










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.26 no.3 Bogotá Sep./Dec. 2008
Fabio Ernesto Martínez1, Jenny Sarmiento2, Gerhard Fischer3 y Francisco Jiménez4
1 Ingeniero agrónomo, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. femartinezma@unal.edu.co
2 Ingeniera agrónoma, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. jsarmientoc@unal.edu.co
3 Profesor asociado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. gfischer@unal.edu.co
4 Jefe de asistencia técnica, Monómeros Colombo-Venezolanos, Bogotá (Colombia). frjimenez@monomeros.com.co
Fecha de recepción: 3 de junio de 2008. Aceptado para publicación: 5 de noviembre de 2008
RESUMEN
A pesar de ser la uchuva el segundo fruto de exportación más importante de Colombia, existe poca información sobre el efecto de una nutrición deficiente en la producción y calidad de sus frutos. Plantas de uchuva, propagadas por semilla, se sembraron en sustrato arena cuarcítica en materas de plástico (25 L) y se mantuvieron durante 135 días en condiciones de invernadero de plástico en Bogotá. Las deficiencias nutricionales se indujeron a través de soluciones nutritivas, usando el método del elemento faltante y empleando un diseño completamente al azar con 8 tratamientos y 5 repeticiones: 1. testigo con fertilización completa; 2. fertilización completa -N; 3. -P; 4. -K; 5. -Ca; 6. -Mg; 7. -B y 8. solo agua potable. En producción se evaluó el diámetro, peso y número de los frutos, el rendimiento y el peso seco de frutos y cáliz. Para la calidad del fruto se consideraron las variables sólidos solubles totales (SST), acidez total titulable (ATT) y el pH. El tratamiento -B afectó negativamente las variables de producción como tamaño, peso fresco y seco del fruto y masa seca del cáliz. Las plantas deficientes en N y K redujeron el peso fresco y seco de sus frutos considerablemente. Comparados con el testigo, todos los otros tratamientos disminuyeron el número de frutos y el rendimiento por planta, los -B, -N y -K disminuyeron en más del 90% la cantidad de frutos producidos y el rendimiento, y el tratamiento -P en un 50%. El contenido de SST en frutos se redujo en plantas deficientes de B, mientras la ATT se incrementó en aquellas con deficiencia de P, comparada con la de -Ca.
Palabras clave: deficiencias nutricionales, peso fruto, sólidos solubles, acidez titulable, pH.
ABSTRACT
The cape gooseberry is the second largest export fruit from Colombia but little information exists about how deficient nutrition affects production and quality of fruits. Seed propagated cape gooseberry plants were kept in 25 L plastic containers filled with quartz sand, under plastic greenhouse conditions, during 135 days in the city of Bogotá. Through nutrient solutions, nutrient deficiencies were applied using the missing element technique in a completely randomized design with 8 treatments and 5 replicates: 1. the control treatment with complete fertilization; 2. complete fertilization -N; 3. -P; 4. -K; 5. -Ca; 6. -Mg; 7. -B, and 8. only tap water. The production variables such as diameter, fresh and dry weight, number of fruits, yield, and calyx dry mass were evaluated. For fruit quality the variables total soluble solids (TSS), total titratable acids (TTA), and pH were evaluated. The treatment without B negatively affected the production variables: diameter, fresh and dry weight of fruit and calyx dry mass. Plants with N and K deficiencies had considerably reduced fresh and dry weight of fruits. In comparison with the control all other treatments had diminished fruit quantity and yield per plant. The treatments deficient in B, N and K decreased the amount of fruits and yield by 90%, and in the treatment -P fruit and yield decreased by more 50%. Fruit TSS content was reduced in plants with B deficiency, whereas TTA concentration increased in plants with P deficiencies, compared with those of -Ca.
Key words: nutrient deficiencies, fruit weight, soluble solids, titratable acids, pH.
Introducción
La uchuva (Physalis peruviana L.) es una solanácea originaria de Perú, de hábito silvestre o semisilvestre que crece en Suramérica en zonas entre 1.500 y 3.000 msnm. En Colombia se cultiva en sitios entre 1.800 y 2.800 msnm, con temperaturas promedio de 13 a 18 °C y precipitaciones de 1.000 a 1.800 mm/año (Fischer, 2000). En el país la uchuva es cultivada por pequeños productores con bajo nivel de asistencia técnica; esto ha generado diversas prácticas basadas en la experiencia de cada agricultor, lo que a su vez implica un alto consumo de insumos, ausencia de tecnología y elevado uso de mano de obra (Sanabria, 2005).Los efectos nutricionales dependen de la influencia que ejerce cada nutriente en particular sobre los procesos bioquímicos y fisiológicos de la planta (Mengel et al., 2001). Con respecto al trópico existen muchas imprecisiones en cuanto a las verdaderas respuestas de los frutales plantados con relación a la fertilización y comportamiento agronómico de especies y en lo referido a las deficiencias de elementos y requerimientos de una nutrición mineral garante de buenos rendimientos (Avilán et al., 1992). La nutrición de las plantas afecta tanto la calidad interna como la externa, actuando sobre la firmeza y la respiración de los frutos cosechados (Villamizar y Ospina, 1995).
Si la concentración de un elemento nutriente esencial en el tejido vegetal está por debajo del nivel necesario para un óptimo crecimiento, indica que la planta es deficiente en ese elemento, produciendo así una alteración en la ruta metabólica en la que participa dicho elemento, afectando además otros procesos inmediatamente involucrados. Las degeneraciones metabólicas producidas por deficiencias de nutrientes esenciales se manifiestan eventualmente en anormalidades visibles (Epstein y Bloom, 2005).
Samra y Arora (1997) describen el nitrógeno (N) como un elemento de gran influencia en el crecimiento y desarrollo de la planta al promover la producción de ramas, hojas y frutos. En general las plantas deficientes en nitrógeno presentan un pobre crecimiento y una baja productividad. Según Fischer y Angulo (1999), el nitrógeno es el elemento que más influye en la disminución de la producción en plantas de uchuva, su deficiencia se manifiesta en una reducción en número y longitud de las ramas lo que afecta la cantidad y el tamaño de los frutos en formación.
