










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.27 no.2 Bogotá May/Aug. 2009
Erika P. Martínez1, Juan C. Hío1, Jairo A. Osorio1, 2 and María F. Torres1
1 Laboratory of Phytopathology, Centro de Investigación Tibaitatá, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Mosquera (Colombia).
2 Corresponding author. josorio@corpoica.org.co
Received for publication: 29 october, 2008. Accepted for publication: 2 July, 2009
ABSTRACT
In Colombia, citrus, tree tomato and mango crops are likely to suffer considerable losses from anthracnose caused by several Colletotrichum species, which were identified by the present study on infected organs of the three fruit crops, sampled in different regions of the country. Identification was based on their morphological and molecular characteristics, as well as on fungicide (benomyl and copper hydroxide) sensitivity and pathogenicity tests. The latter assessed infectivity on both the original hosting crop and the other two crops (crossed infection), by putting the fungi in contact with organs taken from the three fruit crops. Molecular identification of the Colletotrichum species was carried out through amplification of rDNA ITS regions by means of C. gloeosporioides (CgInt) and C. acutatum (CaInt2) specific primer PCR combining the use of ITS4 universal primer. The results indicate that C. acutatum is the infectious agent in Tahiti lime and tree tomato, and so is C. gloeosporioides in mango. Although C. acutatum is the infectious agent in two different fruit species, the strains proved to be specific of their original hosts.
Key words: Colletotrichum acutatum, Colletotrichum gloeosporioides, tropical fruit, diseases.
RESUMEN
En Colombia los cultivos de cítricos, tomate de árbol y mango presentan elevadas pérdidas en su producción a causa de la antracnosis ocasionada por varias especies del género Colletotrichum. Por esta razón, en este estudio se identificaron las especies de Colletotrichum de los tres frutales, provenientes de varias zonas productoras del país, mediante sus características morfológicas, patogénicas, moleculares y de sensibilidad a los fungicidas benomil e hidróxido de cobre. Para la prueba de patogenicidad se colocó el hongo en contacto con órganos desprendidos de los tres frutales, para evaluar la infectividad de cada aislamiento sobre su hospedero de origen, y sobre las otras dos especies, a fin de detectar infección cruzada. La identificación molecular de las especies de Colletotrichum se realizó mediante amplificación de secuencias de las regiones ITS del ADN ribosomal por medio de PCR, utilizando cebadores específicos de C. gloeosporioides (CgInt) y de C. acutatum (CaInt2), en combinación con el iniciador universal ITS4. De acuerdo con los resultados obtenidos, se determinó que C. acutatum es el agente causal de la antracnosis en lima Tahití y en tomate de árbol, en tanto que C. gloeosporioides lo es en mango. A pesar de ser el agente causal de la enfermedad en dos especies frutales diferentes, C. acutatum mostró especificidad hacia su hospedero de origen.
Palabras clave: Colletotrichum acutatum, Colletotrichum gloeosporioides, frutas tropicales, enfermedades.
Introduction
Colletotrichum induced anthracnose is considered an important disease in Colombian fruit crops, due to the considerable losses it determines. In Tahiti lime (Citrus latifolia Tanaka), it is featured by mainly attacking the flowers, determining total fruit rottening and premature dropping (Agostini et al., 1992), and lowering productivity by 50% in the citrus growing regions of the country (Osorio, 2000).
In tree tomato (Solanum betaceum Cav.), anthracnose directly affects the fruit, producing oily stains that turn black as they grow in size. In commercial crops that receive continuous fungicide applications, losses range from 10 to 25% of the harvested fruit. When management measures are not efficient, losses can go up to 80 or even 100% of the harvest. This is, therefore, the most expensive of all the crop's issues, sometimes determining its abandonment or substitution (Tamayo, 2001).
In mango, (Mangifera indica L.) anthracnose mainly attacks inflorescences and fruits (both during ripening and post harvest), occasionally affecting young leaves. In physiologically mature fruits, the disease is featured by causing black or brown superficial damage (Arauz, 2000), determining losses that go up to 35% of the harvested fruit (Páez, 1995).
Controlling Colletotrichum is still a deficient task, as is our knowledge about the basic aspects of its biology such as the infectious agent and its genetic variability, inoculum dispersal and host specificity (one single Colletotrichum species has been found causing crossed infection on several hosts).
