










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.27 no.1 Bogotá Jan./Apr. 2009
Karen Muñoz1, Luz Fuentes2, Fernando Cantor1, 3, Daniel Rodríguez1 y José Ricardo Cure1
1 Programa de Biología Aplicada, Universidad Militar Nueva Granada, Bogotá (Colombia).
2 Centro de Investigaciones y Asesorías Agroindustriales (CIAA), Universidad de Bogotá Jorge Tadeo Lozano, Chía (Colombia).
3 Autor de correspondencia. ecologia@umng.edu.co
Fecha de recepción: 27 de octubre de 2008. Aceptado para publicación: 19 de febrero de 2009
RESUMEN
Se evaluó la preferencia de presas de Balaustium sp., enemigo natural de diferentes artrópodos plaga, y el cual es nativo de la Sabana de Bogotá. En unidades experimentales construidas con foliolos de plantas de rosa se colocaron independientemente individuos de Balaustium sp. y se registró el número de presas consumidas. De esta manera se determinó la preferencia de los tres estados móviles del ácaro depredador Balaustium sp. por diferentes edades de tres presas. Las especies y edades de las presas estudiadas fueron: huevos, ninfas y adultos de Trialeurodes vaporariorum, huevos, ninfas y adultos de Tetranychus urticae, y larvas de primer y segundo instar y adultos de Frankliniella occidentalis. Los estados menos desarrollados fueron preferidos, aunque se observó que los adultos del depredador tienen gran habilidad para consumir adultos de T. vaporariorum. La presa preferida por las larvas de Balaustium sp. fue los huevos de T. urticae con una proporción de consumo de 0,54 de los huevos que se ofrecieron de esta presa; las deutoninfas del depredador eligieron huevos de T. vaporariorum (0,537) o de T. urticae (0,497) y los adultos de Balaustium sp. prefirieron los huevos de T. vaporariorum (0,588).
Palabras clave: enemigo natural, control biológico de plagas, Trialeurodes vaporariorum, Frankliniella occidentalis, Tetranychus urticae.
ABSTRACT
The mite Balaustium sp. is native of the Bogotá Plateau, and a natural enemy of different pests arthropods. With the aim of evaluating its prey preferences we placed separate individuals of its three motile stages in experimental units containing rose leaf lets and different prey samples. Prey consumption was recorded for eggs, nymphs and adults of Trialeurodes vaporariorum and Tetranychus urticae, and for first and second instar larvae and adults of Frankliniella occidentalis. Although the mites fed mostly on initial stages of their prey, adults of Trialeurodes vaporariorum were seen to be easily preyed by adult predators. The larvae consumed 54% of the offered eggs of Tetranychus urticae, which were their preferred prey. The deutonymphs chose similar proportions of Trialeurodes vaporariorum (53.87%) and Tetranychus urticae (49.7%) eggs. Adult predators fed more abundantly on eggs of Trialeurodes vaporariorum (58.8%).
Key words: natural enemy, biological control of pests, Trialeurodes vaporariorum, Frankliniella occidentalis, Tetranychus urticae.
Introducción
Tetranychus urticae (Acari: Tetranychidae), Frankliniella occidentalis (Thysanoptera: Thripidae) y Trialeurodes vaporariorum (Hemiptera: Aleyrodidae) constituyen algunas de las plagas de mayor impacto económico en cultivos de flores de corte (Barrera et al., 1997; Mendoza et al., 2001).
En el mercado internacional de flores, principalmente en Europa, existen exigencias para comercializar productos más limpios, en cuyo proceso de producción se minimice el uso de químicos para el control de plagas, debido a los efectos negativos que su empleo genera en el medio ambiente y en la salud humana. Además la utilización continua de acaricidas selecciona poblaciones resistentes de los fitófagos y disminuye la eficacia del control por esta técnica. Por estas razones se considera el uso de otras alternativas de control de plagas, como el control etológico, el uso de extractos de plantas y el control biológico con el uso de enemigos naturales (Cardona et al., 2001; Bielza et al., 2007; Flores et al., 2007; Pallini et al., 1997; Venzon et al., 2001).
En Europa, las especies de artrópodos depredadores nativos son de gran importancia en programas de manejo integrado de plagas, principalmente por su adaptación a condiciones ambientales variables, lo cual les confiere mayor posibilidad de permanecer en el cultivo y de controlar las plagas de una manera muy eficiente (Symondson et al., 2002). Especies del género Balaustium (Acari: Erythraeidae) son consideradas como depredadores generalistas con potencial como controladores biológicos (Cadogan y Laing, 1977; Welbourn y Jennings, 1991; Yoder et al., 2006).
Balaustium es un género cosmopolita de la familia Erythraeidae (Acarina) (Hayes, 1985) subfamilia Balaustiinae (Haitlinger, 2005). Esta familia a su vez pertenece a un grupo denominado Parasitengona terrestre. En Norteamérica se consideran para propósito de estudio como controladores biológicos de Tetranychus urticae (Yoder et al., 2006).
Hayes (1985) reportó que las especies del género Balaustium consumen huevos y larvas de lepidópteros, dípteros y áfidos adultos. También pueden usar otros recursos alimenticios como polen. B. muororum es una especie que ha sido considerada como depredadora benéfica en el sur de Australia. B. putmani pasa por siete estados de desarrollo: huevo, prelarva o deutovum, larva, protoninfa, deutoninfa, tritoninfa y adulto. Los estados móviles son larva, deutoninfa y adulto (Putman, 1970).