El contenido de fósforo (P) interviene en la formación de los órganos reproductores, razón por la cual su contenido debe ser suficiente en los frutos (Pruiggros y Morin, 1985). Mengel et al. (2001) reportan el fósforo como un elemento que se utiliza en bajas cantidades que, relacionado con la calidad, interviene en la utilización del azúcar y del almidón. El suministro inadecuado de fosfato impide la exportación de triosas fosfatadas del cloroplasto y por ende la síntesis de sacarosa. Además, el efecto de la nutrición con fósforo en la calidad del fruto puede atribuirse a su papel como componente de los fosfolípidos, que son los mayores constituyentes de la membrana celular (Knowles et al., 2001).
De acuerdo con Samra y Arora (1997), el potasio mejora la calidad y duración del cultivo y también alivia las condiciones de estrés. El potasio es activador de muchas enzimas que son esenciales en la fotosíntesis y en la respiración, además activa enzimas necesarias para formar almidón y proteínas y es involucrado en el transporte de los fotoasimilados (Swietlik, 2003). Asimismo, el K es un ión que desempeña un papel fundamental en la osmorregulación celular y su deficiencia produce pérdida de turgencia y marchitamiento, más acentuado cuando hay déficit hídrico (Bonilla, 2000), además de dar firmeza a los tejidos y grosor a las paredes celulares; la deficiencia de potasio genera descomposición del tejido parenquimatoso (Black, 1975).
El calcio (Ca) se usa en la síntesis de nuevas paredes celulares, particularmente en la lámina media que separa las nuevas células divididas; se requiere para un normal funcionamiento de las membranas vegetales y ha sido implicado como segundo mensajero en diferentes respuestas de las plantas tanto en señales ambientales como hormonales (Taiz y Zeiger, 2006). En frutos, el Ca es importante porque regula la maduración, activa ciertas enzimas y afecta las tasas de respiración y producción de etileno (Ferguson y Droback, 1986). El incremento del nivel de Ca es una medida para mejorar la resistencia natural a enfermedades y mantiene la calidad del fruto (Fallahi et al., 1997).
En las células vegetales el magnesio (Mg) cumple un rol específico como activador de enzimas envueltas en la respiración, fotosíntesis y síntesis de ADN y ARN. También forma parte de la molécula de clorofila (Taiz y Zeiger, 2006). Al Mg se le atribuye participación en el desarrollo de frutos (Bonilla y Mahecha, 1990), contribuyendo a la labor de la fructosa 1,6 difosfatasa, la cual regula la síntesis de almidón (Clavijo, 2001), factor que puede ser determinante en el nivel de azúcares y la calidad de los frutos.
El boro (B) juega un rol primario en la biosíntesis y estructura de la pared celular y en la integridad de la membrana plasmática (Marschner, 2002); así mismo se involucra en el transporte de azúcares, la lignificación de la pared celular (Loomis y Durst, 1992), la elongación celular, síntesis de ácidos nucleicos y respuestas hormonales (Taiz y Zeiger, 2006).
Es para considerar que la planta, por regla general, no cuenta con grandes capacidades para almacenar elementos nutricionales por lo que necesita absorberlos constantemente del exterior. Esto sucede en especial para macroelementos como el nitrógeno y fósforo, y la ausencia de estos al poco tiempo desarrollará síntomas de deficiencia morfológicos y fisiológicos (Schopfer y Brennicke, 2006).
Debido a los pocos conocimientos de las deficiencias nutricionales en la uchuva y el alto número de problemas que se presentan en las plantaciones del país por este desconocimiento, el objetivo de este trabajo fue identificar en plantas de uchuva el efecto de una nutrición deficiente en N, P, K, Ca, Mg y B sobre la producción y calidad del fruto.
Materiales y métodos
La investigación se llevó a cabo en la Facultad de Agronomía de la Universidad Nacional de Colombia, sede Bogotá, bajo condiciones de invernadero. Inició el 28 de junio de 2007 con el trasplante de plantas de uchuva ecotipo Colombia, de 1 mes de edad, en materas de plástico con una capacidad de 25 kg; estas se llenaron con arena cuarcítica con un tamaño de grano de 0,5 mm y una conductividad eléctrica (CE) de 0,011 dS·m-1. Se ubicaron las materas con un espacio de 1,5 m y se implementó un diseño completamente al azar con 8 tratamientos y 5 repeticiones, distribuidos así: T0: fertilización completa (testigo); T1: fertilización completa menos nitrógeno (-N); T2: fertilización completa menos fósforo (-P); T3: fertilización completa menos potasio (-K); T4: (-Ca); T5: (-Mg); T6: (-B) y T7: H2O, aplicación de solo agua potable, sin ningún elemento adicional. En el caso sin nitrógeno se adicionó el 15% del nitrógeno aplicado en el tratamiento completo para garantizar un crecimiento mínimo de la planta (Martínez, 1997).
Para evaluar el efecto de los tratamientos se utilizó la solución de Hoagland y Arnon modificada por D.F. Ortega (comunicación personal) (tabla 1), la cual se mezcló para generar soluciones madre en tanques de 40 L; esta se aplicó dos veces por día mediante regadera. Al agua de riego se le realizó un análisis fisicoquímico para verificar que fuera apta para el estudio.
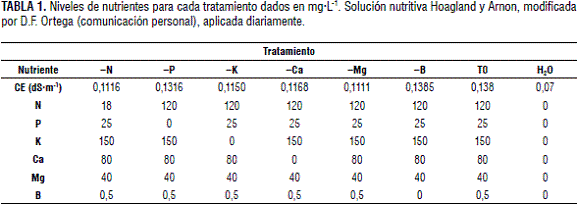
Las fuentes utilizadas fueron: nitrato de amonio líquido, fosfato monopotásico, DAP, nitrato de potasio, sulfato de potasio, nitrato de calcio líquido, Mainstay calcio, sulfato de magnesio, EDTA-Fe 11%, cosmoquel B y molibdato de amonio. El sistema de tutorado fue en ´V´ y no se realizó ningún tipo de poda.
