










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.27 n.2 Bogotá maio/ago. 2009
Johana Carolina Soto S.1 y Juan José Filgueira D.1, 2
1 Biología Aplicada, Facultad de Ciencias, Universidad Militar Nueva Granada, Bogotá (Colombia).
2 Autor de correspondencia. jfilgdu@gmail.com
Fecha de recepción: 17 de marzo de 2009. Aceptado para publicación: 2 de julio de 2009
RESUMEN
El mildeo velloso de la rosa, producido por Peronospora sparsa, es una de las enfermedades más limitantes y costosas en los cultivos comerciales de rosa en Colombia. La estrecha relación de la enfermedad con las condiciones climáticas ha llevado a realizar estudios del parásito en condiciones microclimáticas controladas. Esta investigación buscó ahondar en el conocimiento de la biología del mildeo velloso de la rosa, evaluando para ello el efecto del fotoperiodo y la intensidad lumínica sobre la esporulación del parásito cultivado en folíolos de rosa en cámara húmeda bajo condiciones ambientales controladas. Se evaluaron los siguientes fotoperiodos: 24 horas bajo una intensidad lumínica de 1.000 lux; 8, 12 y 24 horas luz a 790 lux; 8, 16 y 24 horas a 560 lux; 12 horas a 120 lux, y finalmente 0 horas luz (oscuridad total). La esporulación se vio favorecida por fotoperiodos de 8 a 16 horas a 560 lux, y presentó un alto desempeño con periodos de 12 horas a 790 lux. La exposición continua a la luz demostró tener un efecto inhibidor sobre la esporulación de P. sparsa. Así mismo, determinó la producción de esporangios con características atípicas, como bajo contenido citoplasmático y menor tamaño.
Palabras clave: mildeo velloso, rosa, peronosporales.
ABSTRACT
Downy mildew, caused by Peronospora sparsa, is one of the most restrictive and costly diseases in Colombian rose plantations. The close relation of the disease to climatic conditions determines the need to study the parasite under controlled microclimatic conditions. The objective of the present research was to improve the understanding of the biology of Peronospora sparsa, in terms of the effect of photoperiod and light intensity on the sporulation of the parasite, which was grown on rose leaves under controlled environmental conditions in moist chamber. The evaluated photoperiods were: 24 hours under a light intensity of 1,000 lx; 8, 12 and 24 hours at 790 lx; 8, 16 and 24 hours - 560 lx; 12 hours - 120 lx, and finally complete darkness. Sporulation was favored by 8 to 16 hour photoperiods at 560 lx, and strongly enhanced by 12 hour periods at 790 lx. Permanent light exposure not only showed an inhibitory effect on sporulation, but also determined the production of atypical sporangia (little cytoplasm and small size).
Key words: downy mildew, rose, peronosporales.
Introducción
El mildeo velloso de la rosa, causado por el patógeno Peronospora sparsa Berkeley, es una enfermedad que en los últimos años ha venido causando grandes pérdidas económicas en los cultivos de rosa comercial de la Sabana de Bogotá, convirtiéndose en el principal problema sanitario de este cultivo (Arbeláez, 1999; Gómez y Arbeláez, 2005).
El primer registro de esta enfermedad en la rosa lo hizo M.J. Berkeley (Horst, 1983) en 1882 en un invernadero de Inglaterra, quien describió al agente causal tal como hoy en día se conoce. El parásito produce síntomas en hojas, tallos, pedúnculos, cálices y pétalos, aunque la infección generalmente se limita a tejidos jóvenes. Es frecuente observar el desarrollo de manchas irregulares de color rojizo púrpura a pardo-oscuro, rodeadas de un halo clorótico en el haz de las hojas y sobre el envés micelio de color marrón claro con abundante producción de esporangióforos y esporangios (Ayala et al., 2008). El desarrollo de la enfermedad es favorecido por humedades relativas altas y temperaturas bajas (Wheeler, 1981).
