










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.27 n.1 Bogotá jan./abr. 2009
PROPAGACIÓN Y CULTIVO DE TEJIDOS
María Isabel Hernández P.1, Mario Lobo A.2, 5, Clara Inés Medina C.3, José Régulo Cartagena V.4 y Oscar Arturo Delgado P.3
1 Departamento de Ciencias Agrícolas y Pecuarias, Facultad de Ciencias Agrarias y del Medio Ambiente, Universidad Francisco de Paula Santander, San José de Cúcuta (Colombia).
2 Centro de Investigación La Selva, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Rionegro (Colombia).
3 Centro de Investigación La Selva, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Rionegro (Colombia).
4 Departamento de Ciencias Agronómicas, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Medellín.
5 Autor de correspondencia. pnrgvlobo@gmail.com
Fecha de recepción: 27 de octubre de 2008. Aceptado para publicación: 19 de febrero de 2009
RESUMEN
Se categorizaron la germinación y latencia de semillas de mortiño y se desarrolló un protocolo de remoción de esta última, aspectos que son básicos para procesos de conservación, siembra y fitomejoramiento de la especie. No se detectó latencia exógena, con imbibición de agua, por parte de las semillas en un sustrato húmedo. Las simientes exhibieron una respuesta fotoblástica positiva a la luz. Se determinó, mediante la aplicación de tetrazolio, que 15,8% de las semillas eran no viables, con 63,0% de semillas con germinación y 21,2% viables latentes. La germinación y remoción de la latencia fueron significativamente superiores a 20°C que a 25°C. La latencia de la semilla fue eliminada, casi por completo, bajo un régimen de luz blanca constante, a 20°C, con aplicación de GA3 en concentraciones de 500, 1.000 y 2.000 mg L-1. El bloqueo de la germinación se clasificó como latencia fisiológica no profunda.
Palabras clave: especies promisorias, semillas fotoblásticas, tetrazolio, fisiología de semillas, Colombia.
ABSTRACT
After characterizing germination and dormancy of Andean blueberry, a dormancy release procedure was developed. These are basic aspects of germplasm conservation, planting and improvement. The seeds did not display exogenous dormancy under water imbibition in a moist substrate. However, a positive photoblastic response was observed. A tetrazolium test indicated that 15.8% of the seeds were n viable; 21.2% were dormant viable, and 63% germinated. Germination and dormancy release at 20°C were higher than at 25°C. Seed dormancy was almost completely released after 500; 1,000 and 2,000 mg L-1 GA3 applications under constant white light at 20°C. Dormancy was classified as non deep physiological.
Key words: promissory species, photoblastic seeds, tetrazolium, seed physiology, Colombia.
Introducción
El mortiño o agraz (Vaccinium meridionale Swartz) pertenece a la familia Ericaceae, tribu Vacciniae, conjunto que en el neotrópico se concentra en los Andes, en ambientes húmedos de montaña, con elevado número de especies, en Colombia y Ecuador, entre los 1.000 y 3.000 msnm (Luteyn, 2002). Esta diversidad ha sido atribuida a los suelos y tipos de vegetación, como consecuencia de la orografía de la zona (Gentry, 1988; Luteyn, 2002). En Colombia se han reportado en este género V. meridionale, V. floribundum y V. corymbodendron (Luteyn, 2002), con consumo de los frutos de los dos primeros (Romero, 1961).
La planta, considerada como relegada, esto es, taxa ignorados por la ciencia y el desarrollo, con empleo en las áreas de adaptación (Hammer et al., 2001), ha pasado a ser promisoria, como secuela del incremento en la demanda nacional, por su consumo en repostería, jugos, helados, bebidas alcohólicas, etc. (Corantioquia, 2003) y de un uso creciente de sus frutos en los países desarrollados, por los beneficios de la fruta en la nutrición y la salud, de las especies cultivadas y silvestres del género (Taruscio et al., 2004).
En Colombia, el mortiño se encuentra en poblaciones silvestres de los bosques alto andinos, entre los 2.300 y 3.500 msnm (Patiño y Ligarreto, 2006), intercalado frecuentemente con plantaciones de coníferas. Esto favorece las asociaciones micorrizógenas de la planta, y su adaptación a suelos ácidos (USDA, Forest Service, s.f.). Las poblaciones han sufrido procesos de pérdida por deforestación, reconversión productiva, fragmentación, perturbación por la extracción de la llamada "tierra de capote", lo que provoca daños en el sistema radical, deterioro y muerte de individuos por prácticas inadecuadas de cosecha y el corte de ramas para su utilización en floristería (Corantioquia, 2003).
El Informe del Estado de los Recursos Filogenéticos para la Alimentación y la Agricultura (FAO, 1996) indicó que las pérdidas de diversidad genética eran grandes y continuas. Por ello Jarvis et al. (2005) reportaron que la preservación de los acervos genéticos relacionados con los cultivos era importante, y Guarino et al. (2001) indicaron que los conjuntos genéticos de los silvestres han sido escasamente observados, colectados o descritos. Jarvis et al. (2005) señalaron que estas especies son las más amenazadas por su rareza, endemismo y falta general de conocimiento sobre ellas. Por lo anterior, es importante conservar la variabilidad del mortiño para reforestación y desarrollo como alternativa productiva. Shoen y Brown (2001) afirmaron que a medida que los hábitat naturales desaparecen, hay una tendencia de mantener las especies silvestres vegetales en bancos de semillas.