En Colombia, Torrado et al. (2001) y Getiva y Acosta (2004) reportaron la presencia de una especie del género Balaustium en cultivos de flores. Sin embargo, no hay estudios sobre su biología y ecología que permitan caracterizarla como controlador biológico.
Los estudios de preferencia de depredadores son importantes para tomar decisiones en el momento de su liberación para el control de plagas en campo. Además, Venzon et al. (2002) reportaron que los depredadores generalistas muestran preferencias marcadas hacia algunas presas. El fenómeno de preferencia se puede dar debido a dos factores: selección activa por el depredador y selección pasiva por la presa. Estos dos factores interactúan e influyen en la preferencia (Blais y Maly, 1993).
La elección activa consiste en que el depredador discrimina entre presas de diferente calidad (Lang y Gsödl, 2001). En este caso, los aspectos que pueden influir en la depredación son: la variación de tamaño entre presas (Peckarsky, 1980) y la calidad nutricional que suministran las diferentes presas al depredador (Mendes et al., 2002). Esta elección activa depende en buena parte de la interacción de las plantas con los enemigos naturales de los fitófagos, pues las plantas pueden atraer enemigos naturales a través de la producción de sustancias volátiles inducidas por la presencia de los fitófagos; también debido al suministro de refugio o comida para estos enemigos naturales de plagas (Venzon et al., 2001). Los químicos volátiles de origen vegetal desempeñan un valioso papel durante la búsqueda de presas por parte de artrópodos carnívoros (Gnanvossou et al., 2002). Estos pueden variar dependiendo de la especie de planta y del fitófago, de la variedad de planta y de su grado de infestación (Venzon et al., 2001, Gnanvossou et al., 2002).
Por otro lado, la selección pasiva se basa en las diferencias intrínsecas entre los tipos de presas en respuesta al ataque del depredador (Reitz et al., 2006), principalmente aspectos relacionados con la capacidad de las presas para evadir al depredador (Lang y Gsödl, 2001). Para diseñar experimentos de preferencia de depredadores es necesario tener en cuenta el aprendizaje en artrópodos, por ejemplo Boer et al., (2005) reportaron que por medio de este fenómeno los ácaros depredadores pueden encontrar sus presas, especialmente durante el aprendizaje preimaginal que consiste en el comportamiento que adoptan los artrópodos que es dependiente de experiencias olfatorias de los estados inmaduros y que influye en su preferencia al buscar comida, pareja y sitios de oviposición (Gutiérrez, 2007).
Los especímenes de Balaustium sp. utilizados en este estudio se enviaron al Dr. Cal Welbourn (Florida Dept. Agriculture & Consumer Services, Gainesville, FL, USA), especialista de la familia Erythraeidae. Según primera comunicación del Dr. Welbourn durante el desarrollo del presente estudio, se informó que se trata de una nueva especie. De esta manera, se evaluó la preferencia alimenticia de Balaustium sp. por diferentes especies de plagas y por edades de estas.
Materiales y métodos
Los experimentos y las crías de los fitófagos se mantuvieron en los cuartos de ambiente controlado del Centro de Investigaciones y Asesorías Agroindustriales (CIAA-UJTL), con una temperatura promedio de 22,1±2,0°C, humedad relativa de 75% y fotoperiodo de 12 h luz: 12 h oscuridad. El pie de cría del depredador se mantuvo bajo invernadero a una temperatura máxima promedio de 31,7±3,5°C y mínima promedio de 11,1±2,0°C, en recipientes plásticos cilíndricos de 18 cm de diámetro con una circunferencia de malla de acero de 10 cm de diámetro en la tapa para ventilación y turba en el fondo para mantener la humedad. Se introdujeron 15 individuos adultos a los cuales se les suministró como alimento las tres presas dos veces a la semana.
Elaboración de unidades experimentales
Se usaron foliolos de rosa o tomate con el peciolo dentro de un tubo de plástico con agua y con tapa de caucho para mantener la humedad del tejido. El foliolo se infestó con individuos de la plaga por evaluar y se colocó sobre la base de una caja de Petri de 4 cm de diámetro, cubierta con papel toalla, y sobre el foliolo se colocó un segmento de tubo PVC (altura 1 cm, diámetro 2 cm), para confinar los individuos. El montaje fue cubierto con plástico trasparente (vinipel) con el fin de evitar que los individuos escaparan del montaje.
Preferencia por edades de la presa
Se manejaron 20 repeticiones por cada tratamiento. Cada tratamiento correspondió a 10 individuos de cada edad de la presa (en el caso de mosca blanca y T. urticae se dispusieron adultos machos para que no se alterara la densidad de las presas) y un individuo de cada edad del depredador (Tab. 1).
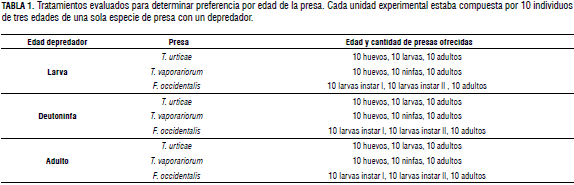
Preferencia por especies de presa
Se escogieron los estados que presentaron el mayor promedio de consumo en el experimento de preferencia por edades. De esta manera se realizaron 20 repeticiones de cada tratamiento. Cada uno de estos tratamientos estaba constituido por individuos de diferentes edades de las tres especies de presa (Tab. 2) y un depredador.