Los parásitos que producen los mildeos vellosos son fitopatógenos muy importantes en varios cultivos en todo el mundo. A mediados de los años noventa del siglo pasado se dio una gran expansión geográfica de especies como el mildeo velloso, y con esto se aumentaron las investigaciones en el tema del patógeno y estudios para su control. Los trabajos comenzaron con Peronospora destructor en cebolla, Peronospora tabacina en tabaco y Peronospora viticola en uva (Virnnot-Bourgin, 1981).
La enfermedad presenta una estrecha relación entre el comportamiento biológico del patógeno y las variables climáticas; sin embargo, como este parásito no se presenta en invernaderos climatizados, las investigaciones realizadas en países nórdicos y la información que hasta hace unos años se conocía sobre la biología del patógeno es escasa (De Vis, 1999). En Colombia la investigación con el mildeo velloso de la rosa se ha venido desarrollando desde hace algunos años a través de universidades y centros de investigación (Gómez, 2004; Gómez y Arbeláez, 2004; Urrea y Arbeláez, 2004; Gómez y Arbeláez, 2005), pero el conocimiento sobre la relación del patógeno y las condiciones ambientales aún es limitado (Gómez y Arbeláez, 2005); de allí que la necesidad apremiante del sector floricultor es conocer mejor al patógeno y su biología, para así llegar a tener un manejo más eficiente de la enfermedad, integrando la manipulación de variables climáticas y demás prácticas culturales.
En el mildeo velloso de la cebolla se ha estudiado el papel de la luz, conjugado con la temperatura y la humedad; observa que existe una dependencia de periodos de luz y oscuridad en la esporulación de Peronospora destructor: el patógeno esporula cuando hojas de cebollas infectadas han sido expuestas a periodos de luz seguido de periodos de oscuridad (Hildebran y Sutton, 1984).
Para el estudio de patologías como el mildeo velloso, que presentan una estrecha relación con condiciones microclimáticas, es de gran utilidad el montaje de sistemas en donde sea posible generar y controlar un ambiente uniforme, reproducible y con una mayor precisión que en otras locaciones de investigación, para así observar y evaluar respuestas diferenciales en el patógeno ante variaciones climáticas. Así, el presente trabajo pretende profundizar en el conocimiento del comportamiento biológico del agente causante del mildeo velloso de la rosa, evaluando el efecto del fotoperiodo y de la intensidad lumínica en la esporulación de Peronospora sparsa en foliolos de rosa en cámara húmeda, en condiciones ambientales controladas y reproducibles.
Materiales y métodos
Sistema de condiciones ambientales controladas
Esta investigación se desarrolló en el Laboratorio de Fitopatología de la Facultad de Ciencias de la Universidad Militar Nueva Granada en el campus universitario ubicado en Cajicá, Cundinamarca, en donde se construyó un sistema de condiciones ambientales controladas, el cual consta de un cuarto climatizado con dimensiones de 288,5 cm de ancho, 190 cm de largo y 250,5 cm de alto, en cuyo interior hay una cámara de acrílico de 100 cm de ancho, 150 cm de largo y 100 cm de alto (Fig. 1). Como fuentes de luz, se tienen seis lámparas situadas en una base móvil ascendente y descendente de luz fluorescente de 40 W, todas conectadas a un temporizador para permitir la manipulación de fotoperiodo (Fig. 1).

Las paredes del cuarto están totalmente cubiertas con láminas de icopor de 2,5 cm de ancho, recubiertas con Duralfoil®. La temperatura puede ser regulada con un sistema de aire acondicionado, o bien aumentando la temperatura con un calentador de ambiente. La puerta de acceso al cuarto es completamente hermética. La humedad relativa dentro de las cámaras es controlada por un sistema de humidificación por ultrasonido conectado al interior de la cámara de acrílico grande por medio de una flauta con escapes cada 10 cm, que cuenta con válvulas para la regulación de la micronebulización (Fig. 1). Para la medición de las variables de temperatura y porcentaje de humedad relativa se cuenta con un Data Logger WatchDog modelo 450 (Plainfiled, IL), un higrotermógrafo Oakton modelo 08368-41 (Vernon Hills, IL) con registro para una semana e higrotermómetros de medición instantánea, máximas y mínimas Fisher modelo 11-661-7D (Sindelfingen, Alemania).