Las evaluaciones se hicieron con una frecuencia de 15 días a partir de la fecha de siembra para las variables número de frutos y rendimiento por planta. La calidad de fruto se evaluó tomando muestras de 10 frutos por planta en el día 90 después de trasplante, registrando sólidos solubles totales (SST), acidez total titulable (ATT) y pH de frutos. Para la determinación de los SST se usó un refractómetro portátil Carl Zeiss; el pH se determinó con un potenciómetro Orion, modelo 420A; la ATT se determinó mediante titulación con NaOH 0,1N hasta pH 8,2 y se expresó como porcentaje de ácido cítrico. Las variables pH y ATT no pudieron evaluarse en los tratamientos -K, -B y H2O, debido a la baja cantidad de frutos generados por las plantas a las cuales se aplicaron dichos tratamientos.
Junto con la calidad de fruto, las variables de producción peso fresco de fruto, peso seco de cáliz y fruto (secado a una temperatura de 72 °C por 42 h), diámetro ecuatorial y longitudinal de frutos se determinaron en el laboratorio de fisiología vegetal de la Facultad de Agronomía, Bogotá.
En el análisis estadístico de todas las variables se utilizó el paquete SAS 9.1 para realizar el análisis de varianza y se compararon promedios mediante la prueba de comparación de Tukey.
Resultados y discusión
Producción
Diámetros ecuatorial y longitudinal de frutos
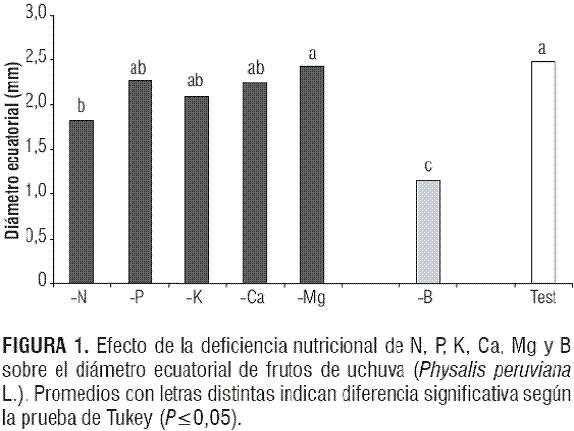
Se presentaron diferencias significativas entre el tratamiento -B y los restantes en estas variables (P<0,0001) para el diámetro ecuatorial y P<0,0116 para el diámetro longitudinal). La deficiencia de boro afectó severamente estos diámetros (figuras 1 y 2), observando promedios de 1,15 y 0,95 cm, respectivamente, que, comparados a los 2,48 y 2,26 cm que arrojó el tratamiento testigo, representan una disminución en 53,62 y 57,96%. Esta drástica reducción del tamaño de los frutos podría estar, sobre todo, originada por la disminución del transporte de azúcares (Loomis y Durst, 1992), y en general, por el efecto negativo de la falta de B en la división y el crecimiento celular (Taiz y Zeiger, 2006; Bonilla, 2008) por lo que lo relaciona con la división y el crecimiento celular por lo que el tamaño del fruto podría haber sido reducido en este estudio. Por otra parte, el B está involucrado en el mantenimiento del potencial de la membrana celular, posiblemente por reacciones con sistemas enzimáticos que alteran el transporte de electrones en la membrana, más concretamente con la permeabilidad selectiva de Na/K. Influye en la incorporación de proteínas y precursores dentro de la pared celular. Un azúcar en la fracción péctica de las paredes celulares forma esteres con ácido bórico o borato (Brown et al., 2002).
También en maracuyá (Freitas et al., 2006) y en fresa (Neilsen y Eaton, 1983) se encontró que plantas deficientes de B disminuyeron significativamente el diámetro de los frutos.
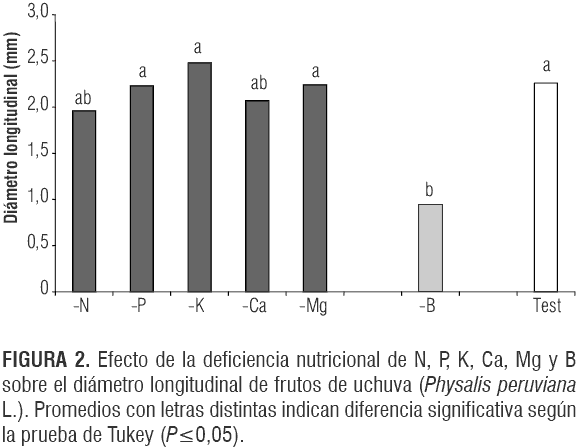
Así mismo, se halló un efecto negativo de la deficiencia de nitrógeno sobre los diámetros ecuatorial y longitudinal de los frutos, aunque en un menor grado que la deficiencia de boro. La deficiencia de N presentó valores promedio de 1,83 y 1,96 cm,respectivamente, que comparados con los valores del tratamiento testigo representaron una reducción apreciable en estas variables del fruto de uchuva. La importancia del N en formación del fruto y en su tamaño fue descrita por Samra y Arora (1997) en mango y Fischer y Angulo (1999) en uchuva, siendo este elemento el segundo más abundante en los frutos (Osterloh et al., 1996). Además, la reducida área foliar de la planta de uchuva en el tratamiento deficiente de N (Martínez y Sarmiento, 2008) podría haber afectado el crecimiento normal del fruto, siendo la restricción de la fuente por un estrés nutricional una limitante para el crecimiento del fruto (Lakso y Goffinet, 2003).
El tratamiento -Mg mostró los mismos tamaños de los frutos del testigo (figuras 1 y 2), lo que coincide con lo reportado por Ogawa (1995) en ciruelo y durazno, pero contrasta con lo observado por Agustí (2003) en cítricos. Aunque el potasio fomenta la expansión celular (Agustí, 2004), su deficiencia no resultó en frutos más pequeños que los del testigo (figuras 1 y 2).
Peso fresco de frutos
También en la variable peso fresco del fruto se encontraron diferencias altamente significativas (P<0,0001) entre los tratamientos con respecto al testigo. La figura 3 muestra que los tratamientos -B, -N y -K afectaron el peso de los frutos en su madurez de cosecha, presentando 0,93; 4,15 y 4,66 g, respectivamente, frente al testigo (7,57 g). Se nota de nuevo la importancia del boro en la formación y crecimiento del fruto, que radica especialmente en el papel de este elemento en la división y elongación celular (Taiz y Zeiger, 2006) y la traslocación de carbohidratos (Loomis y Durst, 1992); de esta manera la expansión del fruto se vio limitada con respecto a plantas con un suministro adecuado de este elemento.
