










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.27 no.2 Bogotá May/Aug. 2009
Carolina Cardozo1, Paola Rodríguez2 y Mauricio Marín1, 3
1 Laboratorio de Biología Celular y Molecular, Facultad de Ciencias, Universidad Nacional de Colombia, Medellín.
2 Centro de Investigaciones del Banano (Cenibanano), Augura, Medellín; actualmente: Investigación y Desarrollo, Bayer CropsScience S.A., Carepa-Antioquia (Colombia).
3 Autor de correspondencia. mamarinm@unalmed.edu.co
Fecha de recepción: 16 de febrero de 2009. Aceptado para publicación: 2 de julio de 2009
RESUMEN
La enfermedad del moko, causada por Ralstonia solanacearum, es uno de los problemas fitosanitarios más importantes de la agroindustria bananera de Urabá. Esta bacteria se caracteriza por presentar una gran plasticidad genética, lo cual dificulta el diseño de estrategias para su control. Con el fin de determinar los componentes del complejo de especies de R. solanacearum presentes en la región de Urabá, en esta investigación se realizó la caracterización genotípica de una población de R. solanacearum procedente de plantas de banano, arvenses y suelos de diversas plantaciones de esta zona. La evaluación se realizó mediante PCR múltiple y secuenciación de tres regiones del genoma (genes egl, fliC y región 16S del ADNr). Los resultados indican que todas las cepas bacteriales estudiadas están asociadas al filotipo II, y en su mayoría pertenecen al secuevar 4, aunque en el caso de algunos aislamientos no fue posible su filiación con ninguno de los tres secuevares reportados para la raza 2. Dos aislamientos obtenidos del departamento del Magdalena, utilizados con fines comparativos, resultaron estar asociados al secuevar 6. Los resultados de esta investigación plantean importantes aspectos epidemiológicos a ser tenidos en cuenta en el manejo de la enfermedad del moko en las plantaciones de banano del Urabá antioqueño.
Palabras clave: ADNr 16S, endoglucanasa, filotipo, flagelina, moko, secuevar.
ABSTRACT
Moko disease, caused by Ralstonia solanacearum, is one of the most important phytopathological problems of the banana agribusiness in Uraba (Colombia). This bacterium is featured by having high genetic plasticity, which difficults the design of control strategies. The aim of the present research was to establish the components of the R. solanacearum species complex, through the study of the genotypic features of a series of strains isolated from banana plants, weeds and soils from different plantations in Uraba. The analysis was carried out through multiple PCR and sequencing of three genomic regions (egl and fliC genes, and region 16S of 16S rDNA). The results indicate that all the studied isolates are associated to phylotype II, and most of them belong to sequevar 4. However, in the case of some strains, it was not possible to associate the PCR results with the sequevars that have been reported for race 2. Interestingly, two isolates from the department of Magdalena used as comparative controls for the bacterial population of Uraba, were found to belong to sequevar 6. These results raise important epidemiological aspects of the management of moko disease in the banana growing region of Uraba.
Key words: rDNA 16S, endoglucanase, phylotype, flageline, Moko disease, sequevar.
Introducción
La enfermedad del moko del plátano y banano causada por Ralstonia solanacearum (Smith) Yabuuchi et al. es uno de los problemas fitosanitarios más limitantes de este cultivo en Colombia, siendo los departamentos de Antioquia (en la región de Urabá) y Quindío las áreas más afectadas por esta enfermedad (Obregón, 2007). R. solanacearum es un fitopatógeno altamente agresivo, con una distribución global y un inusual amplio rango de hospederos de al menos 50 familias botánicas diferentes, incluyendo cultivos como papa, tomate, tabaco, banano, heliconias, anturios y maní (Hayward et al., 1998). En diferentes estudios se ha encontrado que R. solanacearum es una especie heterogénea que debe considerarse como un complejo, es decir, como un grupo de aislamientos relacionados cuyos miembros individuales pueden representar más que una especie (Taghavi et al., 1996; Fegan y Prior, 2005). Tradicionalmente los miembros de R. solanacearum han sido subdivididos en cinco razas con base en su rango de hospederos y en cinco biovares según su capacidad metabólica para la utilización de diversas fuentes de carbono. Una variante tropical del biovar 2 (2T o N2) es considerada por Hayward et al. (1992) como un sexto biovar. Desde el punto de vista genético, la especie se ha segmentado en dos divisiones (I y II) (Cook et al., 1989).