La temperatura dentro del cuarto y de las cámaras puede manejarse desde temperaturas bajas, 5°C, hasta un máximo de 33°C; la humedad relativa dentro de las cámaras puede ser regulada desde 40% hasta el 99%, alcanzando niveles de saturación para mantener una película de agua sobre superficies foliares, si así se requiere; humedades relativas por debajo del 55% se alcanzan por deshumidificación con silicagel. En cuanto al fotoperiodo (h luz por día), este puede programarse para cualquier duración de minutos, horas o bien luz continua. La intensidad lumínica puede ser medida a través de un exposímetro de luz, y esta varía dentro de las cámaras, dependiendo de la distancia en la que se ubique la base móvil de las lámparas y del número de estas encendidas. Los niveles de iluminación, medidos en lux (lumen/m2; lx), se pueden establecer en la cámara externa desde 0,7 lx hasta 1.000 lx y en la cámara interna desde 0,7 lx hasta 970 lx.
Producción y mantenimiento de inóculo en cámara húmeda
El inóculo inicial provino de folíolos infectados en campo, de plantas de rosa de la variedad Charlotte. Se obtuvieron esporangios de estas esporulaciones por barrido con un pincel estéril, el cual fue lavado en un vaso de precipitado en 20 mL de agua destilada estéril (ADE); posteriormente se filtró la suspensión obtenida empleando velo suizo estéril, y mediante un hemocitómetro se cuantificó el número de esporangios/mL.
Para el mantenimiento del inóculo inicial y posterior uso en los diferentes ensayos, cada semana se recolectaron folíolos sanos y tiernos de aproximadamente 15 d de edad del tercio superior de plantas de la variedad Charlotte, susceptible a mildeo velloso. Los folíolos fueron sometidos a los siguientes lavados: el primero con agua destilada estéril (ADE) más hipoclorito de sodio al 5%, durante 3 min; el segundo lavado con ADE más Tween 80 al 0,1% durante 3 min; por último, tres lavados con ADE. Posteriormente los folíolos fueron secados con toallas absorbentes estériles y se colocaron cuatro por caja de Petri, con el envés expuesto sobre hojas de papel filtro estéril completamente húmedas en 3 mL de ADE en cámara húmeda.
En cada folíolo y a cada lado de la vena central se depositaron cuatro gotas de la suspensión de esporangios en una concentración de 20.000 esporangios/mL. Transcurridas 24 h, se secaron los folíolos y se dejaron en cámara húmeda en condiciones de temperatura y luminosidad ambiente durante 120 h aproximadamente, tiempo en el cual se observan las manchas típicas de mildeo velloso y la esporulación (Fig. 2).

Estudio del efecto del fotoperiodo e intensidad lumínica
Dentro de la cámara de acrílico se ubicaron cinco cajas de Petri con montaje de cámara húmeda para producción de mildeo velloso, como se describió anteriormente. El aire acondicionado estuvo encendido en funcionamiento automático y distribución uniforme del aire, manteniendo una temperatura entre 19±1°C dentro de la cámara para todos los ensayos realizados. Para los ensayos de variación de fotoperiodo, la base móvil de lámparas estuvo ubicada a la distancia mínima sobre la cámara externa, a una distancia de 12 cm, con las seis lámparas encendidas, dando una intensidad lumínica de 1.000 lx, evaluando la producción de esporangios en fotoperiodo de 24 h luz d. Se realizaron tres ensayos de intensidad lumínica a 790 lx en donde solo las seis lámparas estuvieron encendidas y ubicadas a la altura mínima, evaluando fotoperiodos de 8, 12 y 24 h luz.
