










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.26 no.1 Bogotá Jan./June 2008
Gabriel Forero1, Martha Rodríguez2, Fernando Cantor3, Daniel Rodríguez3 y José Ricardo Cure3
1 Biólogo, Facultad de Ciencias, Universidad Militar Nueva Granada, Bogotá. gabrielforero@gmail.com
2 Jefe de Control Biológico Americaflor Ltda., Bogotá. lilianarodriguezc@yahoo.com
3 Docentes, Programa de Biología Aplicada, Facultad de Ciencias, Universidad Militar Nueva Granada, Bogotá. ecologia@umng.edu.co
Fecha de recepción: febrero 15 de 2008. Aceptado para publicación: abril 9 de 2008
RESUMEN
En cultivos de rosas en la Sabana de Bogotá, se ha registrado Amblyseius sp. como un ácaro depredador de Tetranychus urticae. Con el fin de evaluar el potencial de este ácaro en el manejo de T. urticae, se comparó en cultivos comerciales de rosa, bajo el sistema de agobio, la efectividad de liberaciones de Amblyseius sp. frente a la aplicación de productos de síntesis química, teniendo en cuenta los siguientes criterios de evaluación: presencia-ausencia del ácaro fitófago, porcentaje de daño causado en hojas y tipo de foco de T. urticae. Además, se evaluó la presencia del ácaro depredador ocho días después de las liberaciones. Se realizaron ensayos de respuesta funcional con densidades crecientes en los tres estados de desarrollo: huevo, larva o ninfas de la presa, bajo condiciones controladas y de invernadero. Se encontró menor cantidad (19,4%) de ácaros fitófagos con la aplicación de productos químicos. Sin embargo, el porcentaje de daño en la hoja fue menor (índice de daño 1 y 3, 8% y 13% menos, respectivamente) con las liberaciones de Amblyseius sp. Se evidenciaron diferencias en el tipo de foco para ambas estrategias de manejo y se registró una presencia semanal promedio del 23% para el ácaro depredador. La capacidad de consumo (respuesta funcional) a las 24 horas por parte de Amblyseius sp. fue de 6,66 huevos, 18,06 larvas y 19,15 ninfas bajo condiciones controladas, y de 4,56 huevos, 12,65 larvas y 15,71 ninfas bajo invernadero.
Palabras clave: ácaro depredador, ácaro fitófago, respuesta funcional, control biológico.
ABSTRACT
In rose cultures of the Bogota plateau, Amblyseius sp. as a mite predator of Tetranychus urticae was recorded. The purpose of this study was to evaluate the potential of this mite predator in the control of T. urticae. in commercial crops under the system of “agobio” comparing the effectiveness of releasing Amblyseius sp. versus the application of chemical products and taking into account the following evaluation criteria: the presence-absence of the phytophagous mite, percentage of damage to the leaves and T. urticae focus type. Also, the presence of the predaceous mite was evaluated eight days after the releases. Functional response tests were performed with increasing densities of three stages of development eggs, larvae and nymphs of the prey under controlled and greenhouse conditions. A minor quantity (19.4%) of phytophagous mites was found when chemical products were used, however the percentage of damage to the leaves was minor (damage index 1 and 3, 8% and 13% less, respectively) when Amblyseius sp. was released. The differences in the focus type were detected for both strategies of control and the weekly average presence was 23% for the predaceous mite. The capacity of consumption (functional response) after 24 h of Amblyseius sp. was 6.66 eggs, 18.06 larvae, and 19.15 nymphs under controlled conditions and 4.56 eggs, 12.65 larvae, and 15.71 nymphs under greenhouse conditions.
Key words: predatory mite, phytophagous mite, functional response, biological control
Introducción
El ácaro fitófago Tetranychus urticae Koch (Acari: Tetranychidae) es considerado actualmente como una de las plagas más importantes en cultivos bajo invernadero (Malais y Ravensberg, 1992), ya que estos alteran los procesos fisiológicos de las plantas como la fotosíntesis y la respiración, y afectan el crecimiento, la floración y la fructificación en las plantas que infestan (Hall y Ferree, 1975, citado por Monetti, 1999). Densidades entre 10 y 50 ácaros por hoja de rosa causan una reducción del largo del tallo de la flor del 17% y 26%, respectivamente, cuando se comparan con plantas que no tienen presencia de ácaros fitófagos (Landeros et al., 2004).
Aunque a nivel mundial se han recomendado varias estrategias para el control de T. urticae, el uso de enemigos naturales se ha aconsejado como estrategia de control biológico de esta plaga, en especial el uso del ácaro depredador Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae) (Scopes, 1985; Van Lenteren y Woets, 1988; Malais y Ravensberg, 1992; Decou, 1994; Zhang, 2003; Pedigo y Rice, 2006). Sin embargo, esta práctica ha sido menos común que la de los acaricidas químicos.
En Colombia existen algunos trabajos relacionados con ácaros depredadores, enfocados hacia el control biológico de ácaros fitófagos, entre ellos se destaca el de Mesa y Duque (1994), quienes realizaron liberaciones de tres especies de ácaros depredadores sobre cultivos de yuca y comprobaron que el proceso de preadaptación es fundamental para garantizar la sobrevivencia de los individuos en campo abierto. En rosas, se destacan los trabajos de França y De Vis (1995) y Barreto (2004), quienes utilizaron el ácaro depredador P. persimilis sobre la presa T. urticae y encontraron que el control biológico con este depredador es una alternativa que debe ser considerada y desarrollada con una continua investigación en la Sabana de Bogotá. Sin embargo, no hay trabajos que se refieran a los criterios de liberación de ácaros depredadores del género Amblyseius (Neoseiulus) en la fase comercial de la producción de rosas.