La conservación ex situ del mortiño requiere conocer su biología reproductiva. Esta es una planta clonal que produce individuos genéticamente iguales por reproducción vegetativa (McGlaughling y Friar, 2007), cuya estructura genética es compleja, con mezcla de plantas provenientes de reproducción sexual y asexual (Ruggiero et al., 2005). Por ello es conveniente, aparte de la colección de campo, con pocas plantas por deme, mantener su semilla, para una representación mayor de la variabilidad genética de la metapoblación.
La conservación de la semilla, a largo plazo, requiere que esta tolere desecación hasta el 5 a 10% de humedad. USDA, Forest Service (s.f.) indicó que, pese a la falta de estudios de almacenamiento a largo plazo de semillas de los Vaccinium de bayas azules, que incluyen el mortiño, había información que sugería un comportamiento ortodoxo; adicionalmente Darrow y Scott (1954) afirmaron que las semillas de estos eran ortodoxas. En Vaccinium meridionale, Castañeda (2006) reportó comportamiento ortodoxo de las simientes, lo cual fue corroborado por Hernández (2008). Igualmente, Hill y Vander (2005), en estudios con diversos Vaccinium, concluyeron que las semillas eran ortodoxas, con periodos de viabilidad de 17 a 20 años en los bancos de semilla del suelo.
En plantas silvestres son importantes, para su conservación y siembra, la germinación y la latencia de las semillas. Para Finch-Savage y Leubner-Metzger (2006), la segunda es una propiedad innata que define las condiciones para su germinación, dependiente de la constitución genética, con influencia ambiental significativa. Esto, según los autores mencionados, es un aspecto adaptativo para evitar condiciones desfavorables durante el establecimiento y crecimiento posterior de las plántulas. En diversos taxa de Vaccinium, como V. angustifolium, V. myrtillus, V. ulginosum y V. vitisidaea, se ha reportado latencia fisiológica (Baskin y Baskin, 2001).
También es importante para la germinación, en algunas especies, el efecto inductor de la luz, lo cual se denomina fotoblastismo; este permite el mantenimiento de la latencia en las simientes enterradas en el suelo (Sawada et al., 2008). Se ha encontrado que las semillas de varias especies de Vaccinium son fotoblásticamente positivas, precisando varias horas de luz diarias para germinar (Giba et al., 1995), con información de que las semillas de V. myrtillus no germinan en completa oscuridad (Baskin et al., 2000).
Con base en lo expuesto, se realizó la presente investigación, con el fin de elucidar el comportamiento germinativo de semillas de V. meridionale, categorizar la latencia existente en estas y desarrollar protocolos de germinación y rompimiento del bloqueo de la germinación.
Materiales y métodos
Localización
El estudio se realizó en el Centro de Investigación "La Selva", de la Corporación Colombiana de Investigación Agropecuaria (Corpoica), Rionegro, Colombia a 2.120 msnm, con temperatura promedio de 17°C, HR de 78% y precipitación promedio anual de 1.700 mm, en la zona ecológica bosque húmedo montano bajo (Espinal, 1977) y el Laboratorio de Semillas, con temperaturas entre 17 y 20°C.
Material biológico
La semilla se extrajo de frutos de una población silvestre de V. meridionale del Parque Arví, vereda Santa Elena, Medellín, Colombia y de plantas de la colección de campo, del Centro de Investigación "La Selva".
Extracción de la semilla
Se hizo a partir de frutos maduros, con coloración homogénea morada oscura en la epidermis. A estas bayas se les adicionó agua y se les aplicó presión manual, con el fin de separar la pulpa. Posteriormente se lavaron en un chorro de agua con la ayuda de un cedazo, hasta obtener semillas completamente limpias. Estas se secaron a 17°C y 78% de humedad relativa, durante 48 h, de acuerdo con las recomendaciones de Medina et al. (2008).
Obtención de curvas de secado de las semillas
La finalidad fue obtener el porcentaje de humedad en las simientes utilizado para procesos de almacenamiento a largo plazo (5%). Para ello, se emplearon cámaras herméticas, con evaluación de tres relaciones de peso semilla:sílica gel, 1:1, 1:2 y 1:3. Las cámaras se ubicaron en un cuarto a 25±4°C, con 14% de HR.
Se monitoreó la pérdida de humedad cada hora, tomando el peso de las muestras, hasta obtener el contenido de humedad en equilibrio, CHE, como porcentaje, un valor en que se igualan la humedad en las semillas y la del ambiente (Victoria, 2005). Cada tratamiento constó de cinco repeticiones, las cuales se mantuvieron en bolsas de papel, en las cámaras. Adicionalmente, se tomaron muestras aleatorias de las semillas en secado para determinar el contenido de humedad empleando estufas y gravimetría, de acuerdo con la metodología propuesta por ISTA (1999).
Protocolo de tinción con tetrazolio
Se estandarizó un protocolo para V. meridionale. Para ello, se evaluaron concentraciones del producto del 0,5 y 1,0%, durante 1, 2 y 3 h a 30°C y 24 h a temperatura ambiente, en condiciones de completa oscuridad. Los procedimientos se aplicaron a 10 semillas, con tres repeticiones. Estas se prehumedecieron durante 24 h y luego se evaluaron tres tipos de cortes: longitudinal, punzamiento de la testa y extracción completa del embrión. El tetrazolio interactúa con los procesos de reducción y oxidación de las células vivas en la semilla, cambiando de color las formas hidrogenadas, lo que hace posible distinguir las partes vivas (coloreadas) de aquellas muertas (incoloras) (Poulsen et al., 2006).
