










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.27 no.2 Bogotá May/Aug. 2009
FISIOLOGÍA DE CULTIVOS
Fabio Ernesto Martínez1,3, Jenny Sarmiento1, Gerhard Fischer1 y Francisco Jiménez2
1 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
2 Departamento de Asistencia Técnica, Monómeros Colombo-Venezolanos S.A., Bogotá (Colombia).
3 Autor de correspondencia. femartinezma@unal.edu.co
Fecha de recepción: 20 de enero de 2009. Aceptado para publicación: 2 de julio de 2009
RESUMEN
El desconocimiento de los signos y síntomas visibles de las deficiencias nutricionales no permite establecer oportunos correctivos para manejar plantas con suficiente vigor, producción y calidad de los frutos. Plantas de uchuva ecotipo Colombia, propagadas por semilla, se sembraron en sustrato de arena cuarcítica en materas de plástico (25 L), y se mantuvieron durante 135 días en condiciones de invernadero de plástico en Bogotá. Las deficiencias nutricionales se indujeron a través de soluciones nutritivas, usando el método del elemento faltante y empleando un diseño completamente al azar con ocho tratamientos y cinco repeticiones, a saber: T0. testigo con fertilización completa; T1. fertilización completa -N; T2. -P; T3. -K; T4. -Ca; T5. -Mg; T6. -B, y T7. agua potable únicamente. Solamente en el caso del nitrógeno se aplicó 15% del N de la fertilización completa. Quincenalmente se efectuó una descripción visual y se llevó un registro fotográfico de los síntomas manifestados. Las plantas con deficiencias de N, K y B presentaron los síntomas más severos, generando efectos negativos sobre la arquitectura y el deficiente porte de las plantas, además de causar una seria afección sobre el tejido vegetal. Las plantas deficientes B tomaron una forma de roseta, típico síntoma hipoplástico. Las uchuvas que sufrieron deficiencias de P, Ca, Mg presentaron distorsiones en el color del follaje, observándose para la deficiencia de P, una coloración púrpura tanto en hojas de la zona reproductiva de la planta como en las del tallo principal. Las deficiencias de Mg y Ca, respectivamente, indujeron clorosis foliar intervenal y coloraciones anaranjadas.
Palabras clave: solución nutritiva, registro fotográfico, clorosis, necrosis, síntoma hipoplástico.
ABSTRACT
The lack of knowledge about the visible signs and symptoms of nutritional deficiencies does not allow timely introducing plant management corrections as to obtain vigorous, productive and qualified crops. In the present experiment, which was carried out in the city of Bogota, seed propagated cape gooseberry plants of the Colombia ecotype were kept under plastic greenhouse conditions in 25 L plastic containers filled with quartz sand, for 135 days. Mineral deficiencies were induced through nutrient solutions treated with the missing element technique, and arrayed in a completely randomized design with five replications and eight treatments, which were: T0. complete fertilization control treatment; T1. complete fertilization without nitrogen (-N); T2. complete fertilization -P; T3. -K; T4. -Ca; T5. -Mg; T6. -B, and T7. tap water. Only in the case of nitrogen, 15% of full N fertilization was applied. Apparent symptoms were visually described and photographed every 2 weeks. Plants with N, K or B deficiencies expressed the most severe symptoms, including negative effects on plant size and architecture, and serious tissular affections. Those plants suffering from P, Ca or Mg deficiency showed distortions in leaf color. In the particular case of P, a purple color in the leaves of the generative shoots and the vegetative stem was observed. Foliar intervein chlorosis and orange colorations were respectively induced by Mg and Ca deficiencies.
Key words: nutrient solution, photographic record, chlorosis, necrosis, hypoplastic symptom.
Introducción
Siendo la uchuva (Physalis peruviana L.) una planta solanácea originaria de Perú, de hábito silvestre o semisilvestre (Fischer, 2000), en Colombia es cultivada por pequeños productores con bajo nivel de asistencia técnica, lo cual ha generado que diversas prácticas basadas en la experiencia de cada agricultor, lo que a su vez implica un alto consumo de insumos y ausencia de tecnología (Sanabria, 2005). Pese al insuficiente nivel tecnológico, la uchuva es el segundo fruto exportado de Colombia. Enfermedades, plagas y deficiencias nutricionales son los principales problemas encontrados en los sistemas de producción de los frutales tropicales de clima frío y frío moderado (Cano et al., 2000).La observación de los síntomas visibles de las alteraciones nutricionales es un método de diagnóstico cualitativo (Garate y Bonilla, 2008). La caracterización del desarrollo foliar y de la planta en general, sumado a los síntomas de deficiencias de nutrientes pueden ser una ayuda en el diagnóstico de desórdenes y desbalances nutritivos (Yeh et al., 2000).