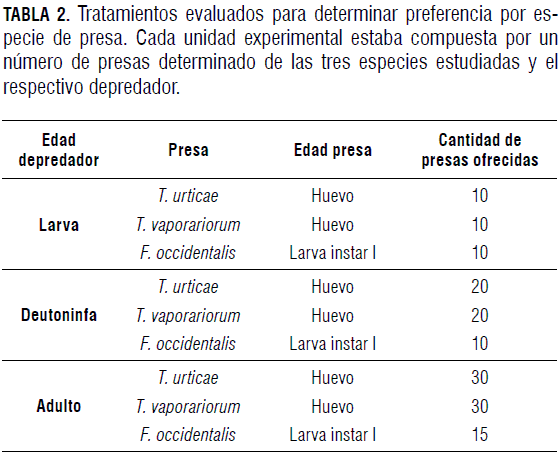
Cada individuo Balaustium sp. que se utilizó en los experimentos de preferencia fue mantenido en ayuno con anterioridad por 24 h. También se evaluó un testigo que consistió en los individuos presa en cada unidad experimental sin el depredador.
Análisis de datos
Al evaluar la preferencia por diferentes estados de las presas y diferentes especies de presa, el consumo se comparó mediante la prueba de Kruskal-Wallis debido a que en algunos casos no se presentó normalidad según la prueba de Shapiro-Wilk. En este caso, las comparaciones múltiples se efectuaron mediante la prueba de Behrens-Fisher. Para datos normales se empleó Anova y pruebas de comparación-múltiple de Tukey. Se usó el software R 2.5.0 para el análisis estadístico.
Resultados y discusión
Preferencia de Balaustium sp. por edad de la presa
Las larvas de Balaustium sp. prefirieron consumir estados con menor desarrollo de T. urticae, de F. occidentalis y de T. vaporariorum (Fig. 1). La preferencia fue significativamente mayor (Tab. 3) hacia huevos de T. vaporariorum, huevos de T. urticae y larvas de primer instar de F. occidentalis.
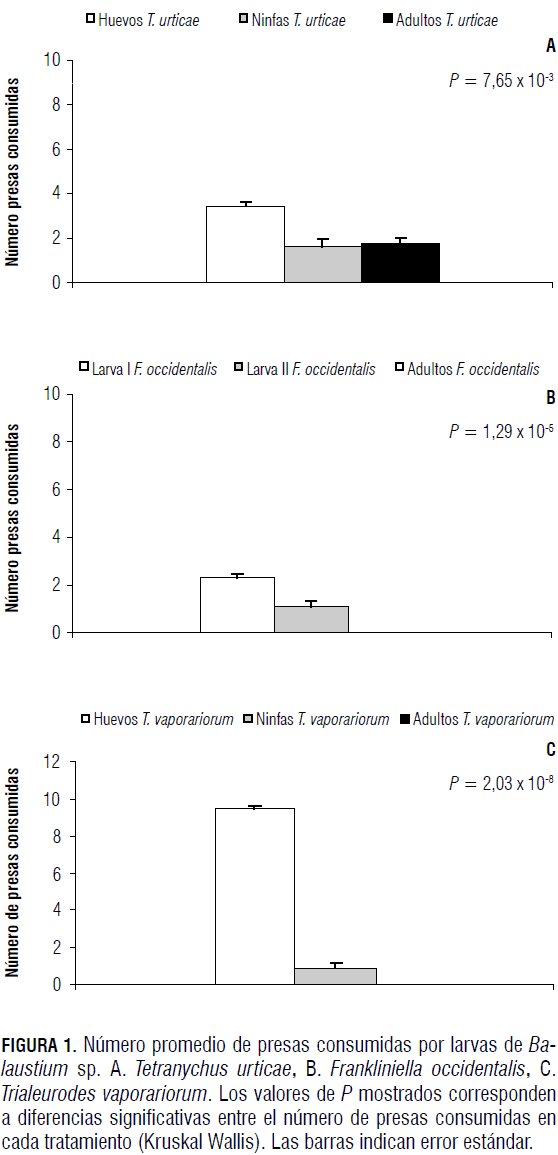
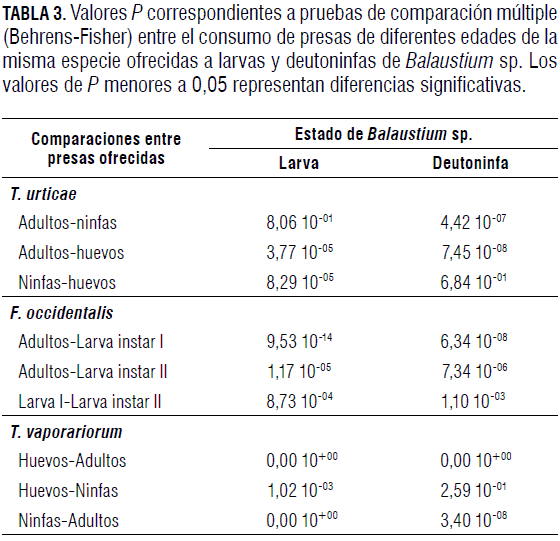
Al comparar la preferencia de presas de diferentes edades de T. urticae, F. occidentalis y T. vaporariorum por parte de deutoninfas Balaustium sp., se observó que estas son capaces de capturar no sólo individuos inmaduros sino también aquellos de edades más avanzadas. Las deutoninfas de Balaustium sp. prefirieron huevos (Fig. 2) y ninfas de T. urticae y de T. vaporariorum. Para el caso de larvas de F. occidentalis de diferentes edades, prefirieron las larvas de primer instar respecto a los individuos de otras edades ofrecidas (Tab. 3A).