C. gloeosporioides and C. acutatum are the two species that have been commonly found in anthracnose infected fruit crops. Their identification is therefore a fundamental criterion in the development of more efficient control measures, as far as it allows better knowledge of the pathogen's epidemiological behavior (Freeman et al., 1998). However, due to their morphological variability, the ample range of their hosting crops, and the wide variety of their cultured isolates, they are partially difficult to identify by traditional taxonomic methods, which must then be complemented with molecular techniques (Andrade et al., 2007; Whitelaw-Weckert et al., 2007).
C. gloeosporioides and C. acutatum specific oligonucleotides have been widely used in differentiating these two species by means of PCR (Freeman et al., 1998, 2001; Peres et al., 2002b; Afanador et al., 2003; Sanabria, 2007). The clear identification of the infectious agent allowed by this technique has sometimes led to discarding crossed infection hypotheses (i.e. those stating that just one fungal species accounts for infecting different fruit crops) (Freeman and Katan, 1997).
In this framework, the objectives of the present work were to identify, by means of molecular and conventional identification techniques, the Colletotrichum species that are associated to anthracnose in Tahiti lime, tree tomato and mango in the main productive regions of Colombia; and to explore the possibility of crossed infection taking place between these fruit species.
Materials and methods
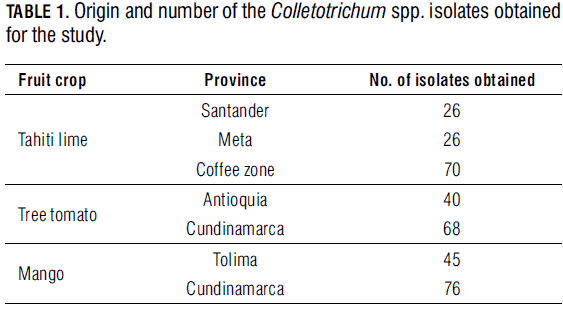
Three hundred and fifty one Merck PDA grown Colletotrichum isolates were obtained from anthracnose affected organs (Tahiti lime flowers, and mango and tree tomato fruits), collected in different productive provinces3 of Colombia (Tab. 1). The morphology of the colonies and fungal structures was registered after 10 days of inoculation at 23°C. The isolates were included and documented in the Colombian Colletotrichum Collection, with an internal code in the corresponding data base of the Laboratory of Phytopathology of Corpoica, at Centro de Investigación Tibaitata4. Each of the isolates was purified through monosporic culturing and then preserved in filter paper embedded in a 20% glycerol solution.
Morphological analysis Out of the above mentioned 351 isolates, 60 (20 of each fruit crop) were randomly chosen for their morphological analysis, which, after 5 days of growing at 25°C, consisted in registering color, general aspect, edge morphology and growth mode of the cultured colonies.
The morphological analysis of the conidia was carried out on 80 spores from each isolate. They were classified in three classes, according to their morphology: 0 (conidia rounded on both ends); 1 (1 round, 1 sharp ended conidia); and 2 (both end sharpened conidia) (Sutton, 1992).
Length and width of each conidia were additionally measured. The obtained results were statistically analyzed through a Ward algorithm conglomerate analysis (respective maximum and minimum inter and intra group variation), applied with a version 9.1.3 SAS® software package.
The mycelium obtained from the PDA and micro cultures was described through observing and registering length and general features of the terminal hyphae (Barnet and Barry, 2003) on trypan blue lactophenol optic microscope (40x) preparations. A statistical analysis of such morphometrical data was carried out by means of analysis of variance.
Benomyl and copper hydroxide susceptibility tests
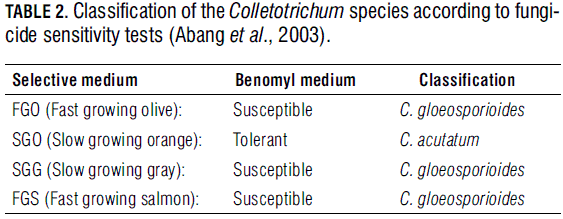
In order to determine fungicide susceptibility of the pathogens, a hundred 0.5 cm agar discs were taken from equal number of randomly chosen isolates of each fruit crop (for a total of 300 isolates), to be grown in solid culture media containing either benomyl or copper hydroxide as fungicides. The benomyl medium was prepared with a 2 μg mL-1 solution of the product dissolved in PDA. After 72 h of incubation in the darkness at 27°C, colony diameter was measured and compared to that of a non-fungicide added medium. The (copper hydroxide) selective medium contained 42 mg of metallic copper (kocide 2000: 53.8% of Cu (OH)2) plus 300 mg of streptomycin sulfate per liter of PDA. After incubation, and under the same conditions of the benomyl test, color and diameter of the colonies were compared to those of the control test. The results of the two tests were interpreted according to the classification chart shown in Tab. 2.