Los efectos nutricionales dependen de la influencia que ejerce cada nutriente en particular sobre los procesos fisiológicos y bioquímicos de la planta (Mengel et al., 2001). Sin embargo, evaluar el efecto de los diferentes nutrientes resulta complejo debido a que el metabolismo celular es, de un lado, regulado por la totalidad de los nutrientes absorbidos y, de otro, por la intensidad de la asimilación fotosintética (Friedrich y Fischer, 2000).
Si la concentración de un elemento nutriente esencial en el tejido vegetal está por debajo del nivel necesario para un óptimo crecimiento, indica que la planta es deficiente en ese elemento, y se produce así una alteración en la ruta metabólica en la que participa dicho elemento, afectando además otros procesos inmediatamente involucrados (Epstein y Bloom, 2005).
Algunos elementos son móviles en el floema (N, K, P, Mg); otros son poco móviles (Ca, B) y son obtenidos del xilema, y otros son de movilidad intermedia (Fe, Mn, Cu, Zn, Mo) (Calderón, 1995; Gil, 2006). De hecho y como regla general, cuando empieza a manifestarse la deficiencia en los elementos móviles, el elemento que está almacenado en las hojas maduras tiende a desplazarse a las más jóvenes para cubrir sus necesidades, por lo que los síntomas de estos se presentan en las hojas maduras de la parte basal de la planta o ramas (Navarro y Navarro, 2000). Situación diferente ocurre en los elementos poco móviles que tienden a manifestarse primero en las hojas jóvenes, mientras los de movilidad intermedia se muestran, en muchos casos, primero en la parte intermedia de la planta (Calderón, 1995).
La aparición de síntomas muchas veces es la primera indicación de que una deficiencia está limitando el crecimiento y a partir de este momento se requiere una cuidadosa observación y descripción de ellos, ya que se relaciona con la función y distribución de los nutrientes y podrían ser confirmados mediante el análisis de los tejidos (Calderón, 1995).
Los primeros estudios en síntomas de deficiencia en la uchuva fueron realizados por Garg y Singh (1975), refiriéndose únicamente a N, P y K. Los nutrientes considerados en la presente investigación fueron seleccionados de acuerdo con el siguiente criterio: los macronutrientes (N, P, K, Ca y Mg) fueron incluidos teniendo en cuenta que el tejido de las plantas los contiene en elevadas concentraciones (Epstein y Bloom, 2005), llegando sus deficiencias a determinar reducciones marcadas en el crecimiento, desarrollo y rendimiento. Adicionalmente se incluyó el B, ya que se encuentra estrecho relacionado con cambios estructurales en las paredes celulares, las cuales llegan a tener hasta un 90% del contenido celular de este elemento (Loomis y Durst, 1992). De hecho, la deficiencia de B puede llegar a determinar condiciones tisulares muy anormales (Rice, 2007).
Las deficiencias de nutrientes en el cultivo de la uchuva reducen el rendimiento y la calidad del fruto (Martínez et al., 2008), y por ende, la rentabilidad del productor, por lo cual el objetivo de este estudio se centró en identificar los síntomas de deficiencia de varios nutrientes en esta especie con el fin de facilitar su reconocimiento en campo y para que el fruticultor pueda tomar medidas de corrección a tiempo.
Materiales y métodos
La investigación se realizó en la Facultad de Agronomía, Universidad Nacional de Colombia, sede Bogotá, en condiciones de invernadero de plástico. Se trasplantaron plantas de uchuva ecotipo Colombia, de 1 mes de edad, en materas de plástico, con una capacidad de 25 kg; estas a su vez se llenaron con arena cuarcítica con un tamaño de grano de 0,5 mm y una conductividad eléctrica de 0,01 dS m-1. Se implementó un diseño completamente al azar con ocho tratamientos y cinco repeticiones, distribuidos así: T0: fertilización completa (testigo); T1: fertilización completa menos nitrógeno (-N); T2: fertilización completa menos fósforo (-P); T3: fertilización completa menos potasio (-K); T4: (-Ca), T5: (-Mg), T6: (-B) y T7: aplicación de agua potable únicamente, sin ningún elemento adicional. En el caso sin nitrógeno se adicionó el 15% del N (fuente: nitrato de amonio), aplicado en el tratamiento completo (testigo) para garantizar un crecimiento mínimo de la planta (Martínez, 1997). Las plantas se ubicaron en cinco camas espaciadas a 1,5 m y dejando 1,5 m entre plantas.