Para evaluar la capacidad de búsqueda y adquisición de presas, el cual es uno de los atributos que determinan la efectividad de un enemigo natural (Huffaker et al., 1976, citados por Cedola y Botto, 1996), hay que precisar parámetros específicos que se definen con estudios de respuesta funcional. Este tipo de estudios permiten identificar la máxima capacidad de adquisición del recurso que puede tener un depredador o parasitoide en función de la densidad en la que se presenta dicho recurso por unidad de tiempo. Cuando el número de presas consumidas/ parasitadas se encuentra en función del número de presas disponibles, se obtienen diferentes tipos de curvas o respuestas funcionales (Trexler et al., 1988).
Para el manejo de T. urticae en cultivos comerciales de rosas, se usan criterios basados en la implementación de estrategias de control químico. En la actualidad no se ha probado si esos criterios son aplicables para la implementación de estrategias de control biológico de T. urticae con liberaciones de controladores y, específicamente, con liberaciones de ácaros depredadores del género Amblyseius (Neoseiulus).
Con el fin de evaluar el potencial de Amblyseius sp. para el manejo de T. urticae los objetivos de este estudio fueron: evaluar el control de T. urticae mediante el uso del ácaro depredador Amblyseius sp., con base en criterios de liberación que se utilizan comúnmente en cultivos comerciales de rosas; determinar la respuesta funcional del ácaro depredador Amblyseius sp. sobre huevos, larvas y ninfas del ácaro fitófago T. urticae en condiciones controladas de laboratorio y de invernadero, como criterio para las liberaciones de Amblyseius sp. en campo.
Materiales y métodos
Obtención de Amblyseius sp.
Los individuos de Amblyseius sp. utilizados para los ensayos en áreas productivas y de respuesta funcional fueron obtenidos de una cría masiva del Laboratorio de Sanidad Vegetal (LSV) de Americaflor Ltda., los cuales provenían inicialmente de invernaderos comerciales de rosas en la Sabana de Bogotá.
Obtención de T. urticae
Los ensayos de respuesta funcional se realizaron con individuos T. urticae provenientes de cultivos comerciales de rosa con presencia de la plaga. Las muestras con ácaros fitófagos llegaban en cajas de cartón con hojarasca y hojas afectadas. Al interior de las cajas se colocaban plantas de fríjol Phaseolus vulgaris L. variedad bola roja de tres semanas de edad, hacia las cuales migraban los ácaros que se encontraban en las muestras recibidas, con lo cual suministraban plantas infestadas con la presa para los ensayos. Estas plantas se mantuvieron bajo condiciones de invernadero con una temperatura promedio de 16 ± 5,7 ºC y una humedad relativa promedio de 71±10,6%.
Experimentos en áreas productivas
El estudio se realizó en la finca comercial Florex C.I., dedicada al cultivo de rosas de exportación bajo invernadero, localizada en el municipio de Madrid (Cundinamarca), a una altura de 2.554 msnm y una temperatura promedio de 14,7 ºC. Las evaluaciones en campo se realizaron sobre plantas de rosa de la variedad Classy de dos años de edad, bajo el sistema de agobio. Estas plantas fueron sembradas en un área total de 6.720 m2, distribuidas en doce naves de invernadero. Cada nave estaba conformada por diez camas, a su vez compuestas por diez unidades experimentales (cuadros) cada una, y el área de la unidad experimental fue de 5,6 m2. Las camas se encontraban sembradas con aproximadamente 420 plantas, esto es, una densidad de 7,5 plantas/m2, las cuales recibieron los programas de fertirriego y de prácticas culturales específicas para el cultivo, de acuerdo con los criterios de manejo de la empresa.
El área total utilizada para la evaluación en campo fue dividida en dos partes iguales: 3.360 m2 fueron tratados con la rotación convencional de químicos que tenía la finca para todos los blancos biológicos (hongos y artrópodos plaga), los ingredientes activos de los productos de síntesis química usados hacia los ácaros fitófagos fueron: Difenilsoczasoline, Lactona macrociclica, Tiourea, Carbazato y Pyridazinona. Los restantes 3.360 m2 fueron tratados con toda la rotación de químicos, excepto acaricidas, ya que en esta área se realizaron las liberaciones del ácaro depredador. En el invernadero, la temperatura mínima promedio fue de 8±1,7 ºC y máxima de 34±3,3 ºC, con una humedad relativa mínima promedio de 33,9±10,8% y máxima de 88,3±14,1%. Estas condiciones fueron medidas con un DataLogger WatchDog®.
Definición y evaluación de tratamientos
En el tratamiento químico, se utilizó la rotación comercial de acaricidas de la finca. Los volúmenes en promedio fueron de ocho litros/cama bajo la dosis de cada producto utilizado. La frecuencia de aplicación fue semanal y para esta se utilizó una lanza japonesa con tres discos C-35 a los tres tercios, de abajo hacia arriba.
Para el tratamiento biológico, semanalmente, se liberaron en promedio 6,52 ácaros depredadores/m2 por cada cama (según recomendación de la empresa, basada en la experiencia adquirida en el uso del ácaro depredador). En los cuadros en los cuales se registró la presencia del ácaro fitófago, se liberaron 100 ácaros depredadores (18 individuos/ m2) y en los cuadros que no presentaban ácaros fitófagos, veinte ácaros depredadores (3,6 individuos/m2).