La división I comprende representantes de los biovares 3, 4 y 5, principalmente encontrados en Asia, mientras que la división II contiene los biovares 1, 2 y N2, de origen americano. Esta agrupación fue posteriormente confirmada mediante análisis de las secuencias 16S del ADNr (Taghavi et al., 1996). Por otra parte, Poussier et al. (2000) demostraron la existencia de un subgrupo de aislamientos de origen africano dentro del biovar 1. Fegan y Prior (2005) plantean la existencia de cuatro grupos genéticos denominados filotipos y correspondientes a diferentes orígenes geográficos (Asia sensu lato, América, Indonesia y África). Cada filotipo está conformado por un número de secuevares, que son definidos como un grupo de cepas con una secuencia altamente conservada dentro de una región determinada del genoma, que para el caso de R. solanacearum corresponde al gen endoglucanasa (egl). Los secuevares incluyen un número de líneas clonales identificadas a partir de análisis con marcadores moleculares tipo AFLP o rep-PCR (Fegan y Prior, 2005).
Uno de los primeros análisis genéticos de los aislamientos de R. solanacearum raza 2 fue realizado por Cook et al. (1989), quienes mediante la utilización de RFLP encontraron la existencia de tres genotipos definidos como MLG 24, MLG 25 y MLG 28. Posteriormente, con el desarrollo de los conceptos de filotipos y secuevares, se determinó que la raza 2 de R. solanacearum está asociada al filotipo II, y que las cepas de los MLG 24, 25 y 28 correspondían a los secuevares 3, 4 y 6, respectivamente (Fegan, 2005). Más recientemente, Fegan y Prior (2006), basados en secuencias del gen egl, demostraron que las cepas causantes del moko tenían un origen polifilético, con algunos genotipos muy relacionados con aislamientos patogénicos a tomate y papa. A pesar de los avances en la caracterización genética de las cepas de la raza 2 de R. solanacearum, es necesario continuar evaluando el grado de variabilidad de este patógeno en diferentes agroecosistemas, por cuanto se cree que la diversidad de dicha raza está subestimada (Fegan y Prior, 2006). Esta investigación se planteó con el fin de obtener la información relacionada con los componentes del complejo de especies de R. solanacearum en la zona bananera de Urabá, como una herramienta para el apoyo de los programas de manejo del moko de las musáceas en Colombia.
Materiales y métodos
Cepas bacteriales
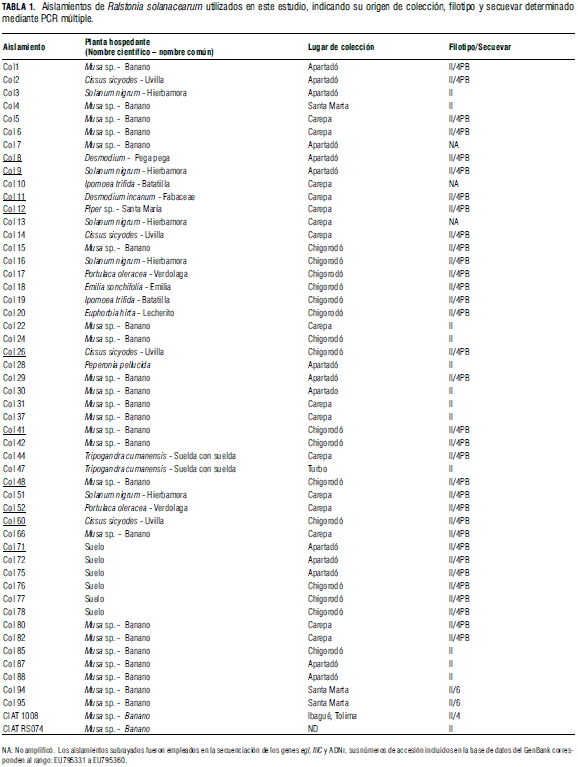
El estudio incluyó 50 aislamientos, 33 de los cuales hacen parte de una colección de cepas bacteriales previamente establecida por Cenibanano en plantas de banano y arvenses asociadas al cultivo (Obregón, 2007), mientras que las otras 17 cepas fueron aisladas y purificadas en esta investigación a partir de cultivos de banano y suelos de la región de Urabá y el departamento del Magdalena; estas últimas cepas fueron utilizadas con fines comparativos. Adicionalmente se incluyeron como control positivo dos cepas representativas del filotipo II proveídas por el CIAT (Tab. 1).