Para definir patrones de tinción relacionados con viabilidad, se incluyeron semillas en un microondas durante un min, para matar los embriones. Luego, a la mitad de éstas se les aplicó una solución al 1% de tetrazolio, durante 24 h en oscuridad, y la otra mitad se utilizó para realizar pruebas de germinación. Lo anterior permitió observar tanto la intensidad de la coloración como la ubicación de la misma dentro de las estructuras, elucidar la intensidad y diferenciar patrones de tinción entre semillas viables y no viables.
Categorización de la germinación
Se realizó en germinadores a 21°C y con condiciones de luz:oscuridad de 12:12 h, con el empleo de lámparas de luz blanca, de 20 W 110-120 V/50-60 Hz. Las unidades experimentales constaron de 100 semillas, con 4 repeticiones, éstas se colocaron en cajas Petri con papel filtro, humedecido periódicamente. El ensayo se llevó a cabo durante 30 d, tiempo recomendado por Baskin y Baskin (2001) para diferenciar semillas germinables no latentes y latentes. A las semillas no germinadas se les aplicó el protocolo de tetrazolio, desarrollado en esta investigación, para diferenciar las latentes de las no viables.
Determinación de imbibición de las semillas
La prueba se realizó para determinar la permeabilidad de las semillas al agua (Baskin y Baskin, 2001). Para ello, se tomaron cinco repeticiones de 0,5 g de semilla con humedad de 13%. Estas se colocaron en cajas Petri con papel filtro humedecido en agua destilada. Cada hora, durante 8 h, se tomaron muestras, a las cuales se les eliminó el agua superficial con la ayuda de servilletas y se les registró el peso. El incremento de este indica que las semillas son permeables, y lo contrario indica la presencia de barreras a la entrada del agua (Bansal et al., 1980).
Evaluación de protocolos de rompimiento de latencia
Tomando como referente reportes de latencia fisiológica en especies de Vaccinium (Baskin y Baskin, 2001), se evaluó el efecto promotor de la germinación del ácido giberélico (GA3) en concentraciones de 500, 1.000 y 2.000 mg L-1, más un testigo sin aplicación de este. El estudio se realizó en condiciones de oscuridad e iluminación constantes, por el efecto promotor de la luz en la germinación en diversos Vaccinium (Giba et al., 1995) y se evaluaron dos regímenes de temperatura, 20°C y 25°C. Se empleó un diseño completamente al azar, con cuatro repeticiones y unidades experimentales de 100 semillas, en arreglo factorial 4x2x2, en el cual los factores correspondieron a dosis de GA3, régimen lumínico y temperatura.
La hormona se aplicó sumergiendo las semillas en las soluciones de cada tratamiento durante 24 h, con secado posterior al ambiente. Luego se colocaron en cajas Petri con papel filtro humedecido con agua destilada, las cuales se mantuvieron en cámaras de germinación con las condiciones de luz y temperatura mencionadas, para evaluar el porcentaje de germinación durante 30 d.
Con los datos de porcentaje de germinación, con base en la semilla viable, transformados a arcosenos, procedimiento recomendado cuando estos presentan dispersión amplia (Little y Hills, 1976), se hizo análisis de varianza y con la significación obtenida, se aplicó la prueba de partición de promedios de Duncan con una probabilidad de error Pâ¤0,05, mediante el empleo del programa SAS, versión 9.1.
Resultados y discusión
Curvas de secado de las semillas
La pérdida de humedad de las semillas expuestas a sílica gel en las cámaras herméticas se ajustó a modelos cuadráticos altamente significativos (P<0,0001) (Fig. 1), cuyas ecuaciones fueron: Y= 0,0088x2 - 0,6956x + 17,79; Y= 0,0108x2 - 0,8151x + 18,156 y Y= 0,0119x2 - 0,8694x + 18,106, para los tratamientos 1:1, 1:2 y 1:3, respectivamente. En el estudio se encontró un CHE promedio de 3,2%, a partir de un contenido inicial de 21,97%, en un tiempo medio de 14 h y 19 min. El tratamiento de semilla: sílica gel 1:3 presentó una desecación más rápida hasta llegar a peso constante, debido a la mayor superficie de absorción de agua.
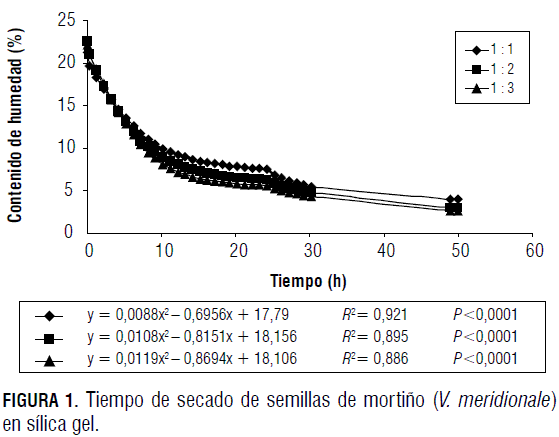
Las curvas logradas son útiles para predecir el tiempo requerido para la obtención de una humedad determinada, a partir del contenido inicial de esta, según el procedimiento descrito por Rao et al. (2006). Los tres tratamientos permitieron alcanzar la humedad requerida para los ensayos de la investigación, 5%. Se seleccionó para los estudios posteriores el tratamiento 1:1, ya que con este la pérdida de humedad es más lenta, lo cual disminuye riesgos de daño de los embriones.
