use efficiency in peanut. Aust. J. Agric. Sci. 45, 575-589. [ Links ]
Haxeltine, A. e I.C. Prentice. 1996. A general model for the light-use efficiency of primary production. Funct. Ecol. 10, 551-561. [ Links ]
Heuvelink, 1996. Dry matter partitioning in tomato: validation of a dynamic simulation model. Ann. Bot. 77, 71-80. [ Links ]
Imai, K., K. Shimabe, K. Tanaka y T. Kawana. 1994. Studies on matter production of edible canna (Canna edulis Ker.). Jpn. J. Crop Sci. 63, 345-51. [ Links ]
Jones, C.A. y J.R. Kiniry. 1986. CERES-Maize. A simulation model of maize growth and development. A& M University Press, Texas. [ Links ]
Jones, J.W. y J.T. Ritchie. 1990. Crop growth models. pp 63-69. En: Hofman, G.J., T.A. Howell y K.H. Solomon (eds.). Management of farm irrigation systems. ASAE, St. Joseph. [ Links ]
Kage, H., C. Alt y H. Stützel. 2001a. Predicting dry matter production of cauliflower (Brassica oleracea L. botrytis) under unstressed conditions. I. Photosynthetic parameters of cauliflower leaves and their implications for calculations of dry matter production. Scientia Hort. 87, 155-170. [ Links ]
Kage, H., H. Stützel y C. Alt. 2001b. Predicting dry matter production of cauliflower (Brassica oleracea L. botrytis) under unstressed conditions Part II. Comparison of light use efficiency and photosynthesis-respiration based modules. Scientia Hort. 87, 171-190. [ Links ]
Kooman, P. L. 1995. Yielding ability of potato crops as influenced by temperature and daylength. PhD. Thesis. Landbouw Universiteit Wageningen. [ Links ]
Kubota, F., Y. Matsuda, W. Ágata y K. Nada, 1994. The relationship between canopy structure and high productivity in napier grass, Pennisetum purpureum Schumach. Field Crops Res. 38, 105-10. [ Links ]
Lentz, W. 1998. Model applications in horticulture: a review. Scientia Hort. 74, 151-174. [ Links ]
Marcelis, L.F.M., E. Heuvelink y J. Goudriaan. 1998. Modelling biomass production and yield of horticultural crops: a review. Scientia Hort. 74, 83-111. [ Links ]
Marcelis, L.F.M. 1996. Sink strength as a determinant of dry matter partitioning in the whole plant. J. Exp. Bot. 47, 1281-1291. [ Links ]
Marcelis, L.F.M. 1994. A simulation model for dry matter partitioning in cucumber. Ann. Bot. 74, 43-52. [ Links ]
Marschner, H. 2002. Mineral Nutrition of Higher Plants. Academic Press, Londres. [ Links ]
Medlyn, B.E. 1998. Physiological basis of the light use efficiency model. Tree Physiol. 18, 167-176. [ Links ]
Monteith, J.L. 1969. Light interception and radiation exchange in crop stands. En: Easten, J.D. (ed.). Physiological aspect of crop yield. ASA, Madison, WI, pp. 89-111. [ Links ]
Monteith, J.L. 1977. Climate and the efficiency of crop production in Britain. Philos. Trans. R. Soc. London Ser. B 281, 277-294. [ Links ]
Monteith, J.L. 1996. The quest for balance in crop modeling. Agron. J. 88, 695-697. [ Links ]
Muchow, R.C. y R. Davies. 1988. Effect of nitrogen supply on the comparative productivity of maize and sorghum in a semi-arid tropical environment. Radiation interception and biomass accumulation. Field Crops Res. 18, 17-30. [ Links ]
Olesen, J.E. y K. Grevsen, 2000. A simulation model of climate effects on plant productivity and variability in cauliflower (Brassica oleracea L. botrytis). Scientia Hort. 83, 83-107. [ Links ]
Olesen, J.E. y K. Grevsen. 1997. Effects of temperature and irradiance on vegetative growth of cauliflower (Brassica oleracea L. botrytis) and broccoli (Brassica oleracea L. italica). J. Exp. Bot. 48, 1591-1598. [ Links ]
Penning de Vries, F.W.T., D.M. Jansen, H.F.M. Berger y A. Bakema. 1989. Simulation of ecophysiological processes of growth in several annual crops. Pudoc. Wageningen. 271 p. [ Links ]
Ross, J. 1981. The radiation regime and architecture of plant stands. Dr. Junk Publishers, The Hague. 391 pp. [ Links ]
Salazar, M.R., J.W. Jones, B. Chaves y A. Cooman. 2008. A model for the potential production and dry matter distribution of Cape gooseberry (Physalis peruviana L.). Scientia Hort. 115, 142-148. [ Links ]
Semillas Arroyave, 2006. Repollo Delus. En: http://www.semillasarroyave. com; consulta: diciembre de 2007. [ Links ]
Spitters, C.J.T., H. van Keulen y D.W.G. van Kraalingen. 1989. A simple and universal crop growth simulator: SUCROS87. pp. 107-122. En: Rabbinge, R., S.A. Ward y H.H. van Laar (eds.). Simulation and systems management in crop protection. Pudoc, Wageningen. [ Links ]
Steer, B.T., S.P. Milroy y R.M. Kamona. 1993. A model to simulate the development, growth and yield of irrigated sunflower. Field Crops Res. 32, 83-99. [ Links ]
Teh, C.B. 2004. Effect of leaf shape on the interception of solar radiation. En: Proc. 15th Malaysian Society of Plant Physiology Conference and the AGRI Congress. [ Links ]
Volkmar, K.M., Y. Hu y H. Steppuhn. 1998. Physiological responses of plants to salinity: a review. Can. J. Plant Sci. 78, 19-27. [ Links ]
Weir, A.H., P.L. Bragg, J.R. Porter y J.H. Rayner. 1984. A winter wheat crop simulation model without water or nutrient limits. J. Agric. Sci. 102, 371-382. [ Links ]
Wermelinger, B., J. Baumgartner y A.P. Gutierrez. 1991. A demographic model of assimilation and allocation of carbon and nitrogen in grapevines. Ecol. Model. 53, 1-26. [ Links ]
Wheeler, T.R., P. Hadley, R.H. Ellis y J.I.L. Morison. 1993. Changes in the growth and radiation use by lettuce crops in relation to temperature and ontogeny. Agric. Forest Meteorol. 66, 173-186. [ Links ]
Williams, J.R., C.A. Jones, J.R. Kiniry y D.A. Spanel. 1989. The EPIC crop growth model. Trans. Am. Soc. Agric. Eng. 32, 497-511. [ Links ]
Yunusa, I.A.M., K.H.M. Siddique, R.K. Belford y M.M. Karimi, 1993. Effect of canopy structure on efficiency of radiation interception and use in spring wheat cultivars during the preanthesis period in a mediterranean-type environment. Field Crops Res. 35, 113-22. [ Links ]