Aislamiento y purificación de las bacterias en medio de cultivo
Una vez detectadas las plantas de banano con sintomatología de marchitez bacterial, se procedió a aislar la bacteria a partir de pequeñas porciones de tejido vascular siguiendo la metodología de Gómez et al. (2005). Para el caso de las muestras de suelo provenientes de los focos de la enfermedad, se tomaron 500 g alrededor de las plantas y a una profundidad de 5 cm. Una vez en el laboratorio se realizaron diluciones seriadas en buffer fosfato 0,05 M a partir de 1 g de suelo, obteniendo 100 μL de las diluciones 10-1 y 10-3 para su siembra en medio SMSA modificado.
Confirmación de la identidad de los aislamientos
En todos los aislamientos utilizados en la investigación se verificó su identidad taxonómica mediante la utilización de los cebadores 759/760 (759: 5' GTC GCC GTC AAC TCA CTT TCC 3'; 760: 5' GTC GCC GTC AGC AAT GCG GAA TCG 3') (Fegan y Prior, 2005). Para esta prueba se tomó una lupada bacterial de cada aislamiento en 100 μL de agua destilada estéril y se llevó a ebullición al baño María por 5 min, utilizándose 1 μL en las reacciones de PCR.
Caracterización del complejo de especies R. solanacearum
Se utilizó la técnica de PCR múltiple para la definición de filotipos con los cebadores Nmult21:1F (5' CGT TGA TGA GGC GCG CAA TTT 3'), Nmult21:2F (5'AAG TTA TGG ACG GTG GAA GTC 3'), Nmult23:AF (5'ATT ACS AGA GCA ATC GAA AGA TT 3'), Nmult22:InF (5'ATT GCC AAG ACG AGA GAA GTA 3') y el cebador reverso Nmult22:RR (5' TCG CTT GAC CCT ATA ACG AGT A 3'), siguiendo la metodología descrita por Fegan y Prior (2005). Para el diagnóstico de los secuevares causantes del moko del banano se utilizaron los cebadores Mus20-F (5' CGG GTC GCT GAG ACG AAT ATC 3') - Mus20-R (5' GCC TTG TCC AGA ATC CGA ATG 3'); Mus35-F (5' GCA GTA AAG AAA CCC GGT GTT 3') - Mus35-R (5' TCT GGC GAA AGA CGG GAT GG 3'), Mus06-F (5' GCT GGC ATT GCT CCC GCT CAC 3') - Mus06-R (5' TCG CTT CCG CCA AGA CGC 3') y S128-F (5'CGT TCT CCT TGT CAG CGA TGG 3') - S128-R (5' CCC GTG TGA CCC CGA TAG C 3'). Las condiciones del PCR fueron similares a las descritas por Prior y Fegan (2005).
Para aquellos aislamientos en los que no fue posible la obtención de amplicones con alguno de los dos juegos de cebadores, se variaron las condiciones del PCR y se utilizaron dos metodologías adicionales de extracción de ADN: kit DNeasy Plant Mini (Qiagen, CA, USA) y el método convencional de SDS (Sambrook y Russell, 2001).