Protocolo de tinción con tetrazolio
La metodología más adecuada para la evaluación de viabilidad de las semillas fue la extracción de los embriones. Esta se realizó separando una pequeña porción de la testa, retirando el embrión por medio de presión manual. Las otras metodologías causaron daño a este.
Como puede apreciarse en las Fig. 2A y Fig. 2B, se obtuvo una mejor tinción con la dosis de 1% de tetrazolio en comparación con la de 0,5%, con resultados más evidentes con imbibición en el químico, al 1%, durante 3 h a 30°C y en condiciones de temperatura ambiente por espacio de 24 h. La prueba de tetrazolio es un método reconocido para el análisis de viabilidad de las semillas (Nurse y Di Tommaso, 2005). Este permite categorizar una semilla como viable o no viable a partir de la tinción de los tejidos del embrión (Borza et al., 2007).

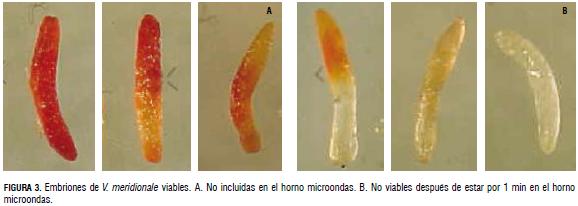
Las semillas prehumedecidas, sometidas a microondas durante un min, perdieron completamente la viabilidad, con germinación nula. Esto permitió caracterizar los patrones de tinción diferentes, con la aplicación del tetrazolio (1% durante 24 h a temperatura ambiente) a los obtenidos con el lote con semillas viables. En las Fig. 3A y Fig. 3B, se incluyen las tinciones obtenidas en las simientes tratadas y no tratadas con microondas. Como puede apreciarse, los embriones de las no tratadas, exhibieron coloración en la mayoría de su superficie, desde rosado claro, hasta rosado fuerte, con presencia también de tonalidades anaranjadas. Los patrones anteriores no fueron evidentes en las sometidas a microondas. Estas últimas, exhibieron unidades transparentes o coloración leve.
Categorización de la germinación
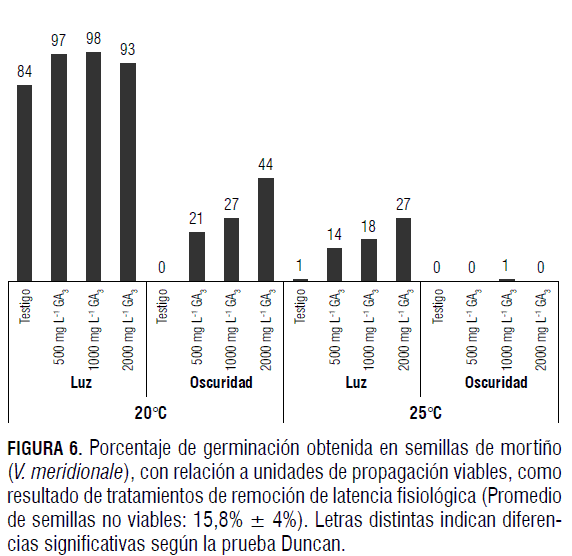
En la Fig. 4, se incluye la categorización de la viabilidad (germinación y latencia) y de la no viabilidad (ausencia de tinción con tetrazolio) obtenida en las semillas de mortiño recién extraídas del fruto, germinadas en cajas Petri a 21°C, en condiciones de luz-oscuridad de 12-12 h, respectivamente.
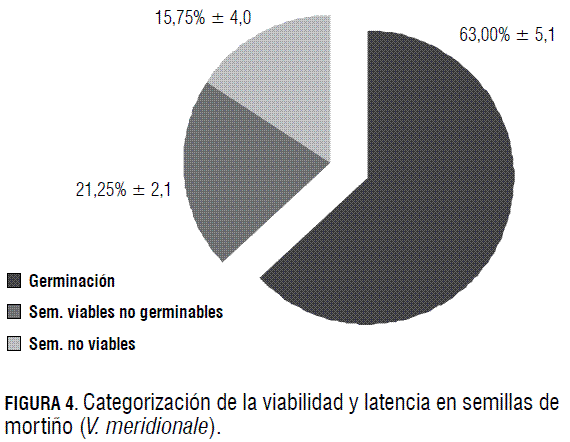
Como se aprecia en la Fig. 4, las semillas exhibieron 84,2% de viabilidad, con 63% de semillas germinadas y 21,2% de semillas viables no germinables (latentes). Además se encontró, 15,8% de semillas no viables, con base en la prueba de tetrazolio, en las no germinadas.
Lo anterior señala presencia de latencia, aspecto típico de las plantas silvestres. Esta permite responder a condiciones ambientales variables, constituyendo una estrategia de dispersión en el tiempo de la germinación (Snyder, 2006). Fenner y Thompson (2005) indicaron que el momento de la germinación es esencial para el ciclo de vida del vegetal, asegurando su ocurrencia en el periodo más favorable para el establecimiento de las plántulas.