Para comparar los tratamientos biológico y químico, se realizaron dos muestreos directos semanales a lo largo de nueve semanas. El primer muestreo se realizó con el fin de identificar el grado de incidencia del ácaro fitófago (presencia- ausencia) en los tratamientos, donde se observaron todas las plantas del invernadero (cuadro por cuadro de todas las camas), para detectar la presencia de T. urticae; y el segundo, para evaluar el tipo de foco del ácaro fitófago, porcentaje de daño causado en hojas y presencia-ausencia del ácaro depredador. Se monitorearon al azar 40 cuadros del invernadero por cada tratamiento, donde se registraba la presencia de ácaros fitófagos con base en el monitoreo anterior. De cada uno de estos se tomaron al azar cinco hojas de la planta, sin importar el tercio, y se determinaba cada uno de los criterios a evaluar. Para clasificar el tipo de foco presente (tabla 1), se modificó una escala propuesta por Acosta (2000) y, para el grado de severidad en la hoja, se utilizó una escala de severidad realizada con anterioridad para la elaboración del trabajo (tabla 2). La evaluación de presencia-ausencia del depredador solamente se realizó en el tratamiento biológico, puesto que las aplicaciones de acaricidas en el tratamiento químico imposibilitaban el establecimiento del depredador al no ser compatible con los químicos utilizados.


Para comparar la incidencia del ácaro fitófago, se realizó un análisis de varianza (Anova) y para establecer las diferencias significativas en el tipo de foco y porcentaje de daño en la hoja presente en cada uno de los tratamientos, se realizaron pruebas de comparación de proporciones. La evaluación de presencia-ausencia del ácaro depredador se obtuvo con el promedio semanal de cuadros donde en este se encontraba presente Amblyseius. Los criterios evaluados se analizaron estadísticamente con el software R (2004, versión 2.0.1).
Determinación de la respuesta funcional de Amblyseius sp.
Se utilizaron hembras adultas de dos días de emergencia. Antes de los ensayos, se dejaron los ácaros depredadores en condiciones de ayuno por un periodo de 4 h, para garantizar el consumo de los ácaros fitófagos. La capacidad depredadora de Amblyseius sp. se evaluó variando la densidad de huevos (5, 10, 20, 40, 80 y 160), larvas (3, 6, 12, 24 y 48) y ninfas (3, 6, 12, 24 y 48) de T. urticae, el tiempo de evaluación del consumo de presas fue de 24 h.
Se realizó un diseño experimental de bloques completamente al azar. Cada bloque experimental consistió en una caja plástica de 10 cm de largo, 4 cm de ancho y 8 cm de alto. Dentro de las cajas se hizo una adaptación de la técnica del disco flotante propuesta por Boykin y Campbell (1982), para realizar el montaje con presas y ácaros depredadores. En el fondo de cada caja se fijó una lámina de cartón, en donde se incrustaron alfileres que atravesaban el centro de cinco a seis discos de hoja (unidades experimentales) dependiendo del número de densidades de presa a evaluar. Los discos obtenidos de foliolos apicales de rosa variedad Charlotte tenían un diámetro de 2,5 cm, en los cuales, con el envés hacia arriba, se colocaron las diferentes densidades de presas (tratamientos) con su respectivo adulto depredador. Al interior de la caja con el montaje se adicionaron 190 mL de agua, con el fin de evitar que los estados móviles de los ácaros se desplazaran a través de los alfileres y de la lámina de cartón. De esta manera, los discos quedaban flotando dentro de la caja. Además, para evitar que los ácaros se desplazaran entre los discos por la tensión superficial del agua, se adicionaron 10 mL de una mezcla de detergente convencional con agua (0,5 g en 200 mL de agua).
Se emplearon seis bloques experimentales para cada instar de la presa (huevos, larvas y ninfas) por ambiente de evaluación (invernadero y laboratorio), se utilizaron de cinco a seis unidades experimentales (discos de hoja) para cada uno de los tratamientos (densidades evaluadas) y se realizaron seis repeticiones por tratamiento en cada uno de los ambientes. En total, fueron utilizadas 72 unidades experimentales para el ensayo del instar de huevos y un total de 60 en cada uno de los estados restantes (larvas y ninfas).
Los experimentos realizados bajo condiciones de laboratorio se mantuvieron bajo control de temperatura (23±1,5 ºC), humedad (65±3,7%) y luminosidad 12:12 (L:O). En invernadero los montajes de las unidades experimentales se hicieron dentro de una estructura de 1,20 m x 1,20 m cubierta con malla de polisombra de color blanco. En este último caso, la temperatura mínima promedio fue de 7±1,9 ºC y la máxima fue de 32±3,1 ºC, con una humedad relativa mínima promedio de 39,1±9,6% y máxima de 88,5±12,1%. Estas condiciones fueron medidas con un datalogger Temptale 4®.
La respuesta funcional de Amblyseius sp. fue ajustada mediante el siguiente modelo propuesto por Gutiérrez (1996):
Donde: Na es el número de individuos atacados; -a es la proporción de presas disponibles para el ataque; D es la demanda per cápita de presas por unidad de tiempo; P es el número de depredadores; N es el número de presas disponibles y 1-e es la proporción de presas en refugio.
La estimación de los parámetros se realizó con el procedimiento PROC NLIN que realiza el ajuste a modelos mediante regresión no lineal y usando el algoritmo Dud (SAS Institute, 2002). Para graficar las curvas de respuesta funcional, los valores de alpha (a) y demanda (D) obtenidos en el procedimiento PROC NLIN fueron ingresados al modelo, posteriormente se reemplazaron las variables en una hoja de cálculo y se realizaron las gráficas de las curvas para un total de 100 presas para los instares de huevos, larvas y ninfas.