Pathogenicity tests
Pathogenicity tests were carried out by inoculating randomly chosen isolates on: 1) organs of the original hosting crop (20, 31 and 30 respective Tahiti lime, tree tomato and mango colonies; and 2) organs of the other two crops (9, 18 and 18 respective Tahiti lime, tree tomato and mango colonies), with the aim of determining crossed infection.
The lime flowers were taken from active plantations, packed in air containing plastic bags, and transported in a styrofoam cool box. Once in the laboratory, 320 petals were chosen to be disinfected in a 0.5% sodium hypochlorite (NaOCl) solution for 45 s, and then washed with water and aspersed with a 70% alcohol solution.
After disinfection, they were inoculated with 0.5 mL of a water suspension containing 1x106 isolate spores per mL. For the original host infection tests, a completely random experimental design was applied to measure anthracnose symptom manifestation. The necessary conditions for the development of the disease were provided by placing the petals into a moist chamber (90% RH) at 12±2°C for 12 h.
Regarding tree tomato, 128 green big fruits were taken from active plantations of the 'Common red' variety. After having been packed in individual paper bags and transported to the laboratory in a plastic styrofoam cool box, they were disinfected in a 2.6% NaOCl solution for 30 s, and washed and aspersed with a 70% alcohol solution. Agar discs taken from the Colletotrichum spp. isolates (1x106 spores/mL) were then placed at two points on each fruit. Again, a completely randomized design with four repetitions per isolate was applied to measure anthracnose symptom manifestation. The fruits were incubated in moist chambers (90% RH) at 20±2°C for 30 d.
Finally, 124 mango fruits at first maturation stages were obtained from active plantations of 'hilacha' variety. They were harvested and packed in individual paper bags, which were in turn placed into plastic ones, and then transported to the laboratory in a styrofoam cool box. The disinfection protocol was the same as for mango, with the only difference that NaOCl immersion took one minute, after which the fruits were washed with sterile distilled water and aspersed with the same alcohol solution. Then, except for the spore concentration of the agar discs, which was 1x105 spores/mL, the inoculation process was also the same, and so was the experimental design. Moist chamber incubation took 14 d at 25±2°C, during which anthracnose symptoms were daily registered.
For the crossed infection tests, fruits and petals were collected, transported and disinfected in the same mode described above, but remaining in the moist chambers for 72 h (lime), 96 h (tree tomato), and 28 d (mango). The data from both tests were treated with a SAS® completely randomized design analysis applying the chi-square dependency test (P≤0.5) for the hypotheses that Colletotrichum spp. isolates are capable of reproducing anthracnose symptoms in: 1) their original hosting crops, and 2) the other two studied crops.
Molecular determination of the species
Identification of the species of the pathogen was carried out on 293 of its monosporic isolates, which had been grown in V8® liquid medium at 28°C and 130 rpm for 8 d. Mycelium from such cultures was macerated for DNA extraction and purification, which followed the method proposed by Kelemu et al. (1997; 1999). The solution was adjusted to a final concentration of 20 ng μL-1 for amplification of rDNA ITS region sequences by means of PCR with species specific primers. Using C. gloeosporioides CgInt (5'-GGCCTCCCGCCTCCGGGCGG-3') and C. acutatum CaInt2 (5'-GGGGAAGCCTCTCGCGG-3') specific primers, and in combination with ITS4 universal primer (Freeman et al., 2000; Afanador et al., 2003), such procedure allowed identifying the species.
In determining C. acutatum, amplification took place in a final volume of 20 μL containing Promega® Taq Polymerase buffer, 1.5 mM of MgCl2, 200 μM of each dinucleotide, 0.3 μM of each primer, 1 unit of Taq Polymerase enzyme, and 40 ng of DNA. The amplification profile consisted of an initial cycle of 5 min at 95°C, 40 cycles of 30 s at 95°C, 30 s at 60°C, 1 min at 72°C, and a final extension of 7 min at 72oc.