Análisis de secuencias de aislamientos de R. solanacearum
Se seleccionaron diez aislamientos de diferentes hospederos (Tab. 1) para la secuenciación parcial de la región 16S del ADNr utilizando los cebadores Oli1 (5'GGG GGT AGC TTG CTA CCT GCC 3') y Y2 (5'CCC ACT GCT GCC TCC CGT AGG AGT 3') (Seal et al., 1993), el gen egl con los cebadores Endo-F (5' ATG CAT GCC GCT GGT CGC CGC 3') y Endo-R (5` GCG TTG CCC GGC ACG AAC ACC 3`) (Fegan et al., 1998) y el gen fliC (flagelina C) con los cebadores Rsol_fliC-F: (5' GAA CGC CAA CGG TGC GAA CT 3') y Rsol_fliC-R (5' GGC GGC CTT CAG GGA GGT C 3') (Sehonfeld et al., 2003). El programa de amplificación constó de una desnaturalización inicial a 94oc por 3 min, seguida por 35 ciclos de 94°C durante 2 min; 62°C para fliC, 64°C para la región 16S del ADNr o 70°C para egl, por 1 min; 72oc por 2 min y un periodo final de extensión a 72°C durante 7 min. Los productos amplificados fueron purificados mediante el kit QIAquick PCR Purification (Qiagen, CA, USA) para proceder a su secuenciación directa en ambas direcciones en la compañía Macrogen (Corea del Sur). Las secuencias obtenidas con cada cebador fueron editadas mediante los programas BioEdit 6.0.6 y Chromas 1.45. Paralelamente, se obtuvieron del GenBank secuencias de los genes estudiados de cepas de R. solanacearum de diferentes regiones del mundo y hospederos, para su alineación con las generadas en este proyecto mediante el software Clustal W. La matriz de alineación fue empleada para el análisis filogenético basado en máxima parsimonia con soporte de Bootstrap (1000 permutaciones) y utilizando PAUP 4.0b.
Resultados
Confirmación de la identidad de los aislamientos La utilización de la prueba de PCR con los cebadores 759/760 permitió la selección de los 50 aislamientos incluidos en la investigación como asociados al complejo R. solanacearum, al obtenerse la amplificación del fragmento esperado de 282 pb.
Caracterización del complejo de especies R. solanacearum
La realización de las reacciones de PCR múltiples dirigidas a determinar los filotipos arrojó la amplificación de un fragmento de 372 pb en 47 de las cepas evaluadas (Fig. 1), mientras que no fue posible la obtención del amplicón en los aislamientos Col 7 (de Musa sp.), Col10 (de Ipomoea trifida) y Col13 (de S. nigrum). La ocurrencia de este amplicón permite asociar dichas cepas con el filotipo II y confirmar la presencia de dicho filotipo en hospederos tan diversos como Musa sp., Cissus sicyodes, Solanum nigrum, Desmodium sp., Piper sp., Portulaca oleraceae, Emilia sanchifolia, Ipomoea trifida, Euphorbia hirta, Peperonia pellucida, Tripogandra cumanenses, y en cepas aisladas a partir de suelos cultivados con banano (Tab. 1).
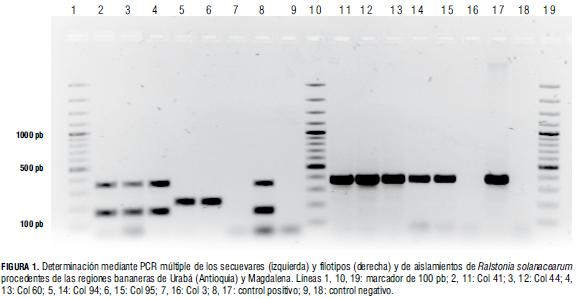
Con respecto a los PCR para secuevares, se logró la amplificación de un fragmento de 351 pb en 33 aislamientos y en la cepa de referencia RS 074 CIAT, mientras que en los aislamientos Col94 y Col95 la banda obtenida correspondió a 167 pb (Fig. 1). Este resultado permite identificar al primer grupo como cepas patogénicas a banano del secuevar 4, y al segundo grupo como cepas del secuevar 6. Es importante resaltar que en 15 de los aislamientos no fue posible la amplificación de ningún fragmento, a pesar de la modificación de diferentes variables del PCR.