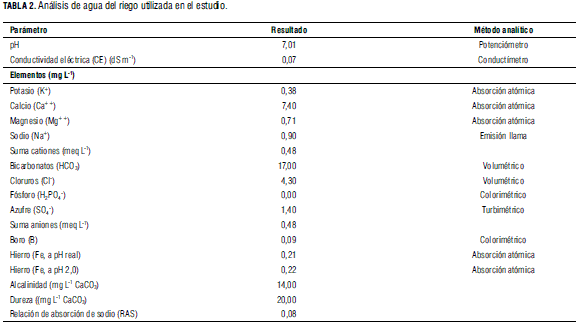
Para evaluar el efecto de los tratamientos, se utilizó la solución de Hoagland y Arnon (1950) modificada (Tab. 1), la cual fue mezclada para generar soluciones madre en tanques de 40 L; esta fue aplicada dos veces por día mediante regadera. Al agua de riego se le realizó un análisis fisicoquímico (Tab. 2) con el fin de verificar que fuera apta para el estudio.
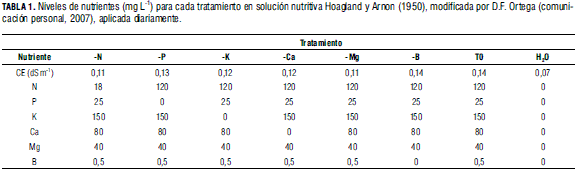
Las fuentes utilizadas fueron: nitrato de amonio líquido, fosfato monopotásico, DAP, nitrato de potasio, sulfato de potasio, nitrato de calcio líquido, mainstain calcio, sulfato de magnesio, EDTA -Fe 11%, cosmoquel B y molibdato de amonio. El sistema de tutorado de las plantas utilizado fue en 'V' y no se realizó ningún tipo de poda.
Las evaluaciones se tomaron con una frecuencia de 15 d (durante 7 meses) a partir de la fecha de la siembra realizando una descripción de los síntomas manifestados por las plantas y un registro fotográfico del desarrollo de los mismos para cada una de las deficiencias. Con el fin de constatar las deficiencias de nutrientes se efectuó un análisis foliar en cada planta a los 135 días después de la siembra (dds).
Resultados y discusión
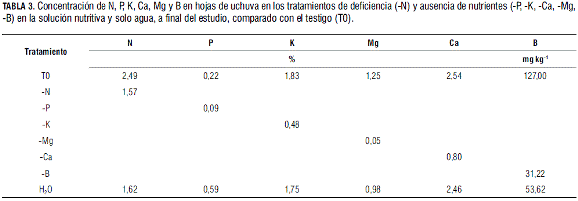
Las plantas regadas solamente con agua presentaron una reducción drástica de crecimiento, especialmente por la deficiente nutrición en N (Tab. 3) y una clorosis severa generalizada; además no indujeron flores y por consiguiente frutos, comparados con el testigo (Fig. 1A).

Nitrógeno
El N es un constituyente de un gran número de compuestos orgánicos, incluidas hormonas de crecimiento (Wild y Jones, 1992) esenciales en el metabolismo de la planta, forma parte de la estructura de todas las proteínas, de los ácidos nucleicos (ADN, ARN), y además se encuentra como constituyente de las clorofilas y enzimas del grupo de los citocromos, indispensables para la fotosíntesis y la respiración, y en varias coenzimas, como los nicotinamida-adenin-dinacleótidos (NAD, NADP) (Navarro y Navarro, 2000).
Las plantas con deficiencia de nitrógeno (Tab. 3) presentaron un menor porte (Fig. 1B), solamente 99,6 cm de altura comparado con el testigo (216,2 cm), lo cual no es sorprendente ya que el N es el cuarto elemento más abundante en la planta, después del C, O y H (Navarro y Navarro, 2000); además mostraron un hábito etiolado (Epstein y Bloom, 2005). Tuvieron tallos y ramas delgados con entrenudos cortos, hojas de reducido tamaño y grosor (Garg y Singh, 1975) y desprendieron prematuramente hojas antiguas, como también observaron en otras solanáceas, deficientes de este elemento, Cabezas et al. (2002) en lulo, Bergmann (1993) en tomate, y Avilán (1975) en guanábana. Se presentó una coloración verde pálido generalizada (Fig. 1C), a causa de la pérdida de clorofila (Calderón, 1995), que fue más acentuada en hojas bajeras (Gómez, 2006); también se desarrolló una coloración púrpura intervenal muy marcada tanto en lámina como en pecíolo. Este síntoma se presenta en caso de una grave deficiencia nitrogenada (Navarro y Navarro, 2000), lo cual explica Bergmann (1993) como una mayor síntesis de carbohidratos en plantas carentes de N, que conduce a formar más antocianinas.