ITS4 and Cgint primers were used in the identification of C. gloeosporioides. Amplification started with an initial denaturation at 95°C for 5 min, followed by 40 amplification cycles (30 s denaturation at 95°C, 65°C annealing for 30 s, and a 1 min extension at 72°C), plus a final extension cycle of 7 min at 72°C.
In estimating size of the amplified products a kb DNA Ladder marker with molecular weight of 1, ranging between 10,000 and 250 bp was used. Positive controls consisted in DNA taken from the following CIAT isolates: TOM 021 (C. acutatum) (Afanador et al. 2003), and 1613 (C. gloeosporioides) (Kelemu et al., 1997; Kelemu et al., 1999). 120-COL (C. lindemuthianum) (Tamayo et al., 1995) was used as negative control.
Results and discussion
Morphology of colonies and conidia
Most of the isolates coming from citrus crops formed grey and salmon cottony colonies. Tree tomato isolates formed orange colonies with a rather flattened mycelium, and grey or white bottom color. Although mango isolate colonies showed wider variety, abundant green, white, grey or orange cottony mycelium was dominant, sometimes showing luxuriant orange conidial masses with grey or white bottom color (Fig. 1).
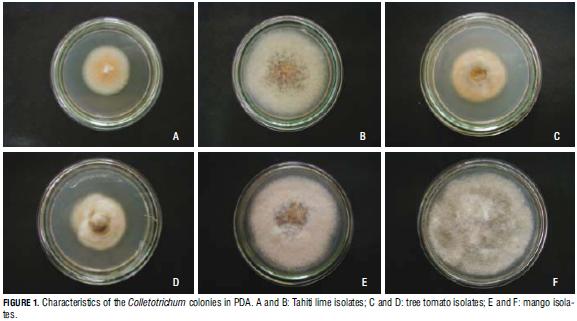
From the results, it can be said that lime and tree tomato isolate colonies showed typical Colletotrichum colors, coinciding with Simmonds' (1965) and Von Arx's (1957) descriptions of C. acutatum. In turn, mango isolate colonies corresponded to descriptions of C. gloeosporioides published by Sutton (1980) and Baxter (1983). However, the morphology of Colletotrichum colonies varies within and among groups, depending on culture medium, substrate and temperature, among other factors (Contreras, 2006).
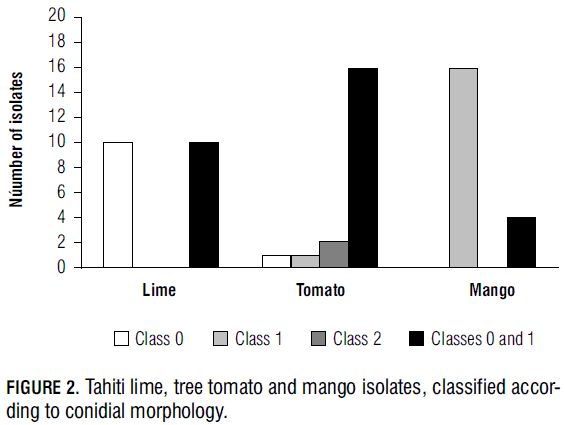
The analysis of conidial morphology showed that one single colony may contain two different types of spores. Some of the Tahiti lime isolates gave rise to classes 0 and 1 conidia. Isolates coming from tree tomato showed several types (0, 1 and 2; or 0 and 1). On the other hand, most of the 20 mango isolate colonies grew class 1 conidia, and a few of them exhibited classes 0 and 1 (Fig. 2).
Mango isolates stood out by exhibiting bigger average conidia (21.5 μm), whereas tree tomato and lime ones proved to be smaller. Conglomerate data analysis of these features showed great variability.
PDA grown hyphae were observed to be hyaline, with defined septa, sometimes exhibiting cytoplasmic contents, and sometimes intercellular spaces. Microscopic characterization of the micro-cultured hyphae of the different fruit crop isolates showed no significant differences in the Anova test (P≤0.5). This is, therefore, a character of little use in differentiating these species.