Resultados y discusión
Evaluación de las diferentes estrategias de manejo para T. urticae
Grado de incidencia del ácaro fitófago. El número promedio de cuadros afectados por ácaros fitófagos durante las nueve semanas de monitoreo cuando se realizó la estrategia de liberación de controladores biológicos fue de 87,55 cuadros, mientras que, cuando se realizó la estrategia de aplicación de químicos fue de 70,55 cuadros (24,3% y 19,6%, respectivamente). Al analizar estos resultados, se encontró que presentan una diferencia significativa a favor del tratamiento con la estrategia de aplicaciones de acaricidas químicos (F = 5,36; gl = 1; P = 0,034).Es evidente que la eficacia del control biológico con liberaciones de Amblyseius sp. es menor con relación a la del control químico con acaricidas para reducir poblaciones de T. urticae en cultivos de rosa; cuando la decisión del control se basa en análisis de criterios como el número de cuadros afectados con la plaga. Este tipo de análisis es común entre los floricultores como criterio para evaluar la efectividad de una estrategia de control químico. Sin embargo, el mismo criterio ya no es funcional para evaluar la efectividad de liberaciones de ácaros depredadores, porque no considera el tamaño real de las poblaciones del ácaro fitófago. Esta modificación es necesaria para estimar el número de ácaros depredadores que se debe liberar para el control de poblaciones reales de T. urticae. Por lo anterior, es probable que en el presente trabajo el efecto del control biológico hubiera sido menor al del control químico, ya que el número de ácaros depredadores pudo ser insuficiente para controlar las poblaciones de T. urticae presentes en el cultivo. Por esta razón, el concepto de respuesta funcional evaluado en este trabajo puede ser una herramienta para complementar la efectividad de Amblyseius sp. sobre poblaciones de T. urticae.
Tipo de foco del ácaro fitófago. En la tabla 3 se evidencian diferencias únicamente en los focos tipo 0, 1 y 2 como resultado de las estrategias de control químico y biológico. Los focos tipo 0 que fueron el resultado de la estrategia de control químico corresponden al 7% de los focos tratados, mientras que los que fueron el resultado de la estrategia de control biológico apenas corresponden al 3%. Además, los focos tipo 1 se obtienen como resultado de la efectividad del control químico en el 62% de los casos, mientras que este tipo de focos se logra como resultado de la efectividad del control biológico en apenas el 41% de los casos. Sin embargo, los focos tipo 2 se logran en aproximadamente el 44% de los casos cuando se realiza una estrategia de control biológico y en apenas el 20% de los casos cuando la estrategia es el control químico.
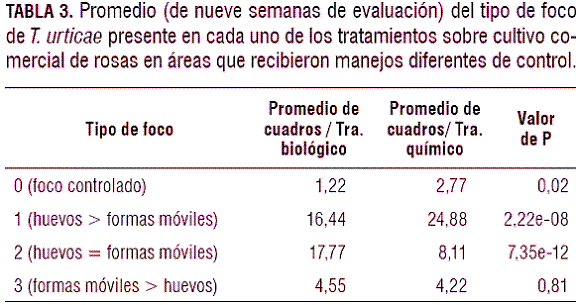
En condiciones comerciales se espera que después de cualquier estrategia de manejo de T. urticae se concentre la mayor cantidad de focos tratados en focos tipo 0 y la menor cantidad de focos en tipo 3. Aunque el control químico permitió la obtención de la mayor cantidad de focos tipo 0- 7%, el mayor porcentaje de focos controlados se concentra en los focos tipo 1 - 62%. En contraste con la situación anterior, la estrategia de control biológico no generó un alto porcentaje de focos tipo 0, sino que estos valores se distribuyen de manera aproximadamente equitativa entre focos tipo 1 - 41% y tipo 2 - 44%.
El bajo índice de control logrado por parte del ácaro depredador Amblyseius sp. pudo verse afectado probablemente por el tamaño de la población inicial del fitófago, porque al momento de la liberación de depredadores se presentaban más huevos del fitófago y pocos estados móviles. Esto podría soportarse por los resultados de Walzer y Schausberger (1999) y Schausberger y Walzer (2001), quienes afirman que ácaros depredadores de la especie Amblyseius californicus McGregor presentan preferencia por larvas y ninfas de T. urticae y no por huevos del fitófago. Además, los bajos índices de control de Amblyseius sp. también podrían explicarse, porque probablemente la cantidad de depredadores liberados no fue suficiente para ejercer un buen control.
Grado de severidad en la hoja. Los resultados de la tabla 4 enseñan diferencias únicamente en los índices de daño 1 y 3 luego de utilizadas las dos estrategias de control (biológico y químico). El resultado para el índice de daño 1 cuando se utiliza la estrategia de control biológico corresponde al 44% de los cuadros tratados. Sin embargo, al utilizarse la estrategia de control químico se obtiene solamente un 36% de cuadros que presentan este índice de daño. Por otra parte, el índice de daño 3 se logra en el 9% de los casos como resultado de la estrategia del control biológico, mientras que, al utilizarse la estrategia de control químico, el resultado es de únicamente el 22% de los casos.