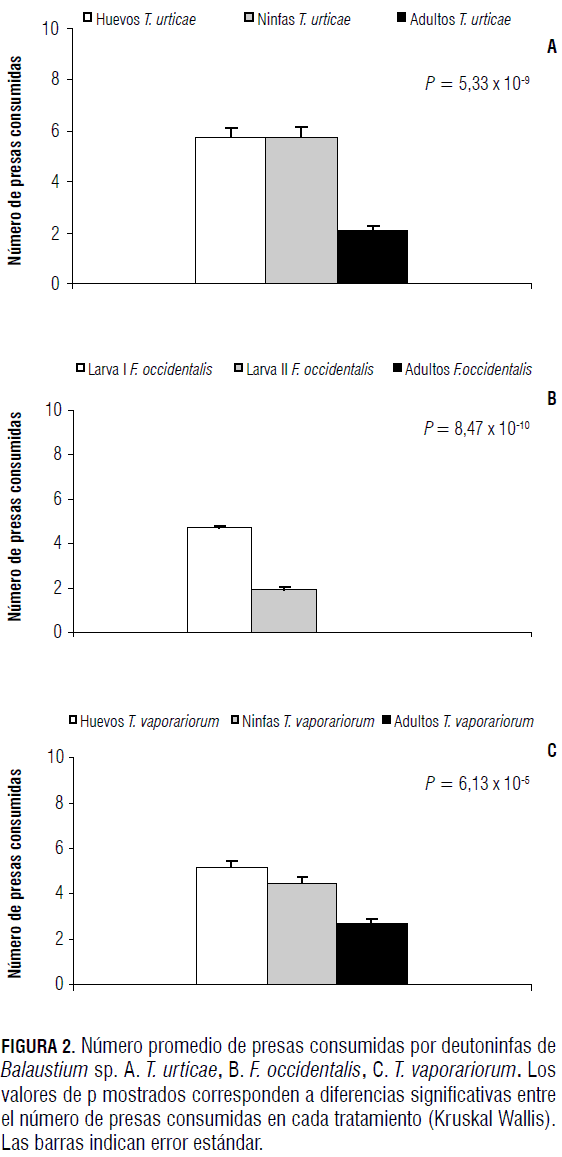
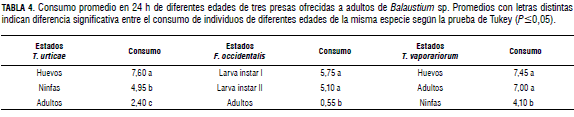
Así como en los casos anteriores, con relación a las larvas y deutoninfas, los adultos de Balaustium sp. presentaron preferencias por los estados con menor desarrollo que por los más avanzados. Sin embargo, a diferencia de las larvas y deutoninfas, los adultos consumieron individuos de todas las edades de la presa (Fig. 3). Cuando se les ofrecieron individuos de diferentes edades de T. urticae, prefirieron huevos, seguido de ninfas y finalmente adultos (Fig. 3A y Tab. 4). Esta misma tendencia se observó con F. occidentalis, caso en el cual los adultos prefirieron larvas de primer instar, seguido de larvas de segundo instar y finalmente adultos del fitófago (Fig. 3B y Tab. 4). Cuando se les ofreció a adultos del depredador diferentes edades de T. vaporariorum, presentaron mayores preferencias por huevos (Tab. 4) y adultos, que por ninfas del fitófago (Fig. 3C).