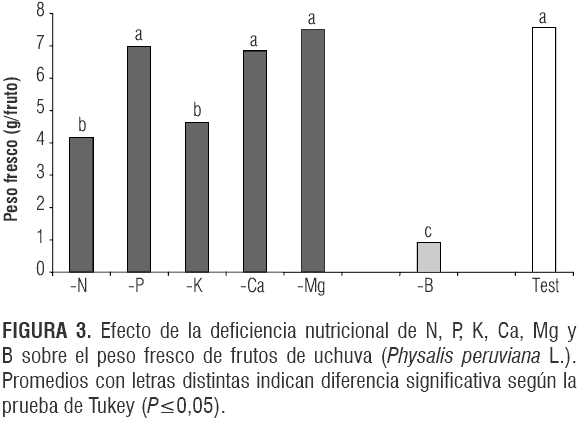
Estos resultados concuerdan con los de Garg y Singh (1975) en uchuva, quienes reportaron que las plantas con deficiencia de N presentaron una reducción del 89,20% y las deficientes en K una reducción del 53,29% en el peso fresco de frutos, con respecto al testigo. También Freitas et al. (2006) observaron en maracuyá que las plantas con deficiencia de potasio y boro registraron los menores pesos frescos de frutos, 158 y 177 g, respectivamente, comparados con el testigo (199 g). Así mismo en maracuyá Carvalho et al. (1999) encontraron frutos de mayor peso en los tratamientos que recibieron K en abundancia.
Las plantas deficientes de Ca produjeron un peso fresco similar al de los frutos del testigo, lo que no concuerda con resultados de Cooman et al. (2005) que reportaron en uchuva una reducción del peso usando soluciones nutritivas sin Ca.
Peso seco del fruto y el cáliz
Hubo diferencias altamente significativas en el peso seco de frutos (P<0,0001) (figura 4). Se puede apreciar que, de los elementos mayores, las deficiencias de nitrógeno y potasio afectaron significativamente el peso seco de los frutos en su madurez de cosecha, con valores de 0,579 y 0,736 g/fruto, respectivamente, en comparación con los 1,093 g/fruto que desarrolló el testigo. El efecto perjudicial de la deficiencia de K y N en la solución nutritiva sobre el peso de los frutos es explicable también porque estos dos elementos son los de mayor abundancia en este órgano reproductivo (Osterloh et al., 1996).
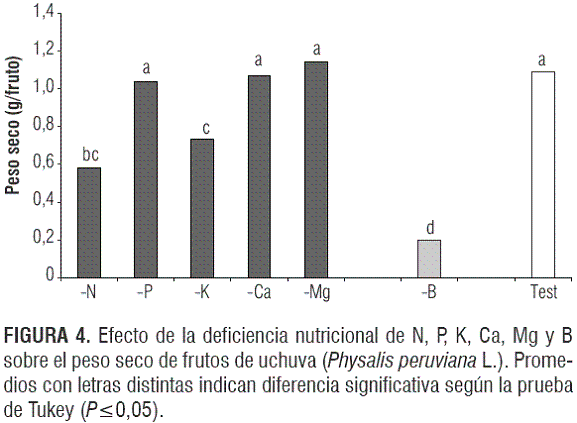
La reducción del peso seco de los frutos con deficiencia de boro fue todavía mayor, con valores altamente más bajos que el testigo y los -N y -K, reduciendo drásticamente su peso seco (81,97%) con respecto a los que se sometieron a una fertilización completa.
La formación característica y el peso del cáliz es importante porque la mayoría de las exportaciones de uchuva se envían con este órgano adherido (Galvis et al., 2005). Hubo un efecto altamente significativo de los tratamientos sobre el peso seco de cáliz según el análisis de varianza (P<0,0001). En la figura 5 se observa que se presentó solamente para el caso del -B una reducción significativa del peso seco del cáliz (0,112 g), comparado con el testigo (0,306 g); así mismo mostró diferencias significativas con los demás tratamientos a excepción de los tratamientos -N y -K.
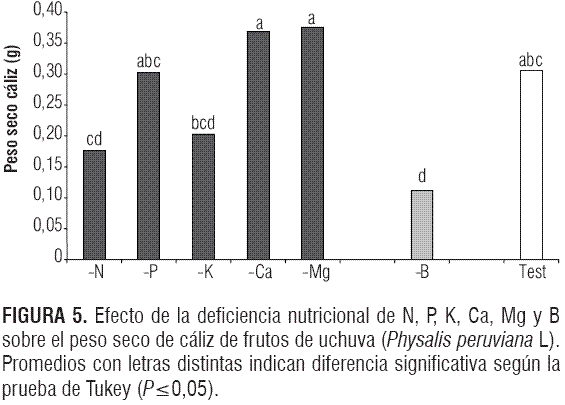
La ausencia de B, según Matoh et al. (1996), disminuye o inhibe el crecimiento de estructuras vegetativas (cáliz) y reproductivas (fruto), debido a la pérdida de la plasticidad y estructura de la pared celular (Loomis y Durst, 1992).
Número de frutos
Desde el inicio de la fructificación hasta los 135 días después del trasplante (ddt) los tratamientos -N, -K y -B presentaron los menores valores para esta variable, desarrollando solo 34, 17 y 27 frutos/planta a los 135 ddt frente al testigo que generó 349 frutos/planta (figura 6). Según Fischer y Angulo (1999) el nitrógeno es el elemento que más influye en la disminución de la producción de frutos en plantas de uchuva, afectando su número y tamaño. La cantidad de frutos para el tratamiento -P se vio reducida en el tiempo, desde el inicio de la fructificación hasta 135 ddt, en que se desarrollaron en promedio 169 frutos, lo que significa una reducción del 51,5% frente al testigo. En contraste, los tratamientos -Ca y -Mg tuvieron un comportamiento similar al testigo hasta el día 120 ddt: desarrollaron alrededor de 200 frutos/planta. Solo 135 ddt mostraron diferencias significativas, cuando se presentó una reducción de 25,5% con respecto al testigo. El menor número de frutos concuerda con el poco crecimiento longitudinal de las ramas generativas de los tratamientos afectados por las deficiencias nutricionales en la solución nutritiva, comparados con el testigo (Martínez y Sarmiento, 2008), teniendo en cuenta que la uchuva forma en cada nudo de la rama reproductiva un fruto (Fischer, 2000).