Análisis de secuencias de aislamientos de R. solanacearum
Gen egl (endoglucanasa)
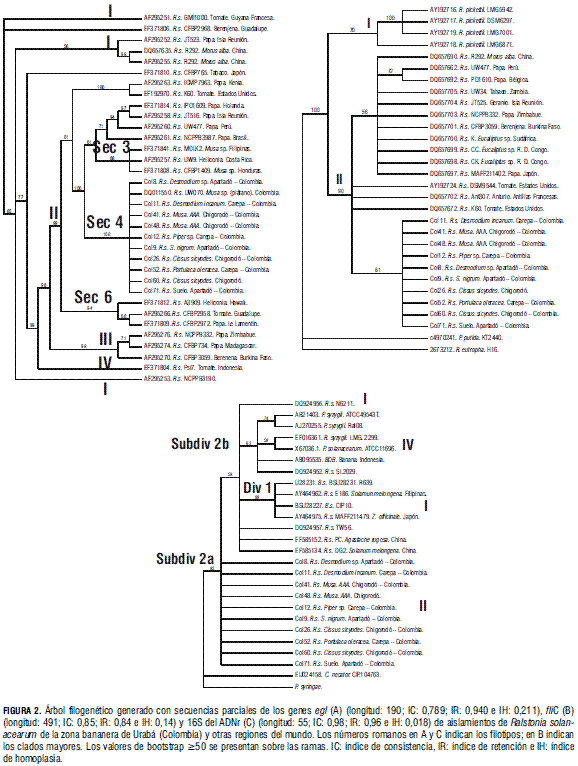
Para el análisis filogenético del gen egl se incluyeron secuencias de 34 aislamientos, 24 del Genbank, con al menos una cepa representativa de cada uno de los cuatro filotipos. El alineamiento final de las secuencias incluyó 686 sitios, de los cuales 107 resultaron informativos para el análisis de parsimonia (Fig. 2A). El filograma presentó cuatro clados bien definidos, que representan los cuatro filotipos del complejo. Sin embargo el filotipo I se presentó segmentado en un grupo principal con tres cepas (JT523, R292 y R292) y tres ramas independientes correspondientes a las cepas GMI100, CFBP2968 y NCPPB3190, cuyo origen corresponde a la división "Asiática". El filotipo II se presenta con un fuerte soporte estadístico (99%) y dividido en cuatro subgrupos, uno de los cuales incluye todas las cepas secuenciadas en esta investigación y procedentes de diferentes hospederos en la región de Urabá, además de una cepa obtenida de plátano en Colombia (Fegan y Prior, 2005). El grupo constituyendo el filotipo III presentó un alto soporte estadístico (99%) y estuvo conformado por tres aislamientos procedentes de papa y berenjena de países de África, mientras que el filotipo IV sólo estuvo representado en el análisis por el aislamiento PSI7 obtenido de tomate en Indonesia.
Gen fliC (flagelina C)
Para el análisis filogenético del gen fliC se incluyeron secuencias de 30 aislamientos, 20 de las cuales fueron obtenidas del Genbank, incluyendo cuatro representantes de la especie R. pickettii, y dos cepas de Pseudomonas putida y R. eutropha como grupos externos para el análisis (Fig. 2B). El alineamiento final de las secuencias incluyó 428 sitios, de los cuales 150 resultaron informativos. El árbol filogenético generado presentó dos clados bien soportados, que claramente separan las especies R. pickettii y R. solanacearum. En este último clado, se presentan varios subgrupos, pero con un débil soporte. Estos subgrupos no están diferenciados por filotipos, procedencia geográfica u hospedante, aunque todos los aislamientos obtenidos en la región de Urabá se agruparon en un solo clado, con un soporte del 86%.
Región 16S del ADNr
Para el análisis filogenético de la región 16S del ADNr se incluyeron secuencias de 26 aislamientos, 16 de las cuales fueron obtenidas del Genbank, dos cepas de P. syzygii, una cepa de BDB y Cupriavidus necator (R. eutropha) y P. syringae como grupos externos (Fig. 2C). El alineamiento final incluyó 289 sitios, nueve de los cuales resultaron informativos. El filograma generado presentó un clado bien soportado (93%) y claramente diferenciado de los grupos externos. Este clado representa el complejo de especies R. solanacearum, incluyendo P. syzygii, BDB y R. solanacearum sensu stricto. En el clado se presentaron tres subgrupos que representan el filotipo IV, el filotipo I y el filotipo II, respectivamente. Sin embargo, el bajo grado de polimorfismo encontrado entre las secuencias analizadas de R. solanacearum para esta región del genoma no permitió generar altos soportes de bootstrap para dichos grupos.
Discusión
En este estudio se realizó una caracterización molecular de los componentes del complejo de especies de R. solanacearum a partir de una población de 50 aislamientos procedentes de cultivos de banano, suelos y plantas arvenses de al menos ocho familias botánicas (Asteraceae, Commelinaceae, Convolvulaceae, Euphorbiaceae, Fabaceae, Piperaceae, Portulacaceae y Vitaceae) colectadas en la región de Urabá.