Se evidenció un retardo en la ramificación, floración y fructificación (8-10 d después del testigo), también reportado por Garg y Singh (1975) en esta especie y por Gómez (2006) en otras plantas. En ramas se manifestó una coloración púrpura y la floración fue reducida. Al igual que en las hojas, los cálices (que cubren el fruto durante todo su desarrollo) exhibieron un color verde claro generalizado acompañado de pigmentación púrpura más acentuada hacia la zona de inserción del pedúnculo y venación rojiza. El color de los frutos fue de un naranja más claro.
Fósforo
El P, no obstante ser un elemento usado en bajas cantidades en la planta (Mengel et al., 2001), desempeña un rol muy importante en un gran número de reacciones enzimáticas que dependen de la fosforilación, incluida la incorporación del fosfato en los nucleótidos (ADP, ATP), por lo que el fósforo representa una función fundamental en la conservación y transferencia de energía. Los fosfatos son, también, constituyentes de los ácidos nucleicos y de los fosfolípidos, además son esenciales para la división celular y para el desarrollo de los tejidos meristemáticos (Wild y Jones, 1992); asimismo intervienen en la utilización del azúcar y del almidón (Mengel et al., 2001).
Con la deficiencia de fósforo (Tab. 3) se observó en las plantas de uchuva una coloración verde levemente más oscura que en el testigo, coincidiendo con Garg y Singh (1975) (Fig. 2A), pero no hasta verde-azul como describen Epstein y Bloom (2005) esta deficiencia. No se presentaron cambios severos en el porte de la planta, ya que no hubo diferencia significativa entre el tratamiento testigo y el sin P para la variable altura de la planta (Martínez y Sarmiento, 2008). Las hojas de la parte bajera de la planta mostraron una coloración púrpura a parda en la venación principal (Epstein y Bloom, 2005) iniciando en el ápice, distribuida a manera de moteado y que posteriormente abarcó la totalidad de la hoja (Fig. 2B).

Las hojas exhibieron una consistencia acartonada y textura rugosa (Friedrich y Fischer, 2000). Este síntoma fue más acentuado en los nudos basales, antes de la bifurcación. Igualmente, las hojas generativas ubicadas después del primer nudo en la segunda bifurcación manifestaron la misma coloración púrpura intensa, más acentuada en la base y márgenes foliares, pero que en estados finales alcanzó toda la lámina (Fig. 2C). Los síntomas fueron más apreciables en el primer y segundo tercio de la planta (Gómez, 2006). En estados finales las hojas se tornaron completamente púrpuras a pardo-amarillo, y subsiguientemente se dio la abscisión foliar prematura, similar a lo observado por Garg y Singh (1975) en donde las hojas en sus últimos estados se tornaron de un color verde-grisáceo, con eventual secamiento y caída prematura.
Potasio
La planta exige el K para generar la turgencia y el mantenimiento del potencial osmótico celular, regulando en las células guarda la apertura de los estomas (Bennet, 1993). Además, según el mismo autor, el K funciona como un estabilizador del pH celular, contrarresta las cargas negativas de los ácidos orgánicos y aniones inorgánicos, y es activador para más de 60 enzimas en el tejido meristemático. También actúa en la producción del ATP, en la síntesis de almidón y proteínas, y en el proceso de fotosíntesis y el metabolismo de los carbohidratos (Samra y Arora, 1997).
Las deficiencias de K se presentan con mayor frecuencia en suelos arenosos (Wild y Jones, 1992), lo que incrementó la manifestación de síntomas de deficiencia en este estudio. Junto con la carencia en boro y nitrógeno, esta deficiencia (Tab. 3) fue la que más afectó el porte de la planta y mostró los síntomas foliares más severos (Fig. 3A) causando una disminución del tamaño de la planta en general, como también lo reportaron Gargh y Singh (1975) en esta especie y Marschner (2002) en general. En el tallo se observaron entrenudos cortos, similar a lo manifestado en fríjol (Bergmann, 1993), ramas poco elongadas (Gómez, 2006) y más delgadas que las del testigo, lo que concuerda con lo reportado por Pissarek y Finck (1972) en maíz debido a un menor grosor del sistema vascular. Algo semejante se presenta en los cítricos, en los cuales una deficiencia muy fuerte del potasio resulta en una pérdida de rigidez y posterior doblamiento de ramas (Montoya et al., 2005).