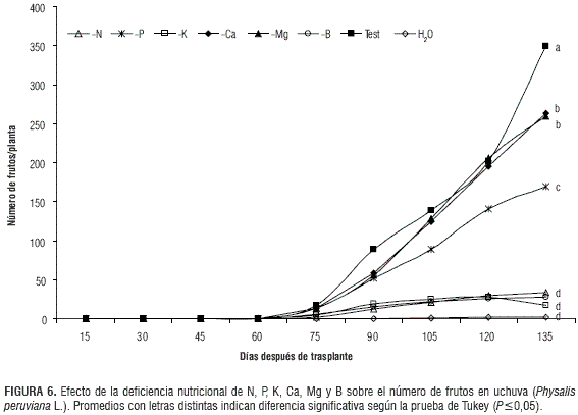
Resultados similares fueron encontrados por Garg y Singh (1975) en uchuva, donde las plantas deficientes en N, P y K tuvieron una reducción significativa en el número de frutos, desarrollando un promedio de 3,5; 6,5 y 11,5 frutos/planta, respectivamente, en comparación con el testigo (20,8 frutos/planta). También en maracuyá, Freitas et al. (2006) reportaron que plantas cultivadas con deficiencia de N, P, K, Ca y B redujeron significativamente el número de frutos con respecto al testigo. Araújo et al. (2005), estudiando los efectos de la nutrición potásica sobre el crecimiento y producción del maracuyá, verificaron que un incremento de la concentración de potasio en la solución nutritiva provocó un aumento linear en el número de frutos/planta.
Con respecto al boro, Keerati-Kasikorn et al. (1993) observaron en maní aborto de frutos y reducidos tamaños de estos. Por otra parte, las plantas deficientes en este elemento presentaron necrosis de la yema terminal (Taiz y Zeiger, 2006) lo que afectó el desarrollo característico de la planta (Martínez y Sarmiento, 2008) perjudicando la producción de los frutos.
Rendimiento
El rendimiento por planta, que se compone del número de frutos (figura 6) con su respectivo peso fresco (figura 3), fue afectado en la misma forma por las deficiencias nutricionales; se observa en la figura 7 que los tratamientos -K, -B y -N presentaron los menores valores desde el inicio de la producción hasta los 135 ddt. Esto significó una disminución de 98,98; 96,83 y 94,68%, respectivamente, con respecto al testigo, en el cual se cosechó al final del ensayo 2.649,49 g/planta. En el tratamiento -P se observó desde el día 120 una tendencia a la estabilización que contrasta con el testigo, el cual sigue un comportamiento exponencial. La deficiencia de P tuvo una influencia negativa sobre el rendimiento desde el día 105 hasta el 135, en que se obtuvieron 1.884,04 g/planta, que equivale a una reducción del 55,31% frente al testigo. Las deficiencias de Ca y Mg presentaron un comportamiento similar al testigo hasta el día 120, sin embargo a los 135 días se observó una reducción de 31,96 y 26,63%, respectivamente, en el rendimiento frente al testigo.
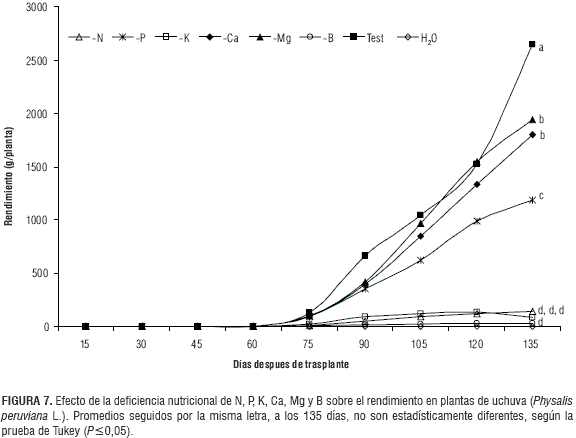
Estos resultados coinciden con los de Gordillo et al. (2004) que evaluaron la fertilización en una plantación de uchuva en San Raimundo (Cundinamarca) y encontraron rendimientos de frutos bajos, de 8,13 kg/planta cuando se aplicaron menores dosis de potasio y de boro (9,65 kg/ planta), en comparación con la producción del testigo (10,93 kg/planta).
Singh y Pandey (1977) hallaron en uchuva que el menor rendimiento lo presentaron las plantas con la menor aplicación de N-P-K (20-40-40 kg·ha-1), con promedios de 32,35 y 39,93 qtl/ha en dos años de evaluación, valores considerablemente más bajos que los de las plantas que se fertilizaron con la dosis más alta (80-80-80 kg·ha-1) las cuales produjeron 47,05 y 56,95 qtl/ha.
En el caso que existan concentraciones foliares <1% del potasio (en este ensayo hubo solamente 0,48% K; Martínez y Sarmiento, 2008) se disminuye severamente la fotosíntesis y a su vez el tamaño del fruto (figuras 1 y 2) y el rendimiento (figura 6) en general (Neilsen y Neilsen, 2003).
Con respecto al Ca, similares reducciones del rendimiento encontraron Cooman et al. (2005) en uchuva, usando soluciones nutritivas carentes de este elemento.
Calidad de fruto
Sólidos solubles totales (SST)
El análisis de varianza para SST mostró una diferencia significativa entre tratamientos (P<0,0014). La ausencia de boro disminuyó significativamente la concentración de grados Brix (8,92) frente al testigo (14,86) (figura 8). Esto, supuestamente, se debe a que el B influye indirectamente el metabolismo de azúcares facilitando el transporte desde las fuentes hacia sus vertederos (Marschner, 2002). Este autor afirma claramente que en condición de deficiencia, la exportación de fotoasimilados desde las hojas se afecta por la falta de actividad vertedero (como es el crecimiento del fruto de uchuva). Aparte del B, el tratamiento -K mostró una tendencia a reducir los SST en el jugo del fruto, encontrándose 2,83 °Brix, que significó un contenido de SST 13,6% menor que el del testigo, pero esta diferencia no fue significativa. En maracuyá un incremento en la nutrición potásica promovió un aumento lineal en la concentración de SST en el jugo del fruto (Carvalho et al., 1999). La deficiencia de nitrógeno no afectó la concentración de los SST lo que concuerda con lo reportado por Link (1980) para el manzano.