Los resultados indican que los 50 aislamientos evaluados están asociados al complejo R. solanacearum, tal como se desprende de la obtención de un amplicón de 282 pb en todas las cepas estudiadas. La secuenciación de la región 16S del ADNr permitió asociar a dichos aislamientos con la subdivisión 2A sensu (Poussier et al., 2000), la cual está conformada fundamentalmente por miembros del biovar 1, 2 y N2, aunque la definición original basada en análisis de RFLP que indica a la división 2 como "Americana" y planteada por Cook et al. (1989) queda desvirtuada en estos análisis, al incluir indistintamente aislamientos africanos y asiáticos obtenidos de musáceas y otros hospederos en estos continentes, lo cual reafirma la mayor resolución que ofrece el análisis de secuencias para plantear este tipo de hipótesis taxonómicas. De acuerdo con los resultados de la PCR múltiple con los cebadores de la serie Nmult, todos los aislamientos corresponden al filotipo II de R. solanacearum, grupo que incluye además los biovares 1, 2 y 2T de origen americano, la raza 3, patogénica a papa y con distribución global, y la raza 2 conocida como el agente causal del moko de las musáceas. Con respecto a los secuevares, se encontró que, de las 50 cepas analizadas, 33 se pueden identificar como pertenecientes al secuevar 4 y dos al secuevar 6, mientras que en 15 cepas no fue posible la amplificación, situación que sugiere la existencia de otros secuevares dentro de la raza 2 de R. solanacearum. Un resultado similar fue encontrado por Fegan y Prior (2006), quienes determinaron que las cepas ICMP6782 e ICMP9600, obtenidas de plantas de banano en Brasil, no se agruparon en los clados que definían los secuevares 3, 4 y 6 en su estudio basado en el análisis de secuencias del gen egl.
La ausencia del secuevar 3 y el bajo número de aislamientos del secuevar 6 encontrados en este trabajo fue inesperado, por cuanto dichos secuevares se han reportado frecuentemente en países cultivadores de banano de Centro y Suramérica, con los cuales existen vínculos comerciales y de intercambio de material de propagación con las zonas bananeras del país. Así por ejemplo, el secuevar 3, previamente definido como el MLG 24 (Cook y Sequeira, 1994) se reporta en cultivos de banano, plátano y heliconia de Costa Rica, Honduras y Panamá (Prior y Fegan, 2005), este último país, fronterizo con la región de Urabá. El secuevar 6, previamente definido como el MLG 28 (Cook y Sequeira, 1994), ha sido detectado en Honduras, Venezuela (Prior y Fegan, 2005) y recientemente en Guatemala por Sánchez et al. (2008). De otra parte, todos los aislamientos definidos como secuevar 4 (MLG25) en nuestro estudio presentaron la banda de 167 pb que identifica a dichas cepas como putativamente patogénicas a Musa sp. (Prior y Fegan, 2005). a patogenicidad en banano de las cepas procedentes de arvenses fue probada en un trabajo reciente realizado por Obregón (2007) en plantas del clon Gran enano. De gran interés resultó el hecho que las especies Desmodium sp., P. pellucida y T. cumanenses no habían sido reportadas como hospederos de la bacteria, y que la especie C. sicyodes representa el primer registro de la familia Vitaceae como hospedante de R. solanacearum. Estos hallazgos basados en pruebas de patogenicidad y evaluaciones moleculares amplían el conocimiento sobre la diversidad genética de esta bacteria en Suramérica y refuerzan las hipótesis de Fegan y Prior (2006), que plantean que es posible que el nivel de variabilidad que presenta R. solanacearum raza 2 sea mayor que el hasta ahora encontrado en los estudios realizados a nivel mundial.
El análisis filogenético realizado con las secuencias parciales del gen egl agrupó las cepas procedentes del Urabá con la cepa UW070, cuya secuencia (DQ011550) fue depositada en el Genbank por Fegan y Prior (2005) y que fue aislada de una planta de plátano en Colombia. Esta cepa había sido caracterizada como perteneciente al filotipo II/secuevar 4 SFR (Small, Fluidal, Round), lo cual coincide con el alto número de cepas que fueron encontradas en el presente estudio como pertenecientes a este secuevar. Este grupo se presentó asociado (con un soporte del 100%) al clado que contenía cepas que causan marchitamiento vascular en heliconia (Costa Rica) y Musa sp. en Filipinas (Bugtok) y Honduras (Moko), así como con el secuevar 1 que contiene cepas causantes de pudrición bacterial en papa. Adicionalmente, estos grupos se relacionaron con las secuencias representativas de cepas del secuevar 6 (heliconia, Hawai) y del secuevar 5 (papa, Isla Martinica y tomate, Isla de Guadalupe). Todo el clado presenta un soporte del 99%, constituyéndose en el subclado B reportado por Wicker et al. (2007), y confirmado por el análisis de Castillo y Greenberg (2007) con base en el análisis de secuencias de cinco genes del cromosoma bacterial y tres genes del megaplásmido.