Fungicide susceptibility
The results obtained through the benomyl and copper hydroxide tests allowed identifying the 100 mango isolates as C. gloeosporioides (Tab. 3), due to their high growth rate, grey and salmon color in the selective medium, and susceptibility to benomyl (Agostini and Timmer, 1992). In turn, the 100 tree tomato isolates were identified as C. acutatum for presenting tolerance to benomyl, low growth rate after 72 h of incubation, and orange color in the selective medium (Tab. 3). Eighty seven of the Tahiti lime isolates were found to be benomyl tolerant, and therefore classified as SGO (C. acutatum). The resting 13 ones were identified as C. gloeosporioides due to their benomyl susceptibility, high growth rate, and orange or grey color in the selective medium (Tab. 3).
It can be seen how the mango isolates have a different growth pattern from the Tahiti lime and tree tomato ones, which were similar in either medium. This indicates the possibility that the population of C. gloeosporioides is inherently variable (Dodd et al., 1991; Estrada et al., 2000; Afanador et al., 2003).
These results can also be used to infer isolate virulence. Abang (2003) found category SGG fungi to be the most aggressive ones, due to the degrading enzymes they posses, which are capable of destroying the cell membranes of young plant tissues. Notwithstanding, other studies point at C. acutatum as the most infectious species, because of its resistance to several fungicides (Peres et al., 2002a).
Pathogenicity
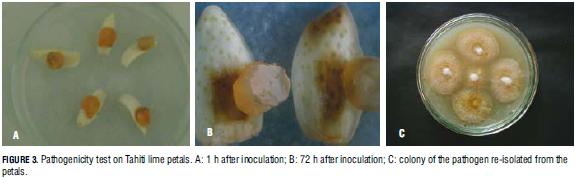
Out of the 20 evaluated Tahiti lime isolates, those identified as C. acutatum provoked anthracnose symptoms in 188 out of 320 inoculated Tahiti lime petals (58.8%) 72 h after inoculation (Fig. 3). The pathogen was recovered and re-isolated from the infected tissue, thus proving that this species is the anthracnose infectious agent in this crop. Regarding tree tomato, the first symptoms of the disease caused by this crop's isolates were seen on the 13th day after inoculation; on day 30th, the infection had reached medium incidence (Fig. 4). Again, the original pathogen was re-isolated from the infected fruits.

In the case of mangoza (Fig. 5), out of the 30 isolates of the pathogen tested for original host infection, 19 were able to produce the first symptoms on 22 fruits on the fourth day after inoculation. By day 14th, the incidence of the disease had reached 100% of the fruit; on days 4th and 5th, the pathogen was grown again from infected fruit tissue samples in PDA medium. These results allowed determining that the evaluated isolates are highly infectious (P≤0.05).

Finally, the crossed infection tests gave negative results, as no symptoms of the disease were detected in organs inoculated with isolates taken from other crops (Tab. 4).
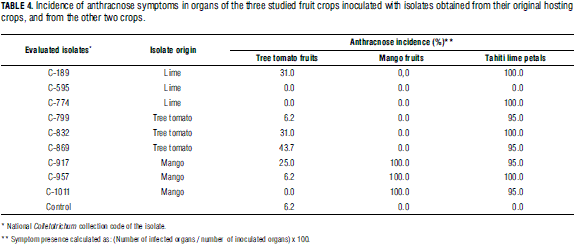
Thus, the pathogens can be said to be host specific, which is confirmed by studies on the genetic diversity of the same collection analyzed in this work (Osorio et al., unpublished results).
Molecular determination of the species
Out of the 293 isolates amplified in the present study, 182 were identified as C. acutatum, and 111 as C. gloeosporioides (Fig. 6). All the tree tomato isolates were identified as C. acutatum, and all the mango ones as C. gloeosporioides. Out of the 93 Tahiti lime isolates, 83 corresponded to C. acutatum, and 10 to C. gloeosporioides. These results confirm those obtained with the morphological analysis and fungicide sensitivity tests. Similarly, Afanador et al. (2003) and Timmer and Brown (2000) characterized Colletotrichum spp. isolates obtained from different fruit crops, identifying C. acutatum as the anthracnose infectious agent in lime and tree tomato. Nevertheless, anthracnose affected plants of the latter crop infected by C. gloeosporioides have also been reported (Aranzazu and Rondón, 2001). Both species can be found in citrus, but only C. acutatum is responsible for premature fruit drop. The other species is just an associated saprophyte (Timmer and Brown, 2000).

Conclusions
1. The morphological, molecular, pathogenicity and fungicide sensitivity analyses conducted on the pathogens that determine anthracnose in the three studied crops allowed identifying them as Colletotrichum spp.