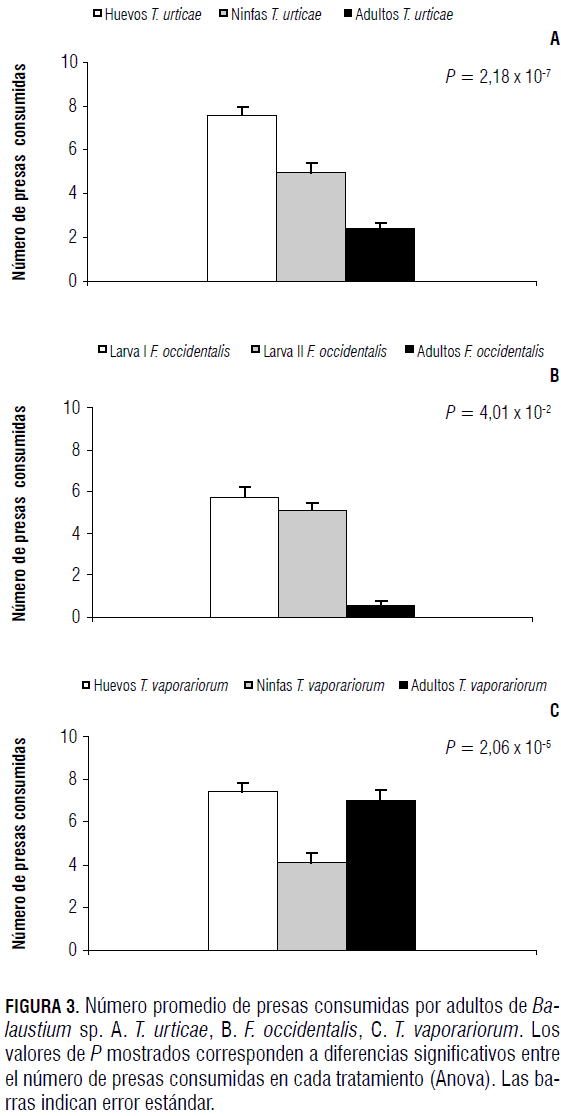
La tendencia a seleccionar estados menos desarrollados también ha sido evidenciada en otros estudios de preferencia de la presa T. urticae, como lo que reportaron Badii et al. (2004), quienes determinaron que el ácaro depredador Euseius hibisci prefiere huevos de T. urticae, en comparación con los otros estados. En ácaros de la familia Phytoseiidae se ha visto que los huevos de T. urticae son preferidos en comparación con otras edades de la presa (Oliveira et al., 2007). Así mismo, otros autores evidenciaron que los estados inmaduros de T. vaporariorum son preferidos por la chinche depredadora Macrolophus caliginosus (Heteroptera: Miridae) (Salem, 2006), y que larvas de F. occidentalis son preferidos por ácaros fitoseidos y Orius insidiosus (Hemiptera: Anthocoridae) en lugar de los adultos de esas mismas presas (Rodríguez et al., 1992; Reitz et al., 2006).
Estos resultados pueden ser atribuidos a aspectos que se relacionan con características nutricionales de la presa, tal como lo han sugerido Mendes et al. (2002) y Collier et al. (2007). Según Carrel y Tanner (2002), el alto contenido de proteína y de carbohidratos que hay en los huevos de los artrópodos influye en la maduración de huevos y en la fecundidad de adultos de sus depredadores, así como en el suministro de energía para la actividad de estados móviles de los mismos depredadores. Este fenómeno pudo ocurrir en la preferencia observada en el presente trabajo de los adultos de Balaustium sp. hacia los huevos de las presas ofrecidas. Por otra parte, para el caso de individuos del género Balaustium, en la literatura se registra que los individuos pertenecientes a este género no pueden aprovechar el agua contenida en la humedad ambiental, como ocurre en el caso de la gran mayoría de ácaros, razón por la cual buscan el agua líquida directamente o la que se encuentre contenida en sus presas (Yoder et al., 2006). Este aspecto pudo influir notablemente en la preferencia durante cualquiera de sus edades móviles.
Otros autores demostraron que además del aporte nutricional que les ofrecen las presas a los depredadores, las preferencias de consumo también se deben a la facilidad de captura de las mismas (Lang y Gsödl, 2001; Reitz et al., 2006). Así se podría afirmar que otra razón por la cual Balaustium sp. prefirió los huevos de la presa consiste en el factor de selección pasiva, ya que según Collier et al. (2007), los huevos de artrópodos que actúan como presas representan un recurso relativamente fácil de adquirir para sus depredadores pues el tiempo de manipulación de huevos es menor que el de presas más desarrolladas. Por el contrario, depredadores evitan presas de edades más avanzadas porque estas presentan diferentes estrategias de defensa que las hace difíciles de capturar (Choh y Takabayashi, 2006). Esta puede ser una razón por la cual larvas y adultos de Balaustium sp. presentaron menor capacidad de captura de ninfas y adultos de T. urticae, ya que se evidenció la huida de estas presas como posible mecanismo de defensa ante la presencia del depredador. De la misma manera, la baja capacidad de captura de Balaustium sp. hacia los adultos de F. occidentalis puede ser debido a factores de defensa manifestadas por la presa, pues adultos de trips utilizaban sus alas para escapar más ágilmente del depredador.
Además de la influencia de los factores nutricionales y de las estrategias de defensa en la preferencia, en el presente estudio se pudo observar que el tamaño de la presa también influye en el momento de elegir entre varias fuentes de alimento, como se registró por otros autores (Peckarsky, 1980; Roger et al., 2000; Reitz et al.,2006; Ferkovich et al., 2007). Se observó que las deutoninfas tienen más agilidad de movimiento que los individuos de los otros estados móviles del depredador; de esta manera se pone en ventaja la deutoninfa al momento de cazar las ninfas de T. urticae.
Las larvas y deutoninfas de Balaustium sp. prefirieron las larvas de primer instar de F. occidentalis debido a su menor tamaño en comparación con las larvas de segundo instar. Pese a que estudios de Nomikou et al. (2002) reportan que adultos de Bemisia tabaci son muy poco vulnerables a la actividad de ácaros depredadores de la familia Phytoseiidae, en comparación con los huevos u otros estados sésiles de la misma plaga, en este estudio se evidenció que debido a su tamaño y agilidad, Balaustium sp. también puede llegar a elegir como presa a los adultos de T. vaporariorum (Fig. 3C).
Preferencia de Balaustium sp. por especie de la presa
Al ofrecerle a Balaustium sp. tres tipos de presas en sus edades preferidas evaluadas anteriormente, tanto larvas como deutoninfas y adultos del depredador prefirieron huevos de T. urticae y de T. vaporariorum (Fig. 4 y Tab. 5).