Vallejo (2000) obtuvo una germinación de 82,7% con semillas de mortiño, valor superior al de la investigación actual. Esto puede deberse al hecho de que el autor condujo la evaluación por más de 30 d, tiempo máximo para la categorización de la germinación de semillas no latentes (Baskin y Baskin, 2001). Por ello la cifra agrupa semillas no latentes y latentes, con un valor cercano de viabilidad al alcanzado en la investigación actual para las dos categorías, 84,2%.
Categorización de la latencia
Latencia exógena
Como puede observarse en la Fig. 5, durante las 8 h en que estuvieron las semillas en contacto con agua, se presentó imbibición, con un incremento en peso cercano al 7%.
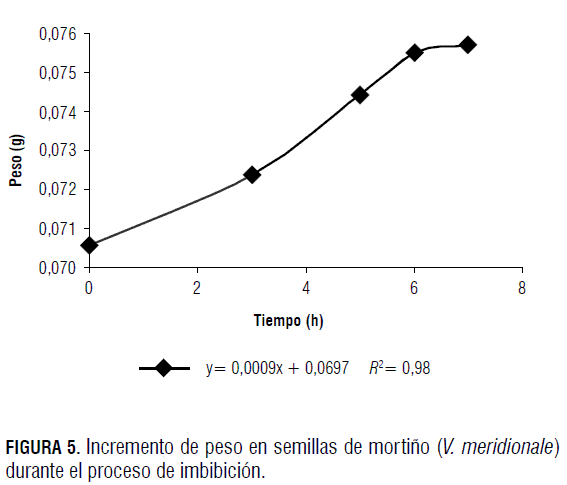
Lo anterior indica permeabilidad en la testa, lo que permite concluir que las semillas de mortiño no presentan latencia exógena o física, de acuerdo con el criterio de Nikolaeva (1977). No se ubicó reporte alguno de este tipo de latencia en estudios realizados con diversos Vaccinium.
Respuesta fotoblástica
Como puede verse en la Fig. 6, las semillas de mortiño son fotoblásticamente positivas, categorización dada a aquellas cuya germinación es estimulada por la luz (Sawada et al., 2008). Esto se colige al comparar la germinación, bajo regímenes de luz y oscuridad en los niveles de temperatura evaluados. Así, en condiciones de luz, sin aplicación de GA3, esta estimuló la germinación con un efecto más marcado, significativamente superior a 20°C que el obtenido a 25°C, 84% y 1%, respectivamente. En contraste, en las semillas sin tratamiento con hormona no hubo emergencia alguna del embrión en la oscuridad. Shahram (2007) reportó un efecto marcado en la germinación de V. arctostaphylos por parte de condiciones alternas de luz y oscuridad, la cual no ocurrió en ausencia de la primera.
En otros Vaccinium se ha informado sobre porcentajes de germinación de alrededor de 90%, en condiciones de luz del espectro rojo (Giba et al., 1995). Este promueve la germinación de las semillas que se encuentran superficiales en el suelo del bosque, cuando incide la luz. Se ha indicado que las especies con simientes pequeñas, como el mortiño, tienen requerimientos definidos y estrechos para su establecimiento por recursos internos limitados, empleando la luz como un indicador de condiciones adecuadas para la germinación (Pearson et al., 2003).
Latencia endógena
Como se puede apreciar en la Fig. 6, fue evidente el efecto de la temperatura sobre la germinación, con una emergencia significativamente superior a 20°C, en comparación con la obtenida a 25°C. Así, el nivel máximo de germinación de semillas viables a 25°C fue de 27%, en tanto que a 20°C este alcanzó valores de 97, 98 y 93% al tratar las simientes con GA3, en condiciones de luz, sin diferencias significativas entre estos valores con concentraciones de 500, 1.000 y 2.000 mg L-1, respectivamente. Igualmente, bajo iluminación, se obtuvo germinación, del 84%, sin aplicación de GA3, a 20°C. En tanto que esta fue de 1% a 25°C, con valores superiores del primer tratamiento a los obtenidos en todos los casos de aplicación de hormona a 25°C, en los cuales se obtuvo una emergencia de embriones del orden de 14, 18 y 27%, para las concentraciones de 500, 1.000 y 2.000 mg L-1 de GA3 (Fig. 6). La respuesta a la temperatura debe corresponder a las condiciones ambientales del hábitat del mortiño, la parte Altoandina (Luteyn, 2002), zona de temperaturas bajas. La germinación es un caso de regulación del crecimiento que comprende interacciones entre la temperatura y la actividad de la giberelina, con reportes de que esta regula la actividad de los genes relacionados con el metabolismo del GA (Penfield, 2008). Igualmente, se han postulado relaciones entre la temperatura y la iluminación para la germinación, con promoción de la biosíntesis del GA (Penfield et al., 2005).
El efecto positivo del GA en la germinación de las semillas de V. meridionale -lo cual también fue reportado en Colombia por Magnitskiy y Ligarreto (2007)- indica que estas, además de ser fotoblásticamente positivas, tienen latencia fisiológica, no profunda, de acuerdo con la clasificación de Nikolaeva (Baskin y Baskin 2001; 2008), Esta se removió a través de la imbibición con GA3, en las concentraciones evaluadas de la hormona, sin diferencias significativas entre estas, incubando las semillas a 20°C, en condiciones de luz blanca (Fig. 6).