En el tejido foliar el daño inició con unas manchas o puntos muy pequeños de forma irregular, con un tono más claro y de apariencia húmeda, distribuidos de forma intervenal (Garg y Singh, 1975) en toda la lámina, lo que está relacionado con el colapso de los cloroplastos (Mengel et al., 2001). Posteriormente incrementaron su tamaño, tornándose en pudriciones secas o manchas necróticas, y se rodearon de un halo clorótico que luego tomó un color amarillo a anaranjado intenso (Fig. 3B), similar a lo reportado por Chang y Liang (1978) en algodón. Estas pudriciones secas incrementaron su área afectando por completo el tejido de la hoja, que finalmente se volvió totalmente amarilla, secándose progresivamente del ápice a la base (Wild y Jones, 1989) en el caso de hojas generativas, o de la base al ápice en hojas bajeras. Tal como en otra solanácea, como el lulo (Cabezas et al., 2002), la deficiencia de potasio se presentó especialmente en las hojas adultas y de estas en sus bordes, donde empezó a manifestarse una necrosis perimetral, la cual aumentó progresivamente y luego alcanzó toda la lámina foliar.
La planta perdió la totalidad de sus hojas en el primer y segundo tercio del dosel, lo que describieron Garg y Singh (1975) como una abscisión prematura de hojas en uchuva debido a esta deficiencia nutricional. Los síntomas se presentaron inicialmente en hojas vegetativas o bajeras (Mengel et al., 2001), las cuales además presentaron una superficie corrugada (Garg y Singh, 1975; Mengel et al., 2001) y epinastias; posteriormente las hojas jóvenes mostraron los mismos síntomas. Las ramas de segundo orden tomaron una tonalidad parda y se secaron progresivamente iniciando desde la zona apical o zonas de nuevo crecimiento hacia la base; finalmente, al igual que las hojas, estas se secaron totalmente y murieron, síntomas que describen Epstein y Bloom (2005) como "dieback" (muerte descendente) en condiciones de deficiencia de K severa.
Se presentó el mismo daño de tejido en el cáliz de los frutos. Inició con puntos y manchas de poco tamaño y de forma irregular, acentuándose más en el tercio medio del cáliz (Fig. 3C), y se expandió hacia su ápice hasta cubrir gran parte del capacho. Finalmente se observaron cáliz y pedúnculos completamente secos y de consistencia crocante. Los frutos maduraron más lentamente, en contraste con los cítricos cuyos frutos bajo este desorden maduran prematuramente (Bergmann, 1993), a pesar de que se apreció madurez de cosecha en el cáliz.
Magnesio
El Mg es un componente específico de la clorofila (porfirina magnésica) en la que un átomo de magnesio está ligado a cuatro anillos pirrólicos (Wild y Jones, 1992). Cumple un rol específico como activador de enzimas envueltas en la respiración, fotosíntesis y síntesis de ADN y ARN (Taiz y Zeiger, 2006). Actúa como cofactor de la mayor parte de las enzimas que intervienen en la fosforilación, y su importancia es grande en la transferencia de la energía (Wild y Jones, 1992).
Los síntomas de este elemento se mostraron principalmente en hojas de ramas generativas del tercio medio de la planta (Fig. 4A), después de la segunda bifurcación, contrario a lo manifestado en los estudios de Mengel et al. (2001) y Epstein y Bloom (2005) en donde se observaron primero en hojas maduras. Además, en las hojas se presentó un moteado o clorosis intervenal (Mengel et al., 2001) de color verde muy claro, que inició en el ápice o por la zona media de la hoja y avanzó por los bordes hacia toda la lámina en forma de 'V' (Fig. 4B), síntoma similar a lo descrito por Navarro y Navarro (2000) en cítricos. En estados avanzados, se desarrollaron pigmentos púrpura (Gómez, 2006) o pardos (Epstein y Bloom, 2005) en las márgenes y entre las venas y las hojas y el cáliz (Fig. 4C), y se tornaron completamente cloróticas. No se apreció un efecto importante sobre el porte de la planta, siendo la deficiencia magnésica de menor impacto sobre el hábito de crecimiento de la planta (Marschner, 2002). Síntomas similares fueron reportados por Cabezas et al. (2002) en lulo, donde además las hojas jóvenes conservaron su color original pero se notaron flexuosas y brillantes.

Calcio
Según Navarro y Navarro (2000), una de la principales funciones del Ca en la planta es formar parte de la estructura de la protopectina, como agente cementante para mantener las células unidas, estando localizado en la lámina media y en la pared primaria celular. Entre las otras funciones del calcio, las describen estos autores como la regulación de la absorción de nitrógeno, su actuación en la traslocación de los carbohidratos y proteínas, la neutralización de los ácidos orgánicos y la activación de algunas enzimas como amilasa y fosfolipasa. Además actúa como segundo mensajero entre factores ambientales y diferentes respuestas de las plantas en crecimiento y desarrollo (Marschner, 2002).