2. In Colombia, anthracnose in Tahiti lime and tree tomato is caused by C. acutatum, while in mango it is determined by C. gloeosporioides. The three infectious agents proved to be host specific, as far as no crossed infection among crops was seen.
3. Given that the therapeutic management tools and strategies currently in use for controlling the pathogens in Colombia have not been sufficiently effective in reducing the impact of the disease, its affecting productivity or fungicide application levels, the newly acquired knowledge contributed by the present research study allows developing alternative disease management protocols. This is due to the fact that the two studied Colletotrichum species present different reactions to control measures, among which we can count fungicides. The alternative scheme would focus on preventive control strategies, which, systematically applied to the plantations, would be in condition to reduce the incidence of the disease to more manageable levels, with less intense use of agrochemicals, and lower costs for the producer.
___________________3 In the original in spanish "departamentos", which are the administrative units of the country. Translator's note.
4 Tibaitata Research Center. Translator's note.
Literature cited
Abang, M., S. Winter, H. Mignouna, K. Green, and R. Asiedu. 2003. Molecular taxonomic, epidemiological and population genetic approaches to understanding yam anthracnose disease. Afr. J. Biotechnol. 2(12), 486-496. [ Links ]
Andrade, E., C. Uesugi, B. Ueno, and M. Ferreira. 2007. Caracterização morfocultural e molecular de isolados de Colletotrichum gloeosporioides patogênicos ao mamoeiro. Fitopatol. Bras. 32, 21-31. [ Links ]
Afanador, L., D. Minz, M. Maymon, and S. Freeman. 2003. Characterization of Colletotrichum isolates from tamarillo, pasiflora and mango in Colombia and identification of a unique species from genus. Phytopathol. 93(5), 579-587. [ Links ]
Agostini, J.P. and L.W. Timmer. 1992. Selective isolation procedures for differentiation of two strains of Colletotrichum gloeosporioides from citrus. Plant Dis. 76, 1176-1178. [ Links ]
Aranzazu, L. and G. Rondón. 2001. Manejo productivo del cultivo de tomate de árbol y de la Antracnosis. Bol. divulgativo, Corpoica, Bogota. [ Links ]
Arauz, L.F. 2000. Mango Anthracnose: economic impact and current options for integrated management. Plant Dis. 84(6), 600-611. [ Links ]
Barnet, H.L. and B. Barry. 2003. Ilustrated genera of imperfect fungi. 4th ed. APS Press, St. Paul, MN. pp. 188-191. [ Links ]
Baxter, A., G.C.A. Westhuizen, and A. Eicker. 1983. Morphology and taxonomy of South African isolates of Colletotrichum. S. Afr. J. Bot. 2, 259-289. [ Links ]
Brown, A.E., S. Sreenivasaprasad, and L.W. Timmer. 1996. Molecular characterization of slow-growing orange and key lime Anthracnose strains of Colletotrichum from citrus as C. acutatum. Phytopathol. 86, 523-527. [ Links ]
Burdon, J. and J. Silk. 1997. Sources and patterns of diversity in plant - pathogenic fungi. Phytopathol. 87(7), 664-699. [ Links ]
Contreras, C. 2006. Caracterización y pruebas de patogenicidad cruzada entre aislamientos de Colletotrichum spp. obtenidos de frutos de lulo, tomate de árbol, granadilla, mango y tallos de mora con síntomas de antracnosis. Undergraduate thesis. Facultad de Ciencias Básicas, Pontificia Universidad Javeriana. Bogotá, Colombia. [ Links ]
Dodd, J.C., R. Bugante, I. Koomen, and P. Jeffries. 1991. Pre-and post-harvest control of mango Anthracnose in the Philippines. Plant Pathol. 40, 576-583. [ Links ]
Estrada, A., J. Dodd, and P. Jeffries. 2000. Effect of humidity and temperature on conidial germination and appressorium development of two Philippine isolates of the mango Anthracnose pathogen Colletotrichum gloeosporioides. Plant Pathol. 49(5), 608-618. [ Links ]