Igualmente, se obtuvo una mayor velocidad de germinación, referida al tiempo tomado para la emergencia de 50% de los embriones (Fig. 6). Así, las semillas tratadas con 1.000 y 2.000 mg L-1 de GA3, a 20°C, tomaron en promedio 12 d para esto, y aquellas que fueron sometidas a 500 mg L-1, emplearon 14 d; en contraste, las que no recibieron la hormona, en condiciones de luz y a 20°C, tardaron 19 d para la emergencia de 50% de los embriones, en las semillas viables (Fig. 7A). En la Fig. 7B, se aprecia que las semillas tratadas con GA3, bajo oscuridad, al igual que las sin tratamiento, no alcanzaron el 50% de germinación de las semillas viables.
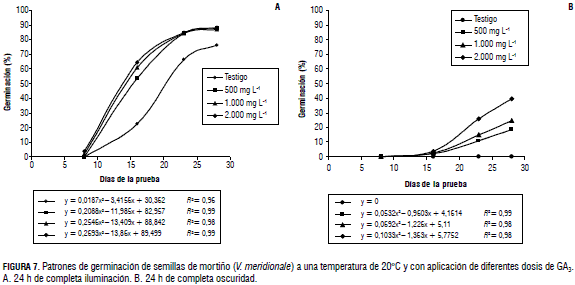
El efecto promotor del GA en la velocidad de germinación corresponde a la acción complementaria de la luz y la hormona. Se ha señalado que ambos están relacionados con un patrón que involucra el fitocromo, mediante una acción antagónica a la acción del ácido abscísico (Leubner-Metzger, 2001). Ballington et al. (1976) afirmaron que la aplicación de giberelinas a las simientes de Vaccinium, más que un incremento en la germinación, producen mayor velocidad del proceso, y Finkelstein et al. (2008) señalaron que la remoción de la latencia es regulada por una combinación de señales ambientales y endógenas, con sinergismo y competencia entre estas, lo cual coincide con afirmaciones de que al aplicar GA a las semillas del género Vaccinium, no se incrementa el porcentaje total de germinación, sino que aumenta su velocidad y la uniformidad del proceso, disminuyendo la necesidad de horas de luz (Giba et al., 1995). Esto no concuerda con lo obtenido en la investigación actual, en la cual el GA indujo incremento en la velocidad de germinación y mayor emergencia de embriones, en comparación con la simiente sometida únicamente a luz continua. Kucera y Leubner-Metzger (2005) indicaron que el GA incrementa el potencial de crecimiento del embrión y ayuda a vencer la resistencia mecánica de los tejidos que rodean la radícula, lo cual facilita la germinación.
Fenner y Thompson (2005) señalaron que la latencia no debe asociarse únicamente con la ausencia de germinación, y que es una característica de la semilla que determina las condiciones requeridas para la germinación. Finch-Savage y Leubner-Metzger (2006) indicaron que, cuando la latencia es considerada en la forma anterior, cualquier aspecto ambiental que altera las condiciones requeridas para la germinación también tiene efecto sobre la latencia. Benech-Arnold et al. (2000) afirmaron que la liberación de la latencia debe ser considerada como un proceso separado de la germinación, y Goggin et al. (2008) indicaron que esto es particularmente aparente cuando las condiciones requeridas para liberar la latencia son diferentes a las requeridas para la germinación. Por ello es importante evaluar la profundidad de la latencia tanto en tiempo como en espacio. Igualmente, se considera relevante evaluar la variabilidad intraespecífica de la germinación y la latencia. Estas varían ampliamente entre poblaciones de una misma especie y taxa del mismo género (Veasey et al., 2004).
Conclusiones
La extracción de embriones por presión manual permitió evaluar, en forma eficiente, la viabilidad de estos, con imbibición en una solución de tetrazolio de 1%, a 30°C, en condiciones de oscuridad por espacio de 24 h, a temperatura ambiente.
La aplicación de microondas a la semilla durante 1 min mató los embriones, lo que permitió diferenciar los patrones de tinción con tetrazolio entre simientes viables latentes y no viables.
Las simientes del mortiño no presentaron latencia exógena, con base en la permeabilidad de la testa al agua.
Las semillas de mortiño son fotoblásticamente positivas, esto es, su germinación es estimulada por la luz.
El mortiño exhibió semillas viables germinables, latentes y no viables, luego de una prueba de germinación a 21°C, por espacio de 30 d, con un régimen de 12:12 h de luz:oscuridad.
La temperatura influyó en la germinación, con valores sensiblemente superiores a 20°C, en comparación con la obtenida a 25°C, con un efecto complementario entre la luz y la temperatura de germinación.
La latencia de las simientes de mortiño corresponde a la categoría de fisiológica poco profunda. Esta se pudo remover a través de imbibición previa con GA3, en concentraciones de 500, 1.000 y 2.000 mg L-1, en incubación con luz blanca continua.
Agradecimientos
La investigación se realizó gracias al apoyo financiero de Colciencias, en el marco del proyecto "Zonificación de las especies de agraz (Vaccinium spp.) y una aproximación de su manejo agronómico como cultivos promisorios para la zona alto andina colombiana". código 1101-07-17186 de 2004.