Para las evaluaciones de fotoperiodo 8, 16 y 24 h luz a intensidad lumínica de 560 lx, la base móvil de las lámparas se mantuvo a la distancia mínima sobre la cámara externa y con tres lámparas encendidas. Se realizaron ensayos para evaluar la intensidad lumínica mínima, en los que la base móvil de lámparas estuvo ubicada a la distancia máxima de 77 cm sobre la cámara externa con una de las seis lámparas encendidas, dando una intensidad lumínica de 120 lx, con fotoperiodo 12 h luz. Por último se evaluó la respuesta de P. sparsa ante oscuridad total. Simultáneamente, a cada uno de los ensayos se montó un control de cinco cajas de Petri con montaje de cámara húmeda para producción de mildeo velloso en condiciones de salón, estos ensayos se realizaron tres veces para corroborar los datos.
La comparación y evaluación del efecto se realizaron a las 120 h del ensayo, recolectando los esporangios producidos durante este tiempo del total de los folíolos de las cajas de Petri, por barrido con pincel en 20 mL de ADE, tanto para el ensayo como para el control, de acuerdo con la concentración del inóculo, medido con hemocitómetro, germinación de esporangios en condiciones de salón, medido en porcentaje de viabilidad, y por último, observación microscópica de la morfología. Dentro de este último parámetro fueron tomados como morfología típica esporangios ovalados, con contenido celular completo, con paredes enteras y de tamaño normal, dimensiones entre 17-22 μm x 14-18 μm, tal como lo describe Wheeler (1981).
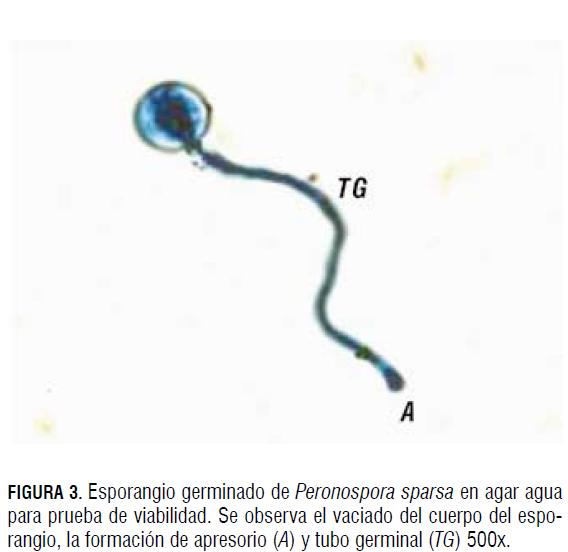
Por otro lado, una morfología atípica fue tomada como esporangios pequeños (EP), con bajo contenido citoplasmático (BCC) o paredes arrugadas (PA). El porcentaje de viabilidad se midió a través de pruebas de germinación en agar agua 15 g L-1, colocando una capa delgada de agar en un portaobjetos de vidrio, en donde se distribuyeron 300 μL de la suspensión de esporangios, ajustada a la misma concentración de esporangios/mL. Los portaobjetos se dejaron en condiciones de salón durante 24 h en cámara húmeda. Transcurrido este tiempo, se realizaron observaciones en el microscopio, determinando el porcentaje de germinación. Se consideró un esporangio germinado a aquel cuya longitud del tubo germinal fuera el doble del diámetro del esporangio (Fig. 3). Tanto los montajes dentro de la cámara de acrílico como los controles fueron realizados por duplicado y en muchos casos por triplicado para corroborar el resultado, tomando el mismo inóculo y concentración inicial de 20.000 esporangios/mL, el mismo número de folíolos, así como su tamaño y edad.
Resultados y discusión
La esporulación de P. sparsa se presentó tanto en los ensayos bajo oscuridad total, fotoperiodo y luz constante, de manera diferencial, mostrando que la duración de la luz tiene un efecto estimulante o inhibidor, según el caso, en el desarrollo y la esporulación de P. sparsa. El sistema climatizado diseñado e implementado en esta investigación permitió generar y controlar un ambiente uniforme, reproducible y con alta precisión, en donde fue posible observar y evaluar respuestas diferenciales en las esporulaciones obtenidas en los ensayos planteados.