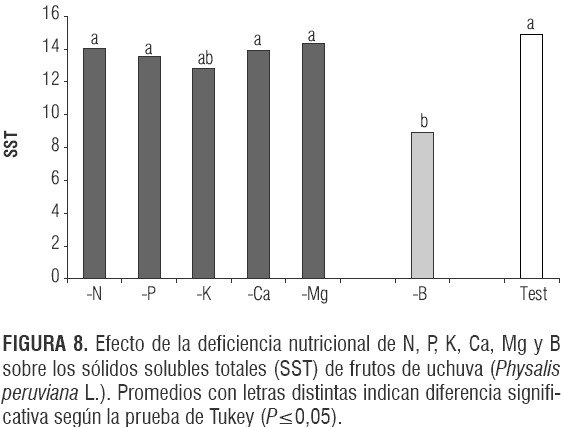
Acidez total titulable (ATT)
En el análisis de varianza la F significativa (P<0,0102) evidencia que existió efecto en la ATT de los frutos a causa de los tratamientos; sin embargo, no hubo diferencias signi- ficativas entre el testigo y las deficiencias nutricionales. El tratamiento -Ca presentó un menor contenido de ácidos que frutos en plantas con deficiencias fosfóricas (figura 9). En contraste, Link (1980) encontró en diferentes variedades de manzano, que recibieron fertilizaciones bajas en nitrógeno, concentraciones altas de ATT y concluyeron que la descomposición de los ácidos en el fruto es más lenta con poca fertilización nitrogenada. Singh y Pandey (1977) detectaron en uchuvas la ATT más baja en plantas que recibieron fertilización de N-P-K baja comparada con aquellas que recibieron dosis altas.
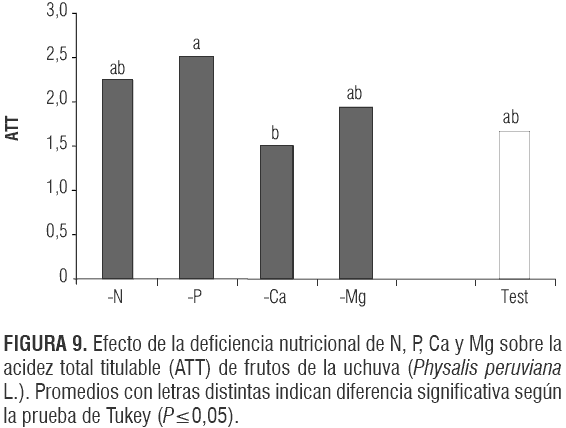
Siendo el Ca sumamente importante para la integridad de las membranas de las paredes celulares, una deficiencia de este causará un aumento en la respiración del fruto (Dris, 2001), elevando la permeabilidad de estas (Stutte, 1995), lo que podría haber jugado un papel importante en la menor concentración de los ácidos en el tratamiento -Ca , que, posiblemente, fueron usados en la respiración del fruto (Kays, 2004). Con respecto al aumento de la ATT por la deficiencia del P se podría suponer que hubo una influencia por el retardo en la formación del fruto (Marschner, 2002) y así una reducción tardía de los ácidos en este órgano, lo que muestra también la figura 10 con una relación de madurez baja en este tratamiento.
Las plantas deficientes en Mg produjeron frutos con los mismos niveles de ATT y SST que los del testigo, contrario a lo reportado por Agustí (2003) en cítricos que desarrollaron frutos con menores contenidos de estos dos ingredientes con una baja fertilización magnésica. Posiblemente las uchuvas no son tan susceptibles a las deficiencias magnésicas relacionadas con el contenido de estos dos ingredientes en el fruto comparado con los cítricos (Ritenour et al., 2002).
Relación de madurez (RM)
En la gran mayoría de los frutos, incluyendo la uchuva (Galvis et al., 2005), durante la maduración se disminuye la proporción de los ácidos y aumentan los azúcares; a mayor RM (SST/ATT) es más alto el grado de madurez del producto (Parra y Hernández, 2005).
La RM del testigo, 9,0 (figura 10), fue un poco mayor que la reportada por Novoa et al. (2006) en uchuva (8,0). Los tratamientos -N y -P presentaron las menores RM (6,2 y 5,9, respectivamente) (figura 10), debido a que estas defi- ciencias tienden a retardar los procesos de maduración del fruto (Yoshioka et al., 1989; Link, 1980; Marschner, 2002). El Ca que no está involucrado en la maduración del fruto (Ferguson et al., 1999) muestra una RM similar a la del testigo.
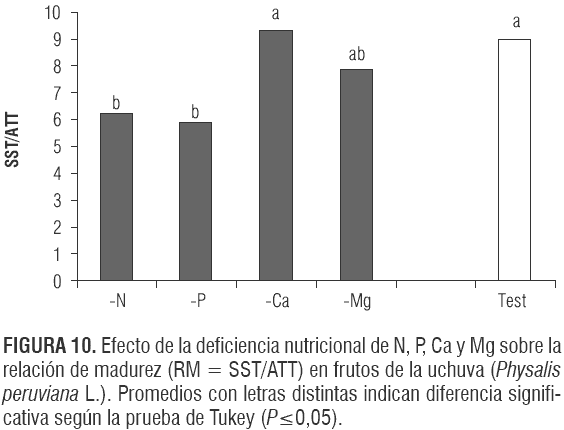
pH
Los tratamientos no presentaron efectos estadísticos (P<0,5940) sobre el pH de los frutos que osciló de 3,5 a 3,7, es decir las deficiencias nutricionales conservaron la característica ácida de los frutos.
Conclusiones
El elemento faltante que más afectó la producción de las uchuvas fue el boro que disminuyó el tamaño de los frutos, su peso fresco y seco y también la masa seca de los cálices, indicando su importancia en la traslocación de los carbohidratos.
También las deficiencias de K y N disminuyeron el peso fresco y seco de los frutos, siendo estos dos elementos los de mayor abundancia en este órgano.
La producción de los frutos disminuyó altamente por las deficiencias de N, K, B y P, coincidiendo con el menor crecimiento de las ramas productivas en estos tratamientos.
La calidad del fruto afectó la deficiencia de B referente a la menor concentración de los sólidos solubles, mientras la del Ca disminuyó la acidez titulable.