Literatura citada
Ballington, J.R., G.J. Galleta y D.M. Pharr. 1976. Gibberellin effects on rabbiteye blueberry seed germination. HortScience 11(4), 410-411. [ Links ]
Bansal, R.P., P.R. Bathi y D.N. Sen. 1980. Differential specificity in water imbibitions of Indian arid zone seeds. Biol. Plant. 22, 327-331. [ Links ]
Baskin, C.C., P.L. Milberg, J.M. Andersson y J.M. Baskin. 2000. Germination studies of three dwarf shrubs (Vaccinium, Ericaceae) of Northern Hemisphere coniferous forests. Can. J. Bot. 78(12), 1552-1560. [ Links ]
Baskin, C.C. y J.M. Baskin. 2001. Seeds. ecology, biogeography, and evolution of dormancy and germination. Academic Press, San Diego, CA. [ Links ]
Baskin, J.M. y C.C. Baskin. 2008. Some considerations for adoption of Nikolaeva's formula system into seed dormancy classification. Seed Sci. Res. 18, 131-137. [ Links ]
Benech-Arnold, R.L., R.A. Sánchez, F. Forcella, B.C. Kruk y C.M. Ghersa. 2000. Environmental control of dormancy in weed seed banks in soil. Field Crops Res. 67, 105-122. [ Links ]
Borza, J.K., P.R. Westerman y M. Liebman. 2007. Comparing estimates of seed viability in three foxtail (Setaria) species using the imbibed seed crush test with and without additional tetrazolium testing. Weed Technol. 21, 518-522. [ Links ]
Castañeda, R.I. 2006. Conservación, caracterización y divulgación del conocimiento de poblaciones de mortiño (Vaccinium meridionale), presentes en los bosques altoandinos de la jurisdicción de Corantioquia. Trabajo de grado. Departamento de Ciencias Agronómicas, Universidad Nacional de Colombia, Medellín, Colombia. [ Links ]
Corporación Autónoma Regional del Centro de Antioquia (Corantioquia). 2003. Conozcamos y usemos el mortiño. Diseño e Impresos, Medellín. [ Links ]
Darrow, G.M. y D.H. Scott. 1954. Longevity of blueberry seed in cool storage. Proc. Amer. Soc. Hort. Sci. 63, 271. [ Links ]
Espinal, S. 1977. Zonas de vida: formaciones vegetales del departamento de Antioquia. Universidad Nacional de Colombia, Medellín. [ Links ]
FAO. 1996. Report on the state of the world's plant genetic resources for food and agriculture. FAO, Roma. [ Links ]
Fenner, M. y K. Thompson. 2005. The ecology of seeds. Cambridge University Press, Cambridge, UK. [ Links ]
Finch-Savage, W.E. y G. Leubner-Metzger. 2006. Seed dormancy and the control of germination. New Phytologist 171, 501-523. [ Links ]
Finkelstein, R., W. Reeves, T. Ariizumi y C. Steber. 2008. Molecular aspects of seed dormancy. Annu. Rev. Plant Biol. 59, 387-415. [ Links ]
Gentry, A.H. 1988. Changes in plant community diversity and floristic composition on environmental and geographic gradients. Ann. Missouri Bot. Garden. 75, 1-34 [ Links ]
Giba, Z., D. Grubisic y R. Konjevic. 1995. The involvement of phytochrome in light- induced germination of blueberry (Vaccinium myrtillus L.) seeds. Seed Sci. Technol. 23(1), 11-19. [ Links ]
Goggin, D.E., K.J. Steadman y S.B. Powles. 2008. Green and blue light photoreceptors are involved in maintenance of dormancy in imbibed annual ryegrass (Lolium rigidum) seeds. New Phytol. 180, 81-89. [ Links ]
Guarino, L., A. Jarvis, R.J. Hijmans y N. Maxted. 2001. Geographic Information Systems (GIS) and the conservation and use of plant genetic resources. pp. 387-404. En: Engels J. (ed.). Managing Plant Diversity. CAB International, Wallingford, UK. [ Links ]
Hammer, K., H. Heller y J. Engels. 2001. Monographs on underutilized and neglected crops. Genet. Resour. Crop Evol. 48, 3-5. [ Links ]
Hernández, M.I. 2008. Estudio morfo-fisiológico de la semilla del mortiño (Vaccinium meridionale Swartz). Tesis de maestría. Magíster en Ciencias Agrarias, Universidad Nacional de Colombia, Medellín. [ Links ]
Hill, N.M. y S.P. Vander. 2005. Longevity of experimentally buried seed in Vaccinium: relationship to climate, reproductive factors and natural seed banks. J. Ecol. 93, 1167-1176. [ Links ]
ISTA. 1999. Reglas internacionales para ensayos de semillas. Manual de ensayos al tetrazolio. Ministerio de Agricultura, Pesca y Alimentación, Madrid. [ Links ]
Jarvis, A., K. Williams, D. Williams, L. Guarino, P.J. Caballero y G. Mottrama. 2005. Use of GIS for optimizing a collecting mission for a rare wild pepper (Capsicum flexuosum Sendtn.) in Paraguay. Genet. Resour. Crop Evol. 52, 671-682. [ Links ]
Kucera B., M.A. Cohn y G. Leubner-Metzger. 2005. Plant hormone interactions during seed dormancy release and germination. Seed Sci. Res. 15, 281-307. [ Links ]
Leubner-Metzger, G. 2001. Brassinosteroids and gibberelins promote tobacco seed germination by different pathways. Planta 213, 758-763. [ Links ]