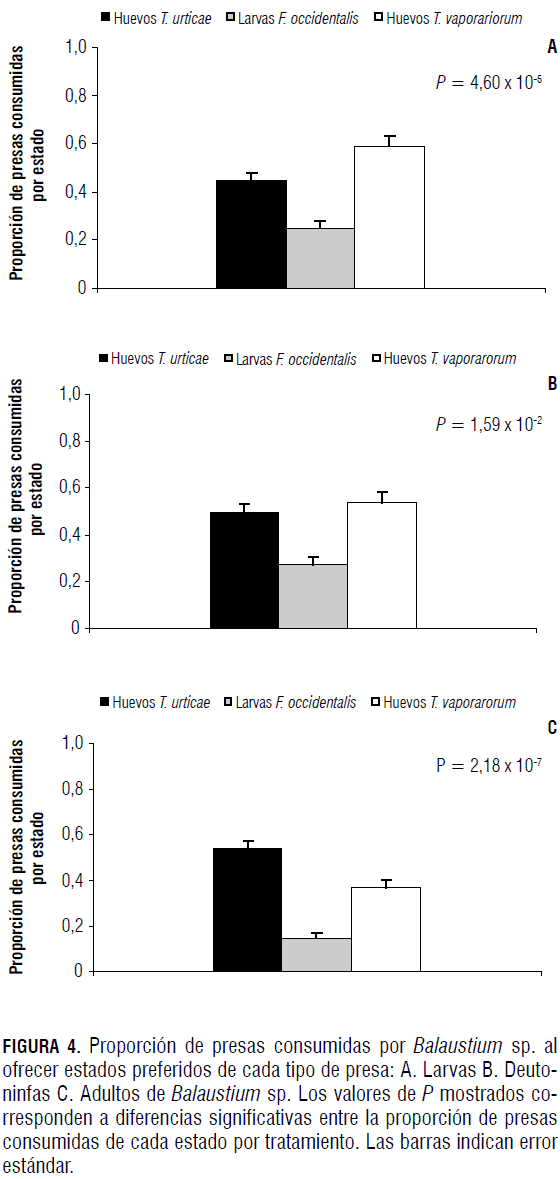
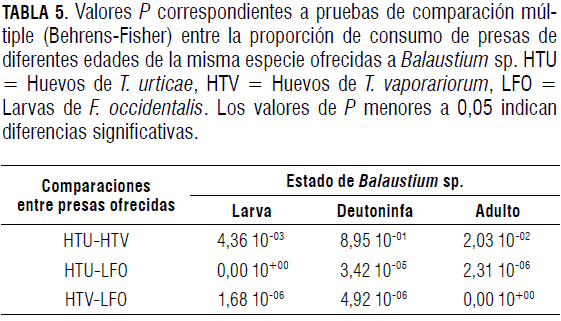
La preferencia de las larvas de Balaustium sp. por huevos de T. urticae observada en la Fig. 4A está acorde con los reportes hechos por Cadogan y Laing (1977) y Putman (1970), en los cuales las larvas de Balaustium sp. fueron observadas alimentándose principalmente de individuos del género Tetranychus. Aparentemente los huevos de T. urticae le proporcionaron a la larva del depredador los nutrientes esenciales para pasar al estado quiescente. Este tipo de selección se estudió para otros casos de ácaros depredadores, pues en otros estudios comprobaron que el contenido de carotenoides en las presas del género Tetranychus es importante para inducir diapausa (Overmeer et al., 1989; Dicke et al., 1990 y Veerman, 1992; Gnanvossou et al., 2003), lo cual podría sugerir que las larvas de Balaustium sp. pueden buscar este recurso al preferir T. urticae en comparación con F. occidentalis o T. vaporariorum.
Los adultos de Balaustium sp. prefirieron huevos de mosca blanca (Fig. 4), probablemente porque se encuentran asociados a la presencia de gotas de miel de rocío (Honey dew) producida por adultos de T. vaporariorum. Según Burger et al. (2005), la miel de rocío de moscas blancas contiene cantidades significativas de aminoácidos.
Las larvas de F. occidentalis fueron la presa menos consumida por Balaustium sp. (Fig. 4), pues se observó que para Balaustium sp. constituye una mayor dificultad la manipulación de larvas de trips en comparación con las otras presas debido a su comportamiento agresivo de defensa. Esto también lo reportaron Rodríguez et al. (1992) y De Bruijn et al. (2006), quienes evidenciaron que después de un periodo de búsqueda activa por parte de fitoseidos depredadores de F. occidentalis, estos contactan las larvas de trips por medio del primer par de patas o los palpos, sujetándolas con los quelíceros. Así mismo, Rodríguez et al. (1992) y De Brujin et al. (2006) evidenciaron que las larvas de trips golpean con el abdomen al depredador y producen pequeñas gotas fecales que en ocasiones obligan al depredador a retirarse para limpiarse.
Según Halliday (2001), Balaustium medicagoense puede ser una plaga en cultivos de cereales. En el presente trabajo se observó que cuando los individuos de Balaustium sp son sometidos a ayuno en foliolos de rosa y tomate sin presas, no se alimentan y después de algunos días mueren, por lo que no se encontró evidencia de que puedan representar algún riesgo potencial para los cultivos.
Conclusiones
Balaustium sp. depreda diferentes estados de Tetranychus urticae, Trialeurodes vaporariorum y Frankliniella occidentalis que se reportan como plagas de diferentes cultivos de importancia económica.