Los demás clados que se conformaron en el análisis (subclado filotipo II-A, clado filotipo III y clado filotipo IV) tuvieron altos soportes bootstrap indicando su naturaleza monofilética. Sin embargo esta situación no ocurrió así para el filotipo I, que aunque en el análisis de máxima parsimonia se presentó como un solo grupo, no tuvo soporte estadístico, y por el contrario fue dividido en al menos tres grupos de cepas. Este resultado difiere del reportado por Villa et al. (2005), quienes utilizando análisis de secuencias de egl y hrpB confirmaron el carácter monofilético de este grupo de "origen asiático". Es evidente que dicha discrepancia se origina de las diferentes cepas utilizadas en los dos estudios, y hace necesario que futuros estudios incluyan un mayor número de aislamientos que confirmen o rechacen la hipótesis del carácter monofilético de dicho grupo.
De otra parte, el filograma generado a partir del análisis de secuencias del gen fliC generó dos grupos principales, que separaron las cepas de R. pickettii de aquellas representantes del complejo de especies R. solanacearum. Sin embargo este análisis no presentó el nivel de resolución necesario para diferenciar los filotipos en que se divide este complejo, ya que algunos clados estuvieron compartidos por aislamientos de diferentes orígenes filogenéticos. Este resultado sugiere que la utilidad del análisis de este gen debe restringirse al empleo de cebadores específicos Rsol_fliC para la detección de los miembros del complejo, aunque la sola prueba de PCR es insuficiente para lograr esto, pues se ha demostrado que dichos cebadores amplifican un fragmento de similar tamaño en P. syzigii, haciéndose necesaria la secuenciación de los fragmentos amplificados o el uso de la técnica DGGE previamente reportada por Schonfeld et al. (2003).
Conclusiones
Los hallazgos de esta investigación y de aquella realizada por Obregón (2007) reafirman la necesidad de eliminar completamente las plantas arvenses de los focos detectados con la enfermedad de moko en la región de Urabá, evitar al máximo el transporte de suelos infestados con la bacteria y garantizar la correcta desinfestación de las herramientas de trabajo utilizadas en las prácticas culturales, por cuanto la población de R. solanacearum en esta zona presenta un alto potencial de ser patogénica a plantas de banano. El establecimiento de prácticas cuarentenarias es fundamental para el manejo de esta enfermedad en la zona bananera de Urabá, de manera que se evite el ingreso de nuevos genotipos bacteriales en esta región, como por ejemplo de aquellos asociados al secuevar 6 detectados en este estudio en el departamento del Magdalena, o de cepas de otros países, que claramente en los análisis filogenéticos de secuencias se agruparon en clados diferentes a los ocupados por las cepas colombianas de esta bacteria.
Agradecimientos
Esta investigación tuvo financiación de Colciencias (8242- 07-16025), Augura y la Universidad Nacional de Colombia, sede Medellín. Los autores agradecen a Mónica Obregón y a Elizabeth Álvarez del CIAT, por proveer algunas de las cepas utilizadas en el estudio.