Little, T.M. y F.J. Hills. 1976. Métodos estadísticos para la investigación en la agricultura. Editorial Trillas, México. [ Links ]
Luteyn, J.L. 2002. Diversity, adaptation, and endemism in neotropical Ericaceae: Biogeographical patterns in the Vaccinieae. Bot. Rev. 68(1), 55-87. [ Links ]
Magnitskiy, S. y G. Ligarreto. 2007. El efecto del nitrato de potasio, del ácido giberélico y del ácido indolacético en la germinación de semillas de agraz (Vaccinium meridionale Swartz). Rev. Colomb. Cienc. Hortic. 1(2), 137-141. [ Links ]
McGlaughlin, M.E. y E.A. Friar. 2007. Clonality in the endangered Ambrosia pumila (Asteraceae) inferred from RAPD markers; implications for conservation and management. Conserv. Genet. 8, 319-330. [ Links ]
Medina, C.I., M. Vargas, M. Lobo y J.L. Toro, 2008. La propagación sexual del mortiño. Corpoica, Corantioquia. Medellín, Colombia. [ Links ]
Nikolaeva, M.G. 1977. Factors controlling the seed dormancy pattern in the physiology and biochemistry of seed dormancy and germination. Nort Holland, Amsterdam. pp. 51-74. [ Links ]
Nurse, R.E. y A. Di Tommaso. 2005. Corn competition alters germinability of velvetleaf (Abutilon theophrasti) seeds. Weed Sci. 53, 479-488. [ Links ]
Patiño, M. Del P. y G.A. Ligarreto. 2006. Caracterización morfológica in situ de poblaciones espontáneas de Vaccinium sp. en los departamentos de Boyac´, Cundinamarca y Nariño. p. 112. En: Fischer, G., D. Miranda, W. Piedrahita y S. Magnitskiy (eds.). Memorias Primer Congreso Colombiano de Horticultura, Unibiblos, Bogotá. [ Links ]
Pearson, T.R.H., D.F.R.P. Burslem, C.E. Mullins y J.W. Dalling. 2003. Functional significance of photoblastic germination in neotropical pioneer trees: a seed's eye view. Funct. Ecol. 17, 394-402. [ Links ]
Penfield, S. 2008. Temperature perception and signal transduction in plants. New Pathol. 179, 615-628. [ Links ]
Penfield, S., E.M. Josse, R. Kannagara, A.D. Gilday, K.J. Halliday e I.A. Graham. 2005. Cold and light control seed germination through the bHLH transcription factor SPATULA. Curr. Biol. 15, 1998-2006. [ Links ]
Poulsen, G., C. Holten y R. Von Bohmer. 2006. Identification and revival of low viability seed samples. Genet. Resour. Crop Evol. 53, 675-678. [ Links ]
Rao, N.K., J. Janson, M.E. Duloo, K. Ghosh, D. Nowell y M. Larinde. 2006. Manual of seed handling in genebanks. Handbook for genebanks No 8. Biodiversity International. ILRI, FAO, Roma. [ Links ]
Romero R. 1991. Frutas silvestres de Colombia. 2a ed. Instituto de Cultura Hispánica, Bogotá. [ Links ]
Ruggiero, M.V., T.B. Reusch y G. Procaccini. 2005. Local genetic structure in a clonal dioecious angiosperm. Mol. Ecol. 14, 957-967. [ Links ]
Sawada, Y., T. Katsumata, J. Kitamura, H.Ç. Kawalde, M. Nakajima, T. Asami, K. Nakaminami, T. Kurahashi, W. Misuhashi, Y. Inoue y T. Toyomasu. 2008. Germination of photoblastic lettuce seeds is regulated via the control of endogenous physiologically active gibberelin content, rather than of giberellin responsiveness. J. Exp. Bot. 59(12), 3383-3393. [ Links ]
Shahram, S. 2007. Seed dormancy and germination of Vaccinium arctostaphylos L. Intl. J. Bot. 3(3), 307-311. [ Links ]
Shoen, D.J. y A.H.D. Brown. 2001. The conservation of wild plant species in seed banks. Bioscience 51(11), 960-966. [ Links ]
Snyder, R.E. 2006. Multiple risk reduction mechanisms: can dormancy substitute for dispersal? Ecol. Lett. 9, 1106-1114. [ Links ]
Taruscio T.G., D.L. Barney y J. Exon. 2004. Content and profile of flavanoid and phenolic acid compounds in conjunction with the antioxidant capacity for a variety of Northwest Vaccinium berries. J. Agr. Food Chem. 52, 3169-3176. [ Links ]
USDA, Forest Service. s.f. Vaccinium L. blueberry or cranberry. En: Woody Plant Seed Manual, http://www.nsl.fs.fed.us/wpsm/Vaccinium.pdf; consulta: 17 de septiembre de 2008. [ Links ]
Vallejo, D.A. 2000. Fomento al mortiño (Vaccinium meridionale) como especie promisoria del Parque Regional Arví. Memoria Institucional. Corantioquia, Medellín, Colombia. [ Links ]
Veasey, E.A., M.G. Karasawa, P.P. Santos, M.S. Rosa, E. Mamani y G.C.X. Oliveira. 2004. Variation in the loss of seed dormancy during after-ripening of wild and cultivated rice species. Ann. Bot. 94, 875-882. [ Links ]
Victoria, J.A. 2005. Caracterización morfofisiológica de semillas de caléndula (Calendula officinalis L.) y eneldo (Anethum graveolens L.). Tesis de maestria. Universidad Nacional de Colombia, Palmira. [ Links ]