Balaustium sp. presenta preferencia hacia los estados menos desarrollados de los tres tipos de presas ofrecidos. Además hay diferencias en preferencia de alimentación según la edad de Balaustium sp.
Literatura citada
Badii, M.H., E. Herna, N. Ortiz, A.E. Flores y J. Landeros. 2004. Prey stage preference and functional response of Euseius hibisci to Tetranychus urticae (Acari: Phytoseiidae, Tetranychidae). Exp. Appl. Acarol. 34, 263-273. [ Links ]
Barrera, A.J., V. Rodríguez y R. De Vis. 1997. Uso de Phytoseiulus y Neoseiulus californicus en el control de Tetranychus urticae en rosa en la Sabana de Bogotá. Parte I. Asocolflores 50, 14-16. [ Links ]
Bielza, P., V. Quinto, J. Contreras, M. Torne, A. Martín y P.J. Espinosa. 2007. Resistance to spinosad in the western flower thrips, Frankliniella occidentalis (Pergande), in greenhouses of south-eastern Spain. Pest. Mgt. Sci. 63, 682-687. [ Links ]
Blais, J. y E. Maly. 1993. Differential predation by Chaoborus americanus on males and females of two species of diaptomus. Can. J. Fish. Aqua. Sci. 50, 410-415. [ Links ]
Boer, J., T. Snoeren y M. Dicke. 2005. Predatory mites learn to discriminate between plant volatiles induced by prey and nonprey herbivores. Anim. Behav. 69, 869-879. [ Links ]
Burger, J.M.S., A. Kormany, J.C. Van Lenteren y L.E.M. Vet. 2005. Importance of host feeding for parasitoids that attack honeydew-producing hosts. Entomol. Exp. App. 117, 147-154. [ Links ]
Cadogan, B.L. y J.E. Laing. 1977. A technique for rearing the predaceous mite Balaustium putmani (Acarina: Erythraeidae), with notes on its biology and life history. Can. Entomol. 109(12), 1535-1544. [ Links ]
Cardona, C., F. Rendón, J. García, A. López-Ávila, J. Bueno y J. Ramírez. 2001. Resistencia a insecticidas en Bemisia tabaci y Trialeurodes vaporariorum (Homoptera: Aleyrodidae) en Colombia y Ecuador. Rev. Colomb. Entomol. 27(1-2), 33-38. [ Links ]
Carrel, J. y E. Tanner. 2002. Sex-specific food preferences in the madagascar hissing cockroach Gromphadorhina portentosa (Dictyoptera: Blaberidae). J. Insect Behav. 5(5), 707-714. [ Links ]
Choh Y. y J. Takabayashi. 2007. Predator avoidance in phytophagous mites: Response to present danger depends on alternative host quality. Oecologia 151(2), 262-267. [ Links ]
Collier, K.F.S., G.S. Albuquerque, J.O.G. De Lima, A. Pallini y A.J. Molina-Rugama. 2007. Neoseiulus idaeus (Acari: Phytoseiidae) as a potential biocontrol agent of the two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae) in papaya: performance on different prey stage - host plant combinations. Exp. Appl. Acarol. 41, 27-36. [ Links ]
De Bruijn, P.J., M. Egas, A. Janssen y M.W. Sabelis. 2006. Pheromone-induced priming of a defensive response in Western flower thrips. J. Chem. Ecol. 32, 1599-1603. [ Links ]
Dicke, M., M.W. Sabelis, M.D. Jong y M.P.T. Alers. 1990. Do phytoseiid mites select the best prey species in terms of reproductive success? Exp. Appl. Acarol. 8, 161-173. [ Links ]
Ferkovich, S.M., R. Venkatesan, J. Shapiro y J.E. Carpenter. 2007. Presentation of artificial diet: effects of composition and size of prey and diet domes on egg production by Orius insidiosus (heteroptera: Anthocoridae). Flor. Entomol. 90(3). 502-508. [ Links ]
Flores, A., G. Silva, M. Tapia y P. Casals. 2007. Susceptibilidad de Tetranychus urticae Koch (Acari: Tetranychidae) colectada en Primula obconica Hance y L. a acaricidas. Agr. Tec. Chile 67(2), 219-224. [ Links ]
Flórez, V.J., A. Leyba, G. Reyes y G. Fischer. 2000. Diagnóstico y evaluación en procedimientos en la poscosecha de rosas. Asocolflores 59, 48-52. [ Links ]
Getiva, J.C. y A. Acosta. 2004. Taxonomía de ácaros asociados a cultivos de flores. Asocolflores 66, 59-68. [ Links ]
Gnanvossou D., R. Hanna y M. Dicke. 2002. Prey-related odor preference of the predatory mites Typhlodromalus manihoti and Typhlodromalus aripo (Acari: Phytoseiidae). Exp. Appl. Acarol. 27, 39-56. [ Links ]
Gnanvossou, D., J.S. Yaninek, R. Hanna y M. Dicke. 2003. Effects of prey mite species on life history of the phytoseiid predators Typhlodromalus manihoti and Typhlodromalus aripo. Exp. Appl. Acarol. 30, 265-278. [ Links ]
Gutierrez, C. 2007. Pre-pupation behavior of the aphid parasitoid Aphidius ervi (Haliday) and its consequences for pre-imaginal learning. J. Naturwiss. 94, 595-600. [ Links ]