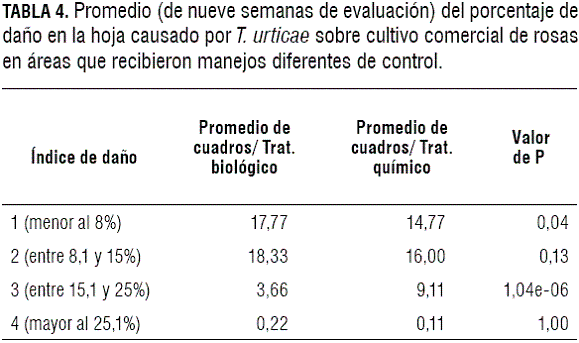
En cultivos comerciales de rosas, se desea obtener la menor cantidad de índices de daño alto luego de realizar cualquier tipo de estrategia para el manejo de T. urticae, por lo tanto, se esperaría que la mayoría de cuadros evaluados se ubicaran en los índices de daño 1 y la menor cantidad en los índices de daño 3 y 4. Cuando se realiza la estrategia de control químico se obtienen valores agrupados en los índices 1, 2 y 3, con 36%, 40% y 22%, respectivamente. En comparación, la estrategia de control biológico presentó valores agrupados de manera similar en los índices 1 (44%) y 2 (45%).
Las implicaciones prácticas que puede tener la presencia de menores daños causados sobre las hojas al utilizar como controlador el ácaro depredador Amblyseius sp. puede ser una ventaja para el cultivo en parámetros de productividad representada en una mayor área fotosintética.
Monitoreo de presencia-ausencia del depredador. En la tabla 5 se exponen los resultados para determinar la presencia del depredador en el tratamiento biológico. Estos resultados corresponden al número de cuadros registrados durante las nueve semanas de evaluación que se realizaron ocho días después de implementar la estrategia de manejo biológico. Para el análisis de este parámetro se obtuvo un promedio semanal de 9,33 cuadros, que equivalen al 23% de presencia a lo largo de las nueve semanas. Además, en el presente trabajo también se demuestra que las liberaciones periódicas de Amblyseius sp. permiten que el depredador pueda establecerse en el cultivo, lo que podría implicar reducción de costos de control por disminución de liberaciones.

Respuesta funcional de Amblyseius sp. sobre huevos, larvas y ninfas de T. urticae
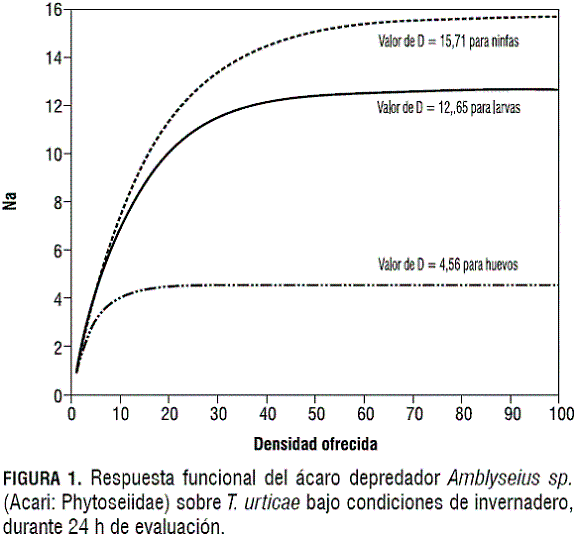
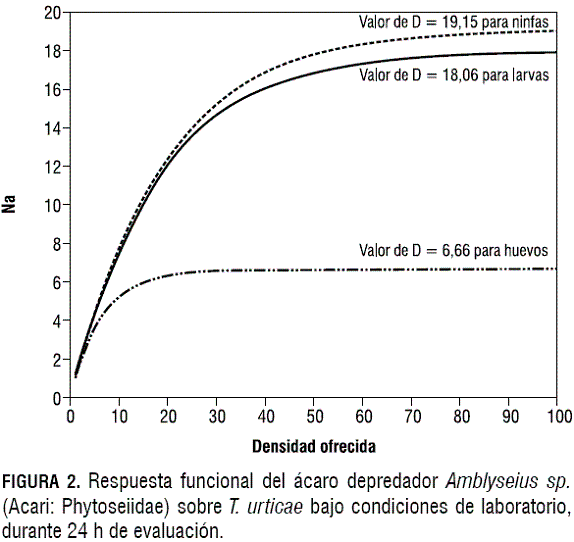
Los modelos ajustados que describen el consumo de huevos, larvas y ninfas de T. urticae por parte de Amblyseius sp., bajo condiciones de invernadero y de laboratorio a las 24 h de evaluación, se presentan en las figuras 1 y 2, respectivamente. Los parámetros de ajuste y los valores de significancia de los modelos ajustados se presentan en la tabla 6.

Se observa una respuesta funcional tipo II (Holling, 1959, citado por Huffaker y Gutiérrez, 1999) para Amblyseius sp., la cual se caracteriza porque la tasa de consumo aumenta conforme aumenta la densidad de la presa, pero gradualmente va desacelerando hasta que llega a un punto donde esta tasa es constante, independientemente de la densidad de la presa.