Los síntomas de la deficiencia cálcica (Tab. 3) se observaron tanto en hojas bajeras como en hojas del tercio medio principalmente, es decir ubicadas después de la segunda bifurcación (Fig. 5A) que se diferenciaron de las observaciones de otros autores (Montoya et al., 2005; Gómez, 2006), los cuales indican que estos síntomas se presentan, más bien, en hojas jóvenes y superiores de la planta. Las hojas afectadas aceleraron su senescencia manifestando una fuerte clorosis o color anaranjado intenso, que inició desde la base de las hojas y se extendió a la totalidad de estas (Fig. 5B). La clorosis o color naranja intenso comenzó por lo general en un lóbulo de la base de la lámina foliar. Además se observaron puntos minúsculos necróticos en el margen foliar, lo cual fue descrito también por Bergmann (1993). En algunos casos los frutos mostraron manchas redondas o alargadas de color crema a blanco y de tamaños variables (Fig. 5C), pero no se observó el rajado de este órgano, descrito por Cooman et al. (2005) y Fischer (2005) debido a esta deficiencia. Angulo et al. (2005) reportan que la carencia de Ca se manifiesta en algunos frutos, especialmente de la parte superior de la planta, con cálices deformes que no cubren la totalidad del fruto y manchas necróticas en el ápice de este, similares a los que muestra el tomate por deficiencia de calcio.

Boro
El boro desempeña un rol primario en la biosíntesis y estructura de la pared celular y en la integridad de la membrana plasmática (Marschner, 2002); asimismo está involucrado en el transporte de azúcares, la lignificación de la pared celular (Loomis y Durst, 1992), la elongación celular, la síntesis de ácidos nucleicos y las respuestas hormonales (Taiz y Zeiger, 2006).
La deficiencia de boro (Tab. 3) afectó severamente la arquitectura de la planta. La planta en general tomó una forma de roseta (Friedrich y Fischer, 2000) que es un típico síntoma hipoplástico (Montoya et al., 2005), debido a la malformación de todas sus estructuras y a la desorganización de sus meristemos, lo que conduce, generalmente, a la muerte prematura de los ápices caulinares (Wild y Jones, 1992) acentuándose durante el transcurso de carencia de este elemento en el ensayo, como lo observaron Hernández y Pacheco (1986) en cardamomo. La baja concentración de B genera disminución rápida en el nivel de RNA y, en consecuencia, cesa la división celular en los meristemos (Wild y Jones, 1992).
Además de una fuerte disminución en el tamaño y porte por un retraso en el crecimiento (Navarro y Navarro, 2000), la planta de uchuva perdió elasticidad en sus tejidos, tornándose muy quebradiza (Fig. 6A). Las hojas del tallo principal y de las ramas generativas en el tercio medio de la planta mostraron una textura corrugada y una consistencia acartonada o crocante muy fuerte, retorciéndose (Navarro y Navarro, 2000) y tomando forma de cilindro o cono, porque no se presentó una expansión completa de la lámina foliar (Yeh et al., 2000). Schopfer y Brennicke (2006) explican la consistencia crocante y quebradiza de los tejidos en las plantas carentes de B debido a la irregular formación de la pared celular por una ineficiente reticulación de la pectina.

Adicionalmente se presentó una clorosis foliar generalizada (Navarro y Navarro, 2000), que fue más acentuada en el tercio medio, y se desarrollaron pigmentos púrpura a manera de mosaicos en las venas, expandiéndose desde el ápice a toda la lámina (Fig. 6B). En estados finales se presentaron pudriciones secas de forma irregular extendidas entre las venas (Navarro y Navarro, 2000).
Similar a lo reportado por Cabezas et al. (2002) en lulo (Solanum quitoense), se observó daño en los meristemos y zonas de crecimiento atrofiadas en uchuva; en consecuencia, las hojas del tercio superior se malformaron tomando la apariencia de una gota o lanza, por la deformación de su base. Como una posible explicación para este síntoma, Christensen et al. (1978) afirmaron que las células pueden continuar dividiéndose pero la estructura de los nuevos tejidos no se forma completamente.
Igualmente se observaron hiponastias (Gómez, 2006), y superficies corrugadas, abultadas y brillantes. Se apreciaron clorosis irregulares en la lámina foliar (Navarro y Navarro, 2000), ya fuese en forma de puntos o manchas diminutas repartidas a lo largo de la hoja, o clorosis parciales acompañadas de coloraciones púrpura muy acentuadas.
Las ramas generativas desarrollaron entrenudos muy cortos y fueron excesivamente quebradizas, similar a lo descrito por Christensen et al. (1978) en la vid. En las zonas apicales se presentó una excesiva brotación de tallos cortos y deformados, semejante a lo observado por Sindoni et al. (1994) en ajonjolí, en el cual la deficiencia severa de B activó las yemas axilares acortando los entrenudos apicales y dando a las hojas un aspecto de roseta. Hubo excesiva caída de flores y aborto de frutos (Gómez, 2006). Los que se desarrollaron, presentaron un tamaño muy reducido, deformaciones (Fig. 6C) (Friedrich y Fischer, 2000), un fuerte daño en tejido debido a pudriciones en la zona de inserción del pedúnculo y cálices deformados. También se observó un reducido número de retoños basales comparado con el testigo.