Haitlinger R. 2005. A new genus and four new species of mites from Argentina, Brazil and Nicaragua (Acari: Prostigmata: Erythraeidae, Eutrombidiidae). Genus 16, 513-525. [ Links ]
Halliday, R.B. 2001. Systematics and biology of the Australian species of Balaustium von Heyden (Acari: Erythraeidae). Aust. J. Entomol. 40, 326-330. [ Links ]
Hayes, L.J. 1985. The predator-Prey Interaction of the mite Balaustium sp. and the pierid butterfly Colias alezandra. Ecol. 66(1), 300-303. [ Links ]
Lang, A. y S. Gsödl, 2001. Prey vulnerability and active predator choice as determinants of prey selection: a carabid beetle and its aphid prey. J. Appl. Entomol. 125, 53-61. [ Links ]
Mendes, S., V. Bueno, V. Argolo y L. Silveira. 2002. Type of prey influences biology and consumption rate of Orius insidiosus (Say) (Hemiptera, Anthocoridae). Rev. Bras. Entomol. 46, 99-103. [ Links ]
Mendoza, L.A., A. Acosta, J. Ospina y L. Diaz. 2001. Evaluación de diferentes estados de ácaros del género Tetranychus, en cultivos de flores de la Sabana de Bogotá. Asocolflores 60, 11-24. [ Links ]
Nomikou, M., R. Men, R. Schraag, M.W. Sabelis y A. Janssen. 2005. How predatory mites find plants with whitefly prey. Exp. Appl. Acarol. 36, 263-275. [ Links ]
Oliveira, H., A. Janssen, A. Pallini, M. Venzon, M. Fadini y V. Duarte. 2007. A phytoseiid predator from the tropics as potential biological control agent for the spider mite Tetranychus urticae Koch (Acari: Tetranychidae). Biol. Control 42, 105-109. [ Links ]
Overmeer, W.P.J., H.J.C.F. Nelis, A.P. De Leenheer, J.M.N. Calis y A. Veerman. 1989. Effect of diet on the photoperiodic induction of diapause in three species of predatory mites, Amblyseius potentillae, Amblyseius cucumeris and Typhlodromus pyri. Exp. Appl. Acarol. 7, 281-288. [ Links ]
Pallini, A., A. Janssen y M. Sabelis. 1997. Odour-mediated responses of phytophagous mites to conspecific and heterospecific competitors. Oecologia, 110, 179-185. [ Links ]
Peckarsky, B. 1980. Predator - prey interactions between stone-flies and mayflies: behavioral observations. Ecol. 61, 932-943. [ Links ]
Putman, W. 1970. Life history and behavior of Balaustium putmani (Acarina: Erythraeidae). Ann. Entomol. Soc. Amer. 63(1), 76-81. [ Links ]
Reitz, S., J. Funderburk y S. Waring. 2006. Differential predation by the generalist predator Orius insidiosus on congeneric species of thrips that vary in size and behavior. Entomol. Exp. Appl. 119, 179-188. [ Links ]
Rodríguez-Reina, J.M., F. García-Marí y F. Ferraguta. 1992. Actividad depredadora de varios ácaros fitoseidos sobre distintos estados de desarrollo del trips de las flores Frankliniella occidentalis (Pergande). Bol. San. Veg. Plagas 18, 253-26. [ Links ]
Roger, C., D. Coderre y G. Boivin. 2000. Differential prey utilization by the generalist predator Colemegilla maculata lengi according to prey size and species. Entomol. Exp. Appl. 94, 3-13. [ Links ]
Salem, A.J. 2006. Functional and numerical responses of the predatory bug Macrolophus caliginous Wagner fed on different densities of eggs of the greenhouse whitefly, Trialeurodes vaporariorum (Westwood). J. Biol. Res. 6, 147-154. [ Links ]
Symondson, W., K. Sundrerland y M. Greenstone. 2002. Can generalist predators be effective biocontrol agents? Annu. Rev. Entomol. 47, 561-94. [ Links ]
Torrado, E., M.M. Pérez, J.R. Cure, M. García y C. Echeverri. 2001. Evaluación de sistemas de control biológico utilizados comercialmente en Europa para el control de plagas de rosa bajo invernadero, en la Sabana de Bogotá. Asocolflores 61, 34-45. [ Links ]
Veerman, A. 1992. Diapause in phytoseiid mites: a review. Exp. Appl. Acarol. 14, 1-60. [ Links ]
Venzon, M., A. Pallini y A. Janssen. 2001. Interactions mediated by predators in arthropod food webs. Neotrop. Entomol. 30(1), 1-9. [ Links ]
Venzon, M., A. Janssen y M.W. Sabelis. 2002. Prey preference and reproductive success of the generalist predator Orius laevigatus. Oikos 97, 116-124. [ Links ]
Welbourn, W.C. y D. Jennings. 1991. Two new species of Erythraeidae (Acari: Prostigmata) associated with the spruce budworm, Choristoneura fumiferana (Clemens) (Lepidoptera: Tortricidae), in Maine. Can. Entomol. 123, 567-580. [ Links ]
Yoder, J.A., J.T. Ark, J.B. Benoit, E.J. Bellinger y K.M. Gribbins. 2006. Water balance components in Adults of Terrestrial Red Mite Balaustium sp. (Acarina: Erythraeidae). Ann. Entomol. Soc. Amer. 99(3), 560-566. [ Links ]