En las figuras 1 y 2 se presentan las curvas de respuesta funcional de Amblyseius sp. cuando se le ofrecen huevos, larvas y ninfas de T. urticae bajo condiciones de laboratorio y de invernadero. Es evidente que, en un periodo de evaluación de 24 h, Amblyseius sp. es capaz de consumir más larvas (D= 18,06) y ninfas (D= 19,15) que huevos (D= 6,66) bajo condiciones de laboratorio. Una tendencia similar se observa cuando se analizan los consumos de los diferentes estados de T. urticae por Amblyseius sp. bajo condiciones de invernadero, en un periodo igual de 24 h. En esta situación Amblyseius sp. consume más larvas (D= 12,65) y ninfas (D= 15,71) que huevos (D= 4,56). Walzer et al. (1999) reportan a Amblyseius californicus como un depredador generalista, sin embargo, este autor menciona la preferencia del depredador hacia los estados de larvas y ninfas. A su vez, Schausberger et al. (2001) afirman la preferencia de este depredador por estados inmaduros móviles de la presa. Basándose en esta información es posible inferir que el mayor consumo de larvas y ninfas presentado por Amblyseius sp. sea por el hábito de preferencia hacia inmaduros móviles del ácaro fitófago.Estudios de respuesta funcional realizados para Amblyseius californicus en otras especies vegetales informan la eficiencia en el consumo de los diferentes estados de T. urticae. Cedola et al. (2001) reportan un consumo de 4 huevos en 24 h para una densidad ofrecida de 64 presas en tomate (Lycopersicum esculentum) por parte de deuteroninfas del depredador. Castagnoli y Simoni (1999) reportan un consumo de 17 huevos en 24 h a una densidad ofrecida de 80 presas, y un consumo de 9,9 ninfas en 24 h a una densidad ofrecida de 30 presas en fresa (Fragaria ananassa) por parte de hembras adultas del depredador. La baja eficiencia en el consumo por parte de Amblyseius californicus en tomate se debe a que esta especie presenta una gran cantidad de tricomas, afectando la movilidad del depredador sobre la hojas (Cedola et al., 2001). Esta misma autora reporta que la fresa presenta tricomas pero no glandulares, los cuales afectan la respuesta funcional del depredador.
En rosa (Rosa sp.), al no presentarse ningún tipo de tricomas en las hojas (Sabelis, 1981, citado por Krips et al., 1999), es probable que la respuesta funcional del depredador se vea favorecida, puesto que no existe la barrera física que limita la capacidad de búsqueda. Skirvin y Fenlon (2001) encontraron que la morfología de la hoja afecta las tasas de consumo del ácaro depredador P. persimilis, y observaron que en hojas glabras hay una mayor eficiencia en el consumo. De igual forma, Workman y Martín (2000) informan que P. persimilis presenta una mayor movilidad en hojas de Rosa sp. comparada con otras flores de corte. Amblyseius sp., al pertenecer a la misma familia de P. persimilis, probablemente exhibe el mismo comportamiento en hojas que carecen de tricomas, por lo que los valores de D encontrados sobre plantas de rosa fueron mayores en comparación con otras especies vegetales.
Al comparar el consumo por parte de hembras adultas utilizadas en este trabajo, en relación con estudios anteriores realizados por Cedola et al. (2001), donde se evaluó el consumo por parte del depredador Neoseiulus californicus en estados de protoninfas y deuteroninfas, se observa que estos estados inmaduros tienen un menor nivel de consumo en comparación con las hembras adultas. Según Shipp y Whitfield (1991), las hembras consumen mayor cantidad de alimento, debido a que poseen un gasto de energía extra relacionado con el desarrollo de huevos, lo que probablemente influye en el mayor consumo de presas.
Los análisis de respuesta funcional realizados en el presente trabajo son importantes, ya que establecen la máxima relación constante de liberación de poblaciones de Amblyseius sp. sobre poblaciones de edad específica de T. urticae, como edades susceptibles de ser consumidas en determinada unidad de tiempo. De acuerdo al presente trabajo, se puede asumir como uno de los criterios de liberación, un individuo de Amblyseius sp. por cada 20 larvas o ninfas de T. urticae para que sean consumidas en un periodo de 24 h. Sin embargo, para que dicha proporción sea efectiva, es necesario realizar monitoreos de la plaga que permitan no solo estimar su población en el campo, sino también determinar su composición de edades, como criterios que permitan la posibilidad de realizar liberaciones de enemigos naturales como el ácaro depredador Amblyseius sp.
Conclusiones
• Los criterios tradicionales para evaluar la presencia de T. urticae en campo permiten identificar la eficacia de estrategias de control químico de la plaga y no de estrategias de control biológico. Cuando se considera la posibilidad de evaluar la eficacia de liberaciones de Amblyseius sp., es necesario incorporar nuevos criterios. Evaluar liberaciones en campo de ácaros depredadores para el control de T. urticae con los criterios tradicionales no permitirá identificar el verdadero potencial de estos enemigos naturales.
• No se lograron altos índices de control de T. urticae con liberaciones de Amblyseius sp. bajo condiciones de campo, por la utilización de criterios establecidos por los cultivos comerciales de rosas. Es necesario tener en cuenta en la liberación del ácaro depredador un criterio que permita relacionar la máxima capacidad de consumo del depredador por unidad de tiempo con la cantidad de presas presentes en un cultivo.
• Si se aplica el concepto de respuesta funcional evaluado en este trabajo como nuevo criterio para aplicar en campo, se requiere de la estimación del tamaño de las poblaciones de la plaga en el cultivo y de la composición de las edades de la misma.
Agradecimientos
A la Universidad Militar Nueva Granada y al Laboratorio de Control Biológico de Americaflor Ltda. por el apoyo financiero y logístico del presente proyecto.