Conclusiones
Los síntomas de deficiencia más severos se manifestaron por disminución marcada del N y ausencia completa de los elementos K y B en la solución nutritiva, en donde, además de efectos sobre la lámina foliar, se presentó un cambio drástico en la arquitectura y en la textura de los tejidos.
Las deficiencias de P, Mg y Ca indujeron un cambio en el color y la calidad de las hojas sin afectar en mayor grado el crecimiento de la planta.
La descripción de síntomas en la planta de uchuva y el material fotográfico presentado en este estudio facilitan el reconocimiento de estas deficiencias nutricionales y la toma de medidas correctivas en el cultivo por el fruticultor.
Literatura citada
Angulo, R., C. Bojacá, A. Cooman, D. Gómez y C. Torres. 2005. Resultados investigaciones CIA . pp. 49-77. En: Angulo, R. (ed.). Uchuva - el cultivo. Universidad de Bogotá Jorge Tadeo Lozano, Bogotá. [ Links ]
Bennett, W.F. (ed.). 1993. Nutrient deficiencies and toxicities in crop plants. APS Press, St. Paul, MN. [ Links ]
Avilán, L. 1975. Efecto de omisión de los macronutrientes en el desarrollo y composición química de la guanábana (Annona muricata L.) cultivada en soluciones nutritivas. Agron. Trop. 25(1), 73-79. [ Links ]
Bergmann, W. 1993. Ernährungsstörungen bei Kulturpflanzen. 3a ed. Gustav Fischer Verlag Jena, Stuttgart, Alemania. [ Links ]
Cabezas, M., T. Botía y L. Medina. 2002. Determinación de síntomas por deficiencia inducida de nutrimientos en lulo (Solanum quitoense Lamb). pp. 176-181. Memorias IV seminario nacional de frutales de clima frío moderado. Medellín, Colombia. [ Links ]
Calderón, F. 1995. Concepción moderna de la nutrición vegetal. pp. 305-323. En: Silva, F. (ed.). Fundamentos para la interpretación de análisis de suelos, plantas y aguas para riego. 2a ed. Sociedad Colombiana de la Ciencia del Suelo, Bogotá. [ Links ]
Cano, D.M., O.A. Muriel, V.A. Tamayo, E.J. Bernal y Z.M. Hincapié. 2000. Efecto del nitrógeno y el potasio en la calidad del fruto de lulo 'La Selva' (Solanum quítense*Solanum hirtum). pp. 263-269. En: Memorias tercer seminario de frutales de clima frío moderado. Manizales, Colombia. [ Links ]
Chang, I.C. y T.Y. Liang. 1978. Effect of potassium fertilization on rice and cotton in Southeast China. Soil and Fertilizer Research Institute of Agriculture Academy of China Annual Report. Beijing, China. pp. 1-5. [ Links ]
Christensen, L.P., A.N. Kasimatis y F.L. Jensen. 1978. Grapevine nutrition and fertilization in the San Joaquin Valley. Div. Agric. Sci. University of California, Publication 4087. Davis, CA. [ Links ]
Cooman, A., C. Torres y G. Fischer. 2005. Determinación de las causas del rajado del fruto de uchuva (Physalis peruviana L.) bajo cubierta. II . Efecto de la oferta de calcio, boro y cobre. Agron. Colomb. 23, 74-82. [ Links ]
Epstein, E. y A.J. Bloom. 2005. Mineral nutrition of plants: principles and perspectives. Sinauer Associates Publishers, Sunderland, MA . [ Links ]
Fischer, G. 2000. Crecimiento y desarrollo. pp. 9-26. En: Flórez, V.J., G. Fischer y A.D. Sora (eds.). Producción, poscosecha y exportación de la uchuva (Physalis peruviana L.). Unibiblos, Universidad Nacional de Colombia, Bogotá. [ Links ]
Fischer, G. 2005. El problema del rajado del fruto de uchuva y su posible control. pp. 55-82. En: Fischer, G., D. Miranda, W. Piedrahita y J. Romero (eds.). Avances en cultivo, poscosecha y exportación de la uchuva (Physalis peruviana L.) en Colombia. Unibiblos, Universidad Nacional de Colombia, Bogotá. [ Links ]
Friedrich, G. y M. Fischer. 2000. Physiologische grundlagen des obstbaus. Ulmer Verlag, Stuttgart, Alemania. [ Links ]