Literatura citada
Castillo, J.A. y J.T. Greenberg. 2007. Evolutionary dinamics of Ralstonia solanacearum. Appl. Environ. Microbiol. 73, 1225-1238. [ Links ]
Cook, D., E. Barlow y L. Sequeira. 1989. Genetic diversity of Pseudomonas solanacearum: detection of restriction fragment polymorphism with DNA probes that specify virulence and hypersensitive response. Mol. Plant Microbe Interact. 2, 113-121. [ Links ]
Cook, D. y L. Sequeira. 1994. Strain differentiation of Pseudomonas solanacearum by molecular genetic methods. pp. 77-93. En: Hayward, A.C. y G.L. Hartman (eds.). Bacterial wilt: the disease and its causative agent, Pseudomonas solanacearum. CAB International, Wallingford, UK. [ Links ]
Fegan, M. 2005. Bacterial wilt diseases of banana: evolution and ecology. pp. 379-386. En: Allen, C., P. Prior y A.C. Hayward (eds.). Bacterial wilt: the disease and the Ralstonia solanacearum species complex. APS Press, St. Paul, MN. [ Links ]
Fegan, M. y P. Prior. 2005. How complex is the "Ralstonia solanacearum species complex"? pp. 449-461. En: Allen, C., P. Prior y A.C. Hayward (eds.). Bacterial wilt: the disease and the Ralstonia solanacearum species complex. APS Press, St. Paul, MN. [ Links ]
Fegan, M. y P. Prior. 2006. Diverse members of the Ralstonia solanacearum species complex cause bacterial wilts of banana. Australas. Plant Pathol. 35, 93-101. [ Links ]
Gómez, E.A., E. Álvarez y G. Llano. 2005. Identificación y caracterización de cepas de Ralstonia solanacearum raza 2, agente causante del Moko de plátano en Colombia. Fitopatol. Colomb. 28, 71-75. [ Links ]
Hayward, A.C., L. Sequeira, E.R. French, H.M. El-Nashar y U. Nydegger. 1992. Tropical variant of biovar 2 of Pseudomonas solanacearum. Phytopathol. 82, 608. [ Links ]
Hayward, A.C., J.C. Elphinstone, D. Caffier, J. Janse, E. Stefani, E. French y A.J. Wrigth. 1998. Round table on bacterial wilt (brown rot) of potato. pp. 420-430. En: Prior, P., C. Allen y J.G. Elphinstone (eds.). Bacterial wilt disease: molecular and ecology aspects. Springer-Verlag, Berlin. [ Links ]
Obregón, M. 2007. Diagnóstico, hospederos y sobrevivencia de la bacteria Ralstonia solanacearum en banano y aplicaciones al control integrado de la enfermedad en la zona de Urabá. Tesis de maestría. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Medellín. [ Links ]
Prior, P. y M. Fegan. 2005. Diversity and molecular detection of Ralstonia solanacearum race 2 strains by multiplex PCR. pp. 405-414. En: Allen, C., P. Prior y A.C. Hayward (eds.). Bacterial wilt: the disease and the Ralstonia solanacearum species complex. APS Press, St. Paul, MN. [ Links ]
Sambrook, J. y D.W. Russell. 2001. Molecular cloning: a laboratory manual. 3a ed. Cold Spring Harbor Laboratory Press, Cold Spring, Harbor, NY. [ Links ]
Sánchez, A., L. Mejía, M. Fegan y C. Allen. 2008. Diversity and distribution of Ralstonia solanacearum strains in Guatemala and rare occurrence of tomato fruit infection. Plant Pathol. 57, 320-331. [ Links ]
Schonfeld, J., H. Heuer, J.D. Van Elsas y K. Smalla. 2003. Specific and sensitive detection of Ralstonia solanacearum in soil on the basis of PCR amplification of fliC fragments. Appl. Environ. Microbiol. 69, 7248-7256. [ Links ]
Seal, S.E., L.A. Jackson, J.P. Young y M.J. Daniels. 1993. Differentiation of a Pseudomonas solanacearum, Pseudomonas syzygii, Pseudomonas pickettii and the blood disease bacterium by partial 16S rRNA sequencing: construction of oligonucleotide primers for sensitive detection by polymerase chain reaction. J. Gen. Microbiol. 139, 1587-1594. [ Links ]
Taghavi, M., C. Hayward, L. Sly y M. Fegan. 1996. Analysis of the phylogenetic relationships of strains of Burkholderia solanacearum, Pseudomonas syzygii, and the blood disease bacterium of banana based on 16S rRNA gene sequences. Inter. J. Syst. Bacteriol. 46, 10-15. [ Links ]
Villa, J.E., K. Tsuchiya, M. Horita, M. Natural, N.Y. Opina y M. Hyakumachi. 2005. Phylogenetic relationships or Ralstonia solanacearum species complex strains from Asia and others continents based on 16S rDNA, endoglucanase, and hrpB gene sequences. J. Gen. Plant Pathol. 71, 39-46. [ Links ]