Literatura citada
Acosta, A. 2000. Escrito sobre lo que seguramente usted ya ha oído de los famosos ácaros Tetranychus spp. Conocimientos generales y manejo integrado de Tetranychus spp. (Acariformes: Tetranychidae) en cultivos de flores. Facultad de Agronomía, Universidad Nacional de Colombia, Novartis. 47 p. [ Links ]
Badii, M.H., E. Hernández, A.E. Flores y J. Landeros. 2004. Prey stage preference and functional response of Euseius hibisci to Tetranychus urticae (Acari: Phytoseiidae. Tetranychidae). Expl. Appl. Acarology 34, 263-273. [ Links ]
Barreto, C. 2004. Introducción del depredador Phytoseiulus Persimilis en rosas. Trabajo de grado. Facultad de Agronomia, Universidad Nacional de Colombia, Bogotá. [ Links ]
Boykin, L. y W. Campbell. 1982. Rate of population increase of the twospotted spider mite (Acari: Tetranychidae) on peanut leaves treated with pesticides. J. Econ. Entomol. 75, 966-971. [ Links ]
Castagnoli, M.y S. Simoni. 1999. Effect of long-term feeding history on functional and numerical response of Neoseiulus californicus (Acari: Phytoseiidae). Expl. Appl. Acarology 33, 217-234. [ Links ]
Cedola, C.V., N.L. Sanchez y G. Liljesthrom. 2001. Effect of tomato leaf hairiness on functional and numerical response of Neoseulus californicus (Acari: Phytoseiidae). Expl. Appl. Acarology 25, 819-831. [ Links ]
Cedola, C.V. y E.N. Botto. 1996. Evaluación de la respuesta funcional de Amblyseius idaeus Moraes y Mac Murtry. 1983 y Phytoseiulus macropilis (Banks. 1905) (Acarina: Phytoseiidae) en condiciones de laboratorio. Rev. Chilena Entomol. 23, 15-18. [ Links ]
Chen, H.C. 2000. Spider-mite problems and control in Taiwan. Expl. Appl. Acarology 24, 453-462. [ Links ]
Decou, G. 1994. Biological control of the two-spotted spider mite (Acarina: Tetranychidae) on comercial strawberries in Florida with Phytoseiulus persimilis (Acarina: Phytoseiidae). Florida Entomologist 77 (1), 33-41. [ Links ]
Easterbrook, M.A., J.D. Fitzgerald y M.G. Solomon. 2001. Biological control of strawberry tarsonemid mite Phytonemus pallidus and two-spotted spider mite Tetranychus urticae on strawberry in the UK using species of Neoseiulus (Amblyseius) (Acari: Phytoseiidae). Expl. Appl. Acarology 25, 25-36. [ Links ]
Franca, S. y R. De Vis. 1995. Control biológico de Tetranychus urticae Koch (Acari: tetranychidae) con Phytoseiulus persimilis Athias-henriot (Acari: Phytoseiidae) en rosa en la Sabana de Bogotá. Rev. Colomb. Entomol.22 (3), 153-157. [ Links ]
Getiva, J. y A. Acosta. 2004. Taxonomía de ácaros asociados a cultivos de flores. Asocolflores 66, 59-68. [ Links ]
Goka, K. 1999. The effect of patch size and persistence of host plants on the development of acaricide resistance in the two-spotted spider mite Tetranychus urticae (Acari: Tetranychidae). Expl. Appl. Acarology 23, 419-427. [ Links ]
Gough, N. 1991. Long term stability in the interactions between Tetranychus urticae and Phytoseiulus persimilis producing succesful integrated control on roses in southeast Queensland. Expl. Appl. Acarology 12, 83 - 101. [ Links ]
Gutiérrez, A.P. 1996. Applied population ecology: a supply-demand approach. John Wiley and Sons, Nueva York. 300 p. [ Links ]
Herron, G.A. y J. Rophail. 1998. Tebufenpyrad (Pyranica®) resistance detected in two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) from apples in Western Australia. Expl. Appl. Acarology 22, 633-641. [ Links ]
Herron, G.A. y J. Rophail. 2002. The stability of tebufenpyrad resistance in two-spotted spider mite (Acari: Tetranychidae) under laboratory conditions. Expl. Appl. Acarology. 26, 253-256. [ Links ]
Herron, G.A. y J. Rophail. 2003. First detection of chlorfenapyr (Secure®) resistance in two-spotted spider mite (Acari: Tetranychidae) from nectarines in an Australian orchard. Expl. Appl. Acarology 31, 131-134. [ Links ]
Herron, G.A., J. Rophail, y L.J. Wilson. 2004. Chlorfenapyr resistance in two-spotted spider mite (Acari: Tetranychidae) from Australian cotton. Expl. Appl. Acarology 34, 315-321. [ Links ]
Huffaker, C.B. y A.P. Gutierrez. 1999. Ecological entomology. 2nd edition. John Wiley and Sons, Nueva York. 756 p. [ Links ]
Krips, O.E., P.W. Kleign, P.E. L. Willems, G.J.Z. Gols y M. Dicke. 1999. Leaf hairs influence searching efficiency and predation rate of the predatory mite Phytoseiulus persimilis (Acari: Phytoseiidae). Expl. Appl. Acarology 23 (2), 119-131. [ Links ]
Landeros, J., L.P. Guevara, M.H. Badii, F. Flores y A. Pámanes. 2004. Effect of different densities of the twospotted spider mite Tetranychus urticae on CO2 assimilation. transpiration and stomatal behaviour in rose leaves. Expl. Appl. Acarology 32, 187-198. [ Links ]
Malais, M. y W.J. Ravensberg. 1992. The biology of glasshouse pests an their natural enemies. Knowing and recognizing. Koppert Biological Systems. 117 p. [ Links ]