Freeman, S. and T. Katan. 1997. Identification of Colletotrichum species responsible for Anthracnose and root necrosis of strawberry in Israel. Phytopathol. 87, 516-521. [ Links ]
Freeman, S., T. Katan, and E. Shabi. 1998. Characterization of Colletotrichum species responsible for Anthracnose diseases of various fruits. Plant Dis. 82, 596-605. [ Links ]
Freeman, S., D. Minz, E. Jurkevitch, M. Maymon, and E. Shabi. 2000. Molecular analysis of Colletotrichum species from almond and other fruits. Phytopathol. 90, 608-614. [ Links ]
Freeman, S., D. Minz, M. Maymon, and A. Zveibil. 2001. Genetic diversity within Colletotrichum acutatum sensu Simmonds. Phytopathol. 91, 586-592. [ Links ]
Kelemu, S., J.L. Badel, C.X. Moreno, J. Miles, W.S. Chakraborty, C.D. Fernándes, and M.J. Charchar. 1997. Biodiversity, epidemiology and virulence of Colletotrichum gloeosporioides. I. Genetic and pathogenic diversity in Colletotrichum gloeosporioides isolates from Stylosanthes guianensis. Trop. Grasslands 31, 387-392. [ Links ]
Kelemu, S., D. Skinner, J. Badel, C. Moreno, C. Rodríguez, C. Fernándes, M. Charchar, and S. Chakraborty. 1999. Genetic diversity in South American Colletotrichum gloeosporioides isolates from Stylosanthes guianensis, a tropical forage legume. Eur. J. Plant Pathol. 105, 261-272. [ Links ]
Páez, A.R. 1995. Uso de variedades tolerantes: alternativa para el manejo de Antracnosis (Colletotrichum gloeosporioides) en mango (Mangifera indica). ASCOLFI-Informa 21, 36-39. [ Links ]
Peres, N., N. Souza, S. Zitko, and L. Timmer. 2002a. Activity of benomyl for control of postbloom fruit drop of citrus caused by Colletotrichum acutatum. Plant Dis. 86, 620-624. [ Links ]
Peres, N., E. Kuramae, M. Dias, and N. Souza. 2002b. Identification and characterization of Colletotrichum spp. affecting fruit after harvest in Brazil. J. Phytopathol. 150, 128-134. [ Links ]
Peres, N.A., L.W. Timmer, J.E. Adaskaveg, and J.C. Corell. 2005. Lifestyles of Colletotrichum acutatum. Plant Dis. 41, 84-95. [ Links ]
Sanabria, A. 2007. Caracterización de la diversidad genética de Colletotrichum spp. como agente causante de Antracnosis en limón, tomate de árbol y mango. M.Sc. thesis. Instituto de Biotecnología, Facultad de Ciencias, Universidad Nacional de Colombia, Bogota. [ Links ]
Simmonds, J. 1965. A study of the species of Colletotrichum causing ripe fruit rots in Queensland. Queensland J. Agr. Sci. 22, 437-459. [ Links ]
Sutton, B. 1980. The coleomycetes. Fungi imperfecti with pycnidia, acervula and stromata commonwealth. Mycological Institute. Kew, UK. [ Links ]
Sutton, B.C. 1992. The genus Glomerella and its anamorph Colletotrichum. pp. 1-27. In: Bailey, J.A. and M.J. Jeger (eds.). Colletotrichum: biology, pathology and control. CAB, Wallingford, UK. [ Links ]
Tamayo, P.J. 2001. Principales enfermedades del tomate de árbol, la mora y el lulo en Colombia. Corpoica, Centro de Investigación La Selva. Bol. Tec. 12, Rionegro, Colombia. [ Links ]
Tamayo, P.J., M.M. Otoya, and M.A. Pastor-Corrales. 1995. Diversidad de razas de Colletotrichum lindemuthianum, el patógeno de la Antracnosis de fríjol en Rionegro, Antioquia. Fitopatol. Colomb. 19(1), 1-6. [ Links ]
Timmer, L.W. and G.E. Brown. 2000. Biology and control of Anthracnose diseases of citrus. pp. 317-336. In: Freeman, S., D. Prusky, and M. Dickman (eds.). Colletotrichum, host specificity, pathology, and host pathogen interaction. APS Press, St. Paul, MN. [ Links ]
Von Arx, J.A. 1957. Die Arten der Gattung Colletotrichum Cda. Phytopathol. Zeitschrift 29, 413-468. [ Links ]
Whitelaw-Weckert, M., S. Curtin, R. Huang, C. Steel, C. Blanchard, and P. Roffey. 2007. Phylogenetic relationships and pathogenicity of Colletotrichum acutatum isolates from grape in subtropical Australia. Plant Pathol. 56, 448-463. [ Links ]