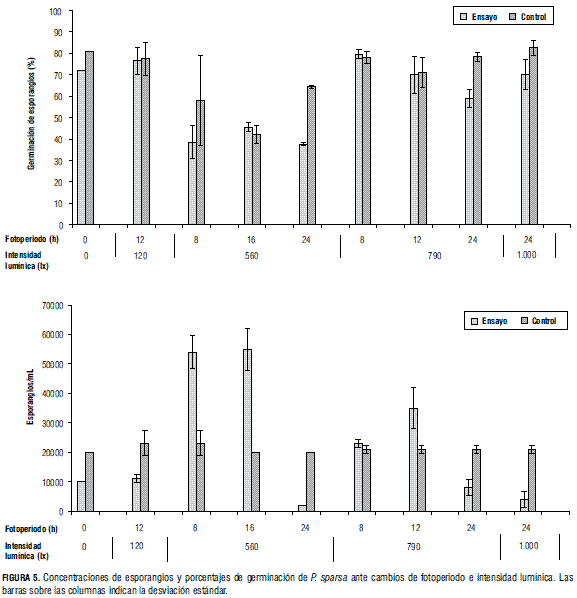
La esporulación en condiciones de oscuridad total (fotoperiodo 0 h luz) produjo una de las concentraciones de esporangios más bajas obtenidas en las diferentes evaluaciones; además, fue muy similar, en términos de concentración de esporulación y porcentaje de viabilidad, a la presentada en los ensayos de baja intensidad lumínica a 120 lx con fotoperíodo 12 h, mostrando que para la esporulación tiene mayor importancia la cantidad de luz, medida en intensidad lumínica presente, que el tiempo de duración de la misma. En cuanto a la morfología, en este ensayo se observaron esporangios con tamaños inferiores a las dimensiones registradas para P. sparsa (Fig. 4B).
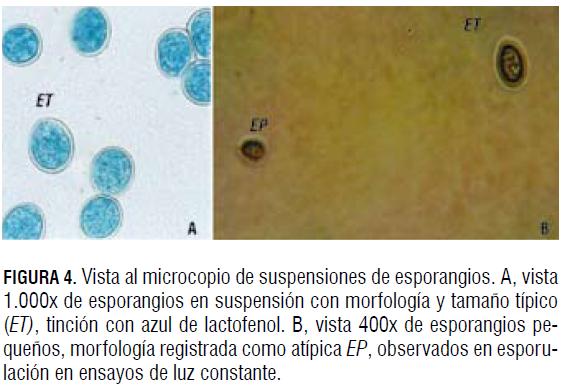
La concentración de esporangios producida ante exposición de 120 lx y fotoperiodo de 12 h luz presentó una disminución considerable de la concentración de esporangios, en comparación con la obtenida con el mismo fotoperíodo pero con intensidad lumínica de 790 lx; sin embargo, el porcentaje de viabilidad fue mayor (Fig. 5B). La morfología de los esporangios obtenidos en el ensayo fue típica (Fig. 4A). Estos resultados muestran que para el caso de P. sparsa que ataca la rosa en Colombia, la luz de baja intensidad no inhibe la esporulación.
El efecto de la luz y otras condiciones medioambientales en la esporulación y desarrollo de los mildeos vellosos han sido anteriormente estudiados, en un comienzo por Weston (1923, 1924) y posteriormente por Yarwood (1937), quienes fueron los primeros en registrar que la esporulación de los agentes causantes de mildeos vellosos ocurre bajo condiciones de oscuridad y humedad, seguidas de períodos de luz, y que en condiciones de luz, oscuridad o alta humedad, la esporulación es muy poca, o no se presenta (Populer, 1981). Así mismo, en P. destructor en cebolla se ha registrado que existe un efecto en ciclos de luz y oscuridad en la esporulación del patógeno (Hildebran y Sutton, 1984).