Martínez, V., F. Sáenz-de-Cabezón, F. Moreno-Grijalba, V. Marco y Perez-Moreno. 2005. Effects of azadirachtin on the two-spotted spider mite. Tetranychus urticae (Acari: Tetranychidae). Expl. Appl. Acarology 35, 215-222. [ Links ]
Mendoza, L.A., A. Acosta, J. Ospina y L. Díaz. 2001. Evaluación de diferentes estados de ácaros del género Tetranychus en cultivos de flores de la Sabana de Bogotá. Asocolflores 60, 11-24. [ Links ]
Mesa, N. y A. Belloti. 1986. Ciclo de vida y hábitos alimenticios de Neoseiulus anonymus. Predador de ácaros Tetranychidae en yuca. Rev. Colomb. Entomol. 12 (1), 54-66. [ Links ]
Mesa, N., A. Belloti y M. Duque. 1988. Ciclo de vida y tasa de incremento natural de Galendromus annectens. Neoseiulus idaeus y Phytoseiulus persimilis (Acari: Phytoseiidae). Rev. Colomb. Entomol. 14 (2), 41-49. [ Links ]
Mesa, N. y M. Duque. 1994. Liberación y establecimiento de tres especies de ácaros Phytoseiidae para el control de ácaros Tetranychidae en un cultivo de yuca. Rev. Colomb. Entomol. 20(3), 169-177. [ Links ]
Monetti, L. 1999. Estudio de los atributos vitales de los ácaros fitoseidos y su aplicación al control biológico de plagas. Rev. Soc. Entomol. Argentina 58 (1-2), 48-57. [ Links ]
Nicetic, O. D. M. Watson, G. A. C. Beattie, A. Meats y J. Zheng. 2001. Integrated pest management of two-spotted mite Tetranychus urticae on greenhouses roses using petroleum spray oil and the predatory mite Phytoseiulus persimilis. Expl. Appl. Acarology 25, 37-53. [ Links ]
Niemeczyk, E. 1998. Effectiveness of predatory mites (Phytoseiidae) in limiting population of two-spotted spider mites (Tetranychus urticae Koch) on Black currants. Akademia Techniczno- Rolnicza I.M. Jana I. Jedrzeja sniadeckich Wbydgoszczy zeszyty Naukowe 214- Ochrona Srodowiska 2, 65-74. [ Links ]
Pedigo, L. y M. Rice. 2006. Entomology and pest management. Pearson Prentice Hall, Nueva York. 749 p. [ Links ]
R. Copyright. 2004. The R. Foundation for Statistical Computing Versión 2.0.1 (2004-11-15). ISBN 3-900051-07-0. [ Links ]
Ríos, L., M. Jiménez y L. Smith, L. 1999. Estudios básicos a la optimización de un sistema de cría masiva de Typhlodromalus tenuiscutus (Acari: Phytoseiidae) un depredador de ácaros de yuca. Rev. Colomb. Entomol. 25 (1-2), 83-90. [ Links ]
Salazar, F., R. Rebolledo, R. Carrillo y P. Aguilera. 2000. Factores ambientales y de la planta relacionados con la diapausa de hembras de Tetranychus urticae Koch (Acari: Tetranychidae) en Temuco, Chile. Rev. Chilena Entomol. 27, 53-56. [ Links ]
Santamaría, G., C. Salas, A. Acosta y J. Amador. 2002. Caracterización de la ubicación de focos de ácaros fitófagos dentro de invernaderos para cultivos de rosa, clavel y alstroemeria. Asocolflores 61, 44-60. [ Links ]
SAS Institute 2002. SAS V.8.0. SAS Institute, Cary, N.C. [ Links ]
Schausberger, P. y A. Walzer, A. 2001. Combined versus single species release of predaceous mites predator-predator interactions and pest suppression. Biol. Control 20, 269-278. [ Links ]
Scopes, N.E.A. 1985. Red spider mite and the predator Phytoseiulus persimilis pp. 43-52. En: Biological pest control - The glasshouse experience. Hussey, N.W. y N. Scopes (eds.). Cornell University Press. 240 p. [ Links ]
Shipp, J.L. y G.H. Whitfield. 1991. Functional response of the predatory mite. Amblyseius cucumeris (Acari: Phytoseidae) on wester flower thrips. Frankliniella occidentalis (Thysanoptera: Thripidae). Environ. Entomol. 20(2), 694-699. [ Links ]
Skirvin, D.J. y J.S. Fenlon. 2001. Plant species modifies the functional response of Phytoseiulus persimilis (Acari: Phytoseiidae) to Tetranychus urticae (Acari: Tetranychidae): implications for biological control. Bull. Entomol. Res. 91, 61-67. [ Links ]
Trexler, J.C., C.E. Mcculloch y J. Travis. 1988. How can the functional response best be determined?. Oecologia 76, 206-214. [ Links ]
Van Lenteren, J.C. y J. Woets. 1988. Biological and integrated pest control in greenhouses. Annu. Rev. Entomol. 33, 239-269. [ Links ]
Walzer, A. y P. Schausberger. 1999. Cannibalism and interspecific predation in the phytoseid mites Phytoseiulus persimilis and Neoseiulus californicus: predation rates and effects on reproduction and juvenile development. Biocontrol 43, 457- 468. [ Links ]
Wilson, L.T., P.J. Trichilo y D. Gonzalez. 1991. Natural enemies of spider mites (Acari: Tetranychidae) on cotton: Density Regulation or Casual Association?. Environ. Entomol. 20 (3), 849-856. [ Links ]
Workman, P. y N. Martin. 2000. Movement of Phytoseiulus persimilis (Acari: Phytoseidae) on leaves of greenhouse carnations an other cut flowers. New Zealand J. Crop Hort. Sci. 28, 9-15. [ Links ]
Zhang, Z. 2003. Mites of greenhouses: identification. biology and control. Cabi Publishing. 256 p. [ Links ]