Garate, A. e I. Bonilla. 2008. Nutrición mineral y producción vegetal. pp. 143-164. En: Azcón-Bieto, J. y M. Talón (eds.). Fundamentos de fisiología vegetal. 2a ed. McGraw-Hill Interamericana, Madrid. [ Links ]
Garg, R.C. y S.K. Singh. 1975. Primary nutrient deficiencies in cape gooseberry (Physalis peruviana L.). Progressive Hort. 7(2), 53-58. [ Links ]
Gil, G.F. 2006. Fruticultura: la producción de fruta. Fruta de climas templado y subtropical y uva de vino. 2a ed. Ediciones Universidad Católica de Chile, Santiago. pp. 401-461. [ Links ]
Gómez, M.I. 2006. Manual técnico de fertilización de cultivos. Microfertisa, Produmedios, Bogotá. [ Links ]
Hernández, R. y R. Pacheco. 1986. Caracterización de síntomas visuales de deficiencias nutricionales en cardamomo (Elettaria cardamomum). Agron. Cost. 10(1-2), 13-27. [ Links ]
Hoagland, D.C. y D.L. Arnon. 1950. The water culture methods for growing plants without soil. Circ. Calif. Agric. Exp. Stn. No. 347. [ Links ]
Loomis, W.D. y R.W. Durst. 1992. Chemistry and biology of boron. BioFactors 3, 229-239. [ Links ]
Marschner, H. 2002. Mineral nutrition of higher plants. Academic Press, London. [ Links ]
Martínez, F.E. y J. Sarmiento. 2008. Estudio de los síntomas de deficiencia de N, P, K, Ca, Mg y B en plantas de uchuva (Physalis peruviana L.). Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. [ Links ]
Martínez, F.E., J. Sarmiento, G. Fischer y F. Jiménez. 2008. Efecto de la deficiencia de N, P, K, Ca, Mg y B en componentes de producción y calidad de la uchuva (Physalis peruviana L.). Agron. Colomb. 26(3), 389-398. [ Links ]
Martínez, M. 1997. Mineral nutrient deficiency in plantain. Hohenheim Tropical Agricultural Series. University of Hohenheim, Stuttgart, Alemania. [ Links ]
Mengel, K., E.A. Kirkby, H. Kosegarten y T. Appel. 2001. Principles of plant nutrition. Kluwer Academic Publishers, Dordrecht, The Netherlands. [ Links ]
Montoya, R., R. Cabrales y J.R. Calderón. 2005. Fertilización y deficiencias. Universidad de Córdoba. Editorial Siglo XXI, Montería, Colombia. [ Links ]
Navarro, S. y G. Navarro. 2000. Química agrícola. Ediciones Mundi- Prensa, Madrid. [ Links ]
Pissarek, H.P. y A. Finck. 1972. Untersuchungen zur anatomischmikroskopischen diagnose des latenten kaliummangels. Landw. Forsch. 27(1), 241-248. [ Links ]
Rice, R.W. 2007. The physiological role of minerals in the plant. pp. 9-29. En: Datnoff, L.E., W.H. Elmer y D.M. Huber. (eds.). Mineral nutrition and plant disease. APS Press, St. Paul, MI . [ Links ]
Samra, J.S. e Y.K. Arora. 1997. Mineral nutrition. pp. 175-201. En: Litz, R.E. (ed.). The mango: botany, production and uses. CAB International, Wallingford, UK. [ Links ]
Sanabria, S. 2005. Situación actual de la uchuva en Colombia. pp. 1-8. En: Fischer, G., D. Miranda, W. Piedrahíta y J. Romero (eds.). 2005. Avances en cultivo, poscosecha y exportación de la uchuva (Physalis peruviana L.) en Colombia. Unibiblos, Universidad Nacional de Colombia, Bogotá. [ Links ]
Schopfer, P. y A. Brennicke, 2006. Pflanzenphysiologie. 6a ed. Elsevier GmbH, Munich, Alemania. pp. 299-309. [ Links ]
Sindoni, M., J. Zamora y R. Ramírez. 1994. Síntomas de deficiencia de boro y producción de materia seca en ajonjolí. Agron. Trop. 44(1), 135-150. [ Links ]
Taiz, L. y E. Zeiger. 2006. Plant physiology. 2a ed. Sinauer Associates, Sunderland, MA . [ Links ]
Wild, A. y L.H.P. Jones. 1992. Nutrición mineral de las plantas cultivadas. pp. 73-119. En: Wild, A. (ed.). Condiciones del suelo y desarrollo de las plantas según Russel. Ediciones Mundi- Prensa, Madrid. [ Links ]
Yeh, D.M., L. Lin y C.J. Wright. 2000. Effects of mineral nutrient deficiencies on leaf development, visual symptoms and shootroot ratio of Spathiphyllum. Scientia Hort. 86, 223-233. [ Links ]