En las evaluaciones de intensidad lumínica 560 lx, al incrementarse el fotoperiodo, h luz día, la esporulación de P. sparsa aumentó, presentándose la máxima concentración de esporangios/mL con respecto al control en el ensayo de fotoperiodo 16 h luz, seguida del ensayo de 8 h luz. La menor concentración de esporangios/mL se presentó ante exposición de luz constante (fotoperiodo 24 h luz). Así mismo, el mayor porcentaje de germinación de los esporangios se presentó en el ensayo 16 h luz. La concentración de la esporulación de los ensayos control se mantuvo constante, alrededor de 20.000 esporangios/mL (Fig. 5A y 5B).
La morfología de los esporangios fue típica en todos los controles y en los ensayos de fotoperiodo 8 y 16 h luz (Fig. 4A); sin embargo, en alrededor del 10% de los esporangios, se observaron características atípicas como bajo contenido citoplasmático y esporangios pequeños en las suspensiones provenientes del ensayo 24 h luz (Fig. 4B).
Ante intensidad lumínica 790 lx, las mayores concentraciones de esporangios se obtuvieron con fotoperiodo 12 h luz; no obstante, la concentración fue inferior a la alcanzada con fotoperiodo 16 h luz e intensidad lumínica a 560 lx. Ante exposición de luz constante, se observó una disminución significativa en la concentración de esporangios/mL. Este resultado muestra que se necesita intensidad de luz por encima de 790 lx o una duración de luz mayor a 16 h para reducir la esporulación de P. sparsa en folíolos de rosa.
La esporulación obtenida en el ensayo de fotoperiodo 8 h luz día e intensidad lumínica 790 lx presentó un comportamiento intermedio entre las esporulaciones ante 12 y 24 h luz día con la misma intensidad lumínica. El mayor porcentaje de germinación en la prueba de viabilidad se presentó en fotoperiodo 8 h luz (Fig. 5B). La morfología de los esporangios obtenidos de los ensayos 8 y 12 h luz fue típica, pero ante luz constante (fotoperiodo 24 h luz) fue atípica, tal como se presentó con intensidad lumínica de 560 lx (Fig. 4B).
Estudios en Pseudoperonospora cubensis muestran que el potencial de esporulación es directamente proporcional a la intensidad lumínica, y es dependiente de la temperatura, exhibiendo un comportamiento similar al encontrado en este estudio. Así, el incremento relativo de producción de esporangios por unidad de área foliar a 15 y 20°C es superior que a 25 y 30°C; además, fotoperiodos largos fueron asociados a un mejor desarrollo del patógeno. Sin embargo, con iluminación continua, el potencial de esporulación fue menor que el obtenido en fotoperiodos de 20 h luz (Cohen y Rotem, 1971).
En el ensayo de intensidad lumínica de 1.000 lx y fotoperiodo 24 h luz se evaluó la condición máxima, tanto en fotoperiodo como en intensidad lumínica. Estas condiciones redujeron la esporulación en los folíolos infectados. La exposición de luz constante de igual manera afectó, disminuyendo la esporulación en los ensayos de intensidad lumínica 560 lx y 790 lx, presentando esporulaciones menores o iguales a 6.000 esporangios/mL.
La exposición de luz continua ha mostrado ser inhibitoria, parcial o totalmente, para la producción de esporangios en especies como Peronospora tabacina (Cruickshank, 1963), Peronospora pisi (Pegg y Mence, 1970), Peronosclerospora sorghi (Schmitt y Freytag, 1974) y Bremia lactuceae (Raffray y Sequeiro, 1971).
Los resultados obtenidos aquí también han sido observados en el mildeo velloso de las cucurbitáceas, en donde se describió el efecto de la intensidad lumínica y del fotoperiodo en el desarrollo del patógeno, que el desarrollo de la enfermedad a 20°C, medido en porcentaje de área foliar infectada, aumenta al ser incrementada la intensidad lumínica durante la incubación. Además, bajas intensidades lumínicas reducen tanto el número como el tamaño de las lesiones (Cohen, 1981).