En general, las deficiencias nutricionales, excepto la del boro, tuvieron un bajo efecto sobre la calidad interna de los frutos de uchuva, lo que confirma que la mayoría de los factores que controlan su calidad son fijados genéticamente (Mengel et al., 2001).
Agradecimientos
Los autores agradecen a las empresas Monómeros Colombo-Venezolanos, K+S Kali GmbH y C.I. Frutas Comerciales por su apoyo financiero en la realización de esta investigación.
Literatura citada
Agustí, M. 2003. Citricultura. Ediciones Mundi-Prensa, Madrid. 422 p. [ Links ]
Agustí, M. 2004. Fruticultura. Ediciones Mundi-Prensa, Madrid. 493 p. [ Links ]
Araújo, R., C.H. Bruckner, H.P. Martínez, L.C. Salomão, V.H. Venegas, J.M. Díaz, W.E. Pereira y J.A. Souza. 2005. Crescimento e produção do maracujazeiro-amarelo em resposta à nutrição potássica. Rev. Bras. Frutic. 27(1), 128-131. [ Links ]
Avilán, L., L. Leal y D. Batista. 1992. Manual de fruticultura. Principios y manejo de la producción. Segunda edición. Editorial América, Caracas. pp. 1387-1401. [ Links ]
Bonilla, I. 2000. Introducción a la nutrición mineral de las plantas. pp. 83-91. En: Azcon-Bieto, J. y M. Talón (eds.). Fundamentos de fisiología vegetal. McGraw-Hill Interamericana, Madrid. [ Links ]
Bonilla, I. 2008. Introducción a la nutrición mineral de las plantas. Los elementos minerales. pp. 103-141. En: Azcon-Bieto, J. y M. Talón (eds.). Fundamentos de fisiología vegetal. McGraw-Hill Interamericana, Madrid. [ Links ]
Bonilla, G. y S. Mahecha. 1990. Efecto de la fertilización edáfica y foliar sobre la producción del mango (Mangifera indica L.). Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 82 p. [ Links ]
Black, C. 1975. Relaciones suelo-planta. Tomo II. Ed. Hemisferio Sur, Buenos Aires. 677 p. [ Links ]
Brown, P., N. Bellaloui, M. Wimmer, E. Bassil, J. Ruiz, H. Hu, H. Pfeffer, F. Dannel y V. Römheld. 2002. Boron in plant biology. Plant Biol. 4, 205-223. [ Links ]
Cabezas, M., T. Botía y L. Medina. 2002. Determinación de síntomas por deficiencia inducida de nutrimientos en lulo (Solanum quitoense Lamb). pp. 176-181. Memorias IV Seminario Nacional de Frutales de Clima Frío Moderado, Medellín. [ Links ]
Carvalho, A.J., P.H. Monnerat, D.P. Martins y S. Bernardo. 1999. Produtividade e qualidade do maracujazeiro-amarelo em resposta à adubação potássica sob lâminas de irrigação. Rev. Bras. Frutic. 21(3), 333-337. [ Links ]
Clavijo, J. 2001. Metabolismo de los nutrientes en las plantas. pp. 13-28. En: Sociedad Colombiana de la Ciencia del suelo. Fertilidad de suelos. Diagnóstico y control. Segunda edición. Sociedad Colombiana de la Ciencia del Suelo, Bogotá. 524 p. [ Links ]
Cooman, A., C. Torres y G. Fischer. 2005. Determinación de las causas del rajado del fruto de uchuva (Physalis peruviana L.) bajo cubierta. II. Efecto de la oferta de calcio, boro y cobre. Agron. Colomb. 23, 74-82. [ Links ]
Dris, R. 2001. Influence of calcium nutrition on the quality and postharvest behavior of apples. pp. 175-186. En: Dris, R., R. Niskanen y S.M. Jain (eds.). Crop management and postharvest handling of horticultural products. Vol. 1. Quality management. Science Publishers, Enfield, NH. 364 p. [ Links ]
Epstein, E. y A.J. Bloom. 2005. Mineral nutrition of plants: principles and perspectives. Sinauer Associates, Sunderland, MA. 400 p. [ Links ]
Fallahi, E., W. Conway, K.D. Hickey y C.E. Sams. 1997. The role of calcium and nitrogen in postharvest quality and disease resistance of apples. HortScience 32, 831-835. [ Links ]
Ferguson, I.B. y B.K. Droback. 1986. Calcium and the regulation of plant growth and senescence. HortScience 23, 262-266. [ Links ]
Ferguson, I., R. Volz y A. Wolf. 1999. Preharvest factors affecting physiological disorders of fruit. Postharvest Biol. Technol. 15, 255-262. [ Links ]
Fischer, G. 2000. Crecimiento y desarrollo. pp. 9-26. En: Flórez, V.J., G. Fischer y A.D. Sora (eds.). Producción, poscosecha y exportación de la uchuva (Physalis peruviana L.). Unibiblos, Universidad Nacional de Colombia, Bogotá. 175 p. [ Links ]
Fischer, G. y R. Angulo. 1999. Los frutales de clima frío en Colombia. La uchuva. Ventana al Campo Andino 2(1), 3-6. [ Links ]
Freitas, M., P.H. Monnerat, L.G. Rocha y A. Carvalho. 2006. De- ficiência de macronutrientes e boro em maracujazeiro doce: qualidade dos frutos. Rev. Bras. de Frutic. 28(3), 492-496. [ Links ]
Galvis, J.A., G. Fischer y O.P. Gordillo. 2005. Cosecha y poscosecha de la uchuva. pp. 165-190. En: Fischer, G., D. Miranda, W. Piedrahita y J. Romero (eds.). Avances en cultivo, poscosecha y exportación de la uchuva (Physalis peruviana L.) en Colombia. Unibiblos, Universidad Nacional de Colombia, Bogotá. 221 p. [ Links ]
Garg, R.C. y S.K. Singh. 1975. Primary nutrient deficiencies in cape gooseberry (Physalis peruviana L.). Progressive Hort. 7(2), 53-58. [ Links ]
Gordillo, O., G. Fischer y R. Guerrero. 2004. Efecto del riego y de la fertilización sobre la incidencia del rajado en frutos de uchuva (Physalis peruviana L.) en la zona de Silvania (Cundinamarca). Agron. Colomb. 22(1), 53-62. [ Links ]