Tal como se evaluó la viabilidad en este estudio, medir la germinación de esporangios de P. sparsa en superficie de agar es una práctica recurrente (Cruickshank, 1961). La viabilidad medida en porcentaje de germinación de los ensayos de fotoperiodo e intensidad lumínica fue, en su mayoría, superior en las suspensiones de esporangios provenientes de los controles, exceptuando aquellos provenientes del ensayo de fotoperiodo 16 h luz día a 560 lx, el cual estuvo por encima del control en promedio tan solo un 4%. Por otro lado, la viabilidad de los esporangios obtenidos en el ensayo de fotoperiodo 24 h luz día a 560 lx tuvo el menor comportamiento de los fotoperiodos evaluados, con respecto a su control (Fig. 5B).
Giraldo et al. (2002) registraron que la iluminación es un factor fundamental en el desarrollo de la infección de P. sparsa, que estimula o inhibe su esporulación, y que al igual que la temperatura, el fotoperiodo y 20°C estimulan la esporulación del parasito. Por tanto, los resultados obtenidos en esta investigación indican que un fotoperiodo de 16 h a 560 lx en las condiciones de temperatura entre 18 y 20°C y humedad relativa presentadas dentro de la cámara de acrílico del sistema climatizado garantiza un incremento importante en la esporulación de P. sparsa en folíolos de rosa.
El efecto de la luz sobre la morfología de los esporangios se ha observado en otros parásitos causantes de mildeos vellosos, como es el caso de P. destructor, en donde se concluyó que la luz afecta el desarrollo de los esporangios, cuando está presente en estados tempranos de la esporulación, produciendo deformidades en esporangios jóvenes (Hildelbrand y Sutton, 1984).
El hecho de que en presencia de luz constante se produzcan, además de baja concentración, esporangios con morfología atípica, de menor tamaño, bajo contenido citoplasmático y baja viabilidad, indica que la presencia de luz, además de desempeñar un papel importante dentro del proceso reproductivo de P. sparsa, puede afectar el desarrollo de esporangios y muy seguramente su capacidad de infección.
Conclusiones
Según los resultados de estos ensayos, el fotoperiodo y la intensidad lumínica son factores esenciales en el desarrollo de P. sparsa, viéndose favorecida la esporulación del parásito por 8 a 16 h de exposición a luz, con un alto desempeño a 12 h de luz, e intensidades lumínicas iguales o mayores a 560 lx, condiciones lumínicas encontradas de un invernadero típico y comercial de rosa. Este hecho, sumado a las condiciones de temperatura y humedad presentadas en nuestros invernaderos, hacen que los cultivos de rosa de la Sabana de Bogotá sean vulnerables al ataque por mildeo velloso.
Finalmente, de acuerdo con lo anterior se recomienda iluminar los invernaderos durante las noches que tienen las más bajas temperaturas en conjunto con la más alta humedad relativa, condiciones óptimas para la esporulación del parásito, con luz que tenga intensidades medibles en la parte baja de la planta de rosa de no menos de 560 lux en los cultivos comerciales de rosa.
Agradecimientos
A la Universidad Militar Nueva Granada y al Instituto Colombiano de Ciencia y Desarrollo Francisco José de Caldas, (Colciencias), por la financiación de la investigación a través del proyecto CIAS 065 y del programa de Jóvenes Investigadores e Innovadores 2006-2007.
Literatura citada
Arbeláez, G. 1999. El mildeo velloso del rosal ocasionado por Peronospora sparsa Berkeley. Acopaflor 6, 37-39. [ Links ]
Ayala, M., L. Argel, S. Jaramillo y M. Marín. 2008. Diversidad genética de Peronospora sparsa (Peronosporaceae) en cultivos de rosa de Colombia. Acta Biol. Colomb. 13, 79-94. [ Links ]
Cohen, Y. y J. Rotem. 1971. Field and growth chamber approach to epidemiology of Pseudoperonospora cubensis on cucumbers. Phytopathol. 61, 736-737. [ Links ]
Cohen, Y. 1981. Capítulo 16: Downy mildew of cucurbits. pp. 345- 348. En: Spencer, D.M. (ed.). The downy mildews. Academic Press, London. [ Links ]
Cruickshank, I. 1961. Germination of Peronospora tabacina: effect of temperature. Aust. J. Biol. Sci. 14, 58-65. [ Links ]
Cruickshank, I. 1963. Environment and sporulation in phytopathogenic fungi. IV. The effect of light on the formation of conidia of Peronospora tabacina Adam. Aust. J. Biol. Sci. 16, 88-98. [ Links ]
De Vis, R. 1999. Manejo del mildeo velloso en rosa con control climático. Acopaflor 6, 14-19. [ Links ]
Giraldo, S., C. García y F. Restrepo. 2002. Influencia de la luz y la temperatura en la germinación de esporangios de Peronospora sparsa Berkeley, en rosa cultivar Charlotte. Agron. Colomb. 20, 31-37. [ Links ]
Gómez, S. 2004. Determinación de componentes de la biología de Peronospora sparsa Berkeley, y caracterización de la respuesta de tres variedades de rosa a la infección del patógeno bajo condiciones de laboratorio e invernadero. Tesis de maestría. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. [ Links ]
Gómez, S. y G. Arbeláez. 2004. Biología de Peronospora sparsa agente causal de mildeo velloso en rosa y su relación con el desarrollo de la enfermedad bajo condiciones de invernadero en la Sabana de Bogotá. En: XXV Congreso Ascolfi. 11-13 de agosto de 2004. Palmira, Colombia. [ Links ]
Gómez, S. y G. Arbeláez. 2005. Efecto de la temperatura en el periodo de latencia y producción de esporangios de Peronospora sparsa Berkeley en tres variedades de rosa. Agron. Colomb. 23, 239-245. [ Links ]
Hildelbrand, P.D. y J.C. Sutton. 1984. Interactive effects of the dark period, humid period, temperature, and light on sporulation of Peronospora destructor. Phytopathol. 74, 1444-1449. [ Links ]
Horst, R. 1983. A compendium of rose disease. APS Press, St. Paul, MN. [ Links ]
Pegg, G. y M. Mence. 1970. Colonization and sporulation of Peronospora viciae on cultivars of Pisum sativum. Ann. Appl. Biol. 66, 417-428. [ Links ]
Populer, C. 1981. Capítulo 5: Epidemiology of downy mildews. pp. 78-88. En: Spencer, D.M. (ed.). The downy mildews. Academic Press, London. [ Links ]
Raffray, J. y L. Sequeiro. 1971. Dark induction of sporulation in Bremia lactucae. Can. J. Bot. 49, 237-239. [ Links ]
Schmitt, C. y R. Freytag. 1974. Evaluation of greenhouse inoculation techniques to screen sorghum for resistance to downy mildew. Plant Dis. 58, 825-829. [ Links ]
Urrea, K. y G. Arbeláez. 2004. Evaluación de la eficacia de fungicidas para el control del mildeo velloso (Peronospora sparsa) en un cultivo comercial de rosa bajo invernadero. En: XXV Congreso Ascolfi. 11-13 de agosto de 2004. Palmira, Colombia. [ Links ]
Virnnot-Bourgin, G. 1981. Capítulo 1: History and importance of downy mildews. pp. 1-13. En: Spencer, D.M. (ed.). The downy mildews. Academic Press, London. [ Links ]
Weston, W. 1923. Production and dispersal of conidia in the Philippine. Sclerosporas of maize. J. Agr. Res. 23, 239-277. [ Links ]
Weston, W. 1924. Nocturnal production of conidia by Sclerospora graminicola. J. Agr. Res. 27, 771-784. [ Links ]
Wheeler, B. 1981. Capítulo 22: Downy mildew of ornamentals. pp. 476-477. En: Spencer, D.M. (ed.). The downy mildews. Academic Press, London. [ Links ]
Yarwood, C. 1937. The relation of light to the diurnal cycle of sporulation otcertain downy mildews. J. Agr. Res. 54, 365-373. [ Links ]