Kays, S. 2004. Postharvest biology. Exon Press, Athens, Georgia. 568 p. [ Links ]
Keerati-Kasikorn, P., R.W. Bell, P. Panya, R. Gilmour y J.F. Loneragan. 1993. Comparison of seed yield and quality of peanut (Arachis hypogaea L.) cultivars in low fertility soils and their response to boron and complete fertiliser. pp. 409-412. En: Barrow, N.J. (ed.). Plant nutrition - from genetic engineering to field practice. Developments in plant and soil sciences 54. Kluwer Academic Publishers, Dordrecht. [ Links ]
Knowles, L., R. Trimble y R. Knowles. 2001. Phosphorus status affects postharvest respiration, membrane permeability and lipid chemistry of European seedless cucumber fruit (Cucumis sativus L.). Postharv. Biol. Technol. 21, 179-188. [ Links ]
Lakso, A.N. y M.C. Goffinet. 2003. Fruit growth patterns. pp. 91-101. En: Baugher, T.A. y S. Singha (eds.). Concise encyclopedia of temperature tree fruit. Food Product Press, Nueva York. 387 p. [ Links ]
Link, H. 1980. Effects of nitrogen supply on some components of apple fruit quality. Acta Hort. 92, 285. [ Links ]
Loomis, W.D. y R.W. Durst. 1992. Chemistry and biology of boron. BioFactors 3, 229-239. [ Links ]
Marschner, H. 2002. Mineral nutrition of higher plants. Academic Press, Londres. 889 p. [ Links ]
Martínez, F.E. y J. Sarmiento. 2008. Estudio de los síntomas de de- ficiencia de N, P, K, Ca, Mg y B en plantas de uchuva (Physalis peruviana L.). Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. [ Links ]
Martínez, M. 1997. Mineral nutrient deficiency in plantain. Ph. D. thesis. Hohenheim tropical agricultural series, University of Hohenheim, Stuttgart. [ Links ]
Matoh, T., S. Kawaguchi y M. Kobayashi. 1996. Ubiquity of a boraterhamno-galacturonan II complex in the cell walls of higher plants. Plant Cell Physiol. 37, 636-640. [ Links ]
Mengel, K., E.A. Kirkby, H. Kosegarten y T. Appel. 2001. Principles of plant nutrition. Kluwer Academic Publishers, Dordrecht. 849 p. [ Links ]
Neilsen, G.H. y D. Neilsen. 2003. Nutritional requirements of apple. pp. 267-302. En: Ferree, D.C. e I.J. Warrington (eds.). Apples - Botany, production and uses. CABI Publishing, Wallingford, UK. 660 p. [ Links ]
Neilsen, B.V. y G.W. Eaton. 1983. Effects of boron nutrition upon strawberry yield components. HortScience 18, 932-934. [ Links ]
Novoa, R.H., M. Bojacá, J.A. Galvis y G. Fischer. 2006. La madurez del fruto y el secado del cáliz influyen en el comportamiento poscosecha de la uchuva (Physalis peruviana L.) almacenada a 12 °C. Agron. Colomb. 24(1), 68-76. [ Links ]
Ogawa, J.M. (ed.). 1995. Compendium of stone fruit diseases. APS Press, St. Paul, Minnesota. pp. 85-91. [ Links ]
Osterloh, A., G. Ebert, W.-H. Held, H. Schulz y E. Urban. 1996. Lagerung von Obst und Südfrüchten. Verlag Ulmer, Stuttgart. 253 p. [ Links ]
Parra, A. y J.E. Hernández. 2005. Fisiología poscosecha de frutas. Facultad de Ingeniería, Universidad Nacional de Colombia, Bogotá. 64 p. [ Links ]
Pruiggros, J. y C. Morin. 1985. Nutrición de los cítricos. pp. 249-410. En: Morin, C. (ed). Cultivo de cítricos. Instituto Interamericano de Cooperación para la Agricultura, Costa Rica. 598 p. [ Links ]
Ritenour, M., W. Wardowski y D. Tucker. 2002. Postharvest effects of water and nutrients. pp. 117-120. En: Boman, B.J. (ed.). Water and Florida Citrus: use, regulation, irrigation systems and management. University of Florida, Institute of Food and Agricultural Sciences, Gainesville. [ Links ]
Samra, J.S. y Y.K. Arora. 1997. Mineral nutrition. pp. 175-201. En: Litz, R.E. (ed.). The mango: botany, production and uses. CAB International. 587 p. [ Links ]
Sanabria, S. 2005. Situación actual de la uchuva en Colombia. pp. 1-8. En: Fischer, G., D. Miranda, W. Piedrahita y J. Romero (eds.). 2005. Avances en cultivo, poscosecha y exportación de la uchuva (Physalis peruviana L.) en Colombia. Unibiblos, Universidad Nacional de Colombia, Bogotá. 221 p. [ Links ]
Schopfer, P. y A. Brennicke. 2006. Pflanzenphysiologie. Elsevier GmbH, Munich. 700 p. [ Links ]
Singh, U.R., I.C. Pandey y R.S. Prasad. 1977. Effect of N, P and K on growth, yield and quality of cape-gooseberry. Punjap Hort. J. 17(3-4), 148-151. [ Links ]
Stutte, C. 1995. Laboratory evaluation of TOG NH4 on cotton, tomato, and soybean. Crop Sci. 35, 1069-1073. [ Links ]
Swietlik, D. 2003. Plant nutrition. pp. 251-257. En: Baugher, T.A. y S. Singha (eds.). Concise encyclopedia of temperature tree fruit. Food Product Press, Nueva York. 387 p. [ Links ]
Taiz, L. y E. Zeiger. 2006. Plant physiology. 2nd ed. Sinauer Associates, Sunderland, Massachusetts. 764 p. [ Links ]
Villamizar, F. y J. Ospina. 1995. Frutas y hortalizas. Manejo tecnológico en poscosecha. SENA, Bogotá. pp. 19-21. [ Links ]
Yoshioka, A.H., K. Aoba, M. Fukumoto y K. Fujimoto. 1989. Effect of nitrate and potasium nutrition on the storability of apple fruit. J. Japan. Soc. Hort. Sci. 58, 475-481. [ Links ]

