










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.27 no.2 Bogotá May/Aug. 2009
FISIOLOGÍA Y TECNOLOGÍA DE POSCOSECHA
Javier Giovanni Álvarez-Herrera1,3, Jesús Antonio Galvis2 y Helber Enrique Balaguera-López1
1 Grupo de Investigaciones Agrícolas, Facultad de Ciencias Agropecuarias, Universidad Pedagógica y Tecnológica de Colombia (UPTC), Tunja (Colombia).
2 Facultad de Ingeniería de Alimentos, Fundación Universitaria Agraria de Colombia (Uniagraria), Bogotá (Colombia).
3 Autor de correspondencia. jgalvarezh@gmail.com
Fecha de recepción: 14 de abril de 2008. Aprobado para publicación: 2 de julio de 2009
RESUMEN
A pesar de su agradable aroma, y de poseer una pulpa jugosa, de atractivo sabor dulce acidulado, el fruto de champa es altamente perecedero. Por esta razón, la comercialización se ha limitado al procesamiento del fruto y/o a su venta en zonas de producción, ya que de otra parte, aún no se ha determinado el comportamiento del producto para poder programar su recolección, y plantear alternativas de comercialización. En este contexto, el objetivo del presente trabajo fue conocer la variación de las características fisicoquímicas de los frutos de champa cosechados en cinco estados de madurez, a partir de los cuales se determinaron la masa, el color, la firmeza, el pH, los sólidos solubles totales (SST), la acidez total titulable (ATT), los contenidos de sacarosa, glucosa y fructosa, y la relación de madurez. Para esto se tomaron 5 frutos por cada estado y por cada medición. Se encontró que la pulpa más la corteza constituyen el 60% de la masa fresca del fruto; el 40% restante corresponde a las semillas. La corteza del fruto varía de color verde a amarillo intenso, indicando la madurez de consumo. La firmeza disminuyó de 85 a 9,5 N para frutos maduros. Los SST se incrementan a medida que el fruto madura. La ATT disminuye durante el proceso de maduración. La sacarosa es el azúcar con mayor concentración en la champa, seguido por la fructosa y la glucosa.
Palabras clave: azúcares, sacarosa, frutas tropicales, precosecha.
ABSTRACT
In spite of its pleasant aroma and the tasty flavor of its juicy, sweet and mildly acid pulp, the champa fruit is highly perishable. For this reason, it is only locally commercialized, unless it has been processed; all the more because its behavior has not been studied yet, therefore making it difficult to program its harvesting and to seek for commercial alternatives. The objective of the present research was to assess the physico-chemical variation of the fruit along five stages of its maturation process. For such purpose, at each maturation stage, five fruits were selected to measure each of the following parameters: mass, color, firmness, pH, Total Soluble Solids (TSS), Total Titratable Acidity (TTA), maturity ratio and sucrose, glucose and fructose contents. The pulp was determined to be 60% of the fruit's fresh mass, while the seeds account for the resting 40%. Along the whole maturation process, the skin of the fruit goes from green to yellow, when it is finally ready for consumption; fruit firmness varied from 85 to 9.5 N for mature fruits; TSS increased, and TTA decreased. Sucrose is the most abundant sugar, followed by fructose and glucose.
Key words: sugars, sucrose, tropical fruits, preharvest.
Introducción
La champa (Campomanesia lineatifolia R. & P.) es un árbol frutal perteneciente a la familia Myrtaceae (Garzón y Barreto, 2002). Es una especie nativa de la Amazonia, de la cual en Colombia se han encontrado diferentes materiales en el Chocó, Amazonas, Caquetá, Casanare, Guaviare y en regiones interandinas cálidas de Boyacá y Cundinamarca. Se conoce en diferentes países y regiones con el nombre de palillo, guayabo anselmo, guayaba de leche, guayaba de mono, guabiroba o gabiroba (guarani) y michinche (López y Rodríguez, 1995; Villachica, 1996).
El fruto de champa es de sabor y aroma agradables, con abundante pulpa jugosa, dulce acidulada, lo que le da buena posibilidad de competir con otras especies ya conocidas. Los frutos se consumen en fresco, y con la pulpa se preparan dulces, mermeladas y jaleas (Villachica, 1996).
La semilla es de sabor amargo, y de esta se han extraído compuestos β-tricetona, denominados champanonas A, B y C, los cuales presentan actividad antimicrobiana (Bonilla et al., 2005). Osorio et al. (2006), al extraer compuestos volátiles de la pulpa, corteza, semilla y hoja de champa encontraron que las β-tricetonas contribuyeron en todos los estados de crecimiento del fruto a expresar los aromas y sabores característicos de la champa.
La cosecha es anual, la mayor producción se presenta en el mes de septiembre. El período de cosecha es de 80 a 90 días en el año (López y Rodríguez, 1995), por lo que la duración del producto en el mercado es baja, lo cual ha hecho que su popularización se vea reducida a las zonas de producción (Villachica, 1996).
La producción por árbol de champa puede oscilar desde los 5 kg para árboles jóvenes hasta los 500 kg para árboles adultos; la cosecha se hace cada tercer día y se pueden recolectar hasta 75 kg por árbol en cada jornada de recolección. No obstante, se presentan pérdidas en la poscosecha cercanas al 97% de la producción, debido a que los frutos tienen una vida poscosecha muy corta que se evidencia en un ablandamiento drástico del fruto y una rápida fermentación que limita su comercialización en fresco y su transformación a pulpa (López y Rodríguez, 1995). Otro agravante es que los frutos son recogidos del suelo a medida que se desprenden del árbol (Villachica, 1996); esta situación hace que se acelere su deterioro, pues no se ha determinado ningún índice de madurez para programar las cosechas, por lo cual, en la mayoría de los casos el fruto se pierde, o su comercialización se limita a centros regionales cercanos al lugar de producción.
Por tanto, el trabajo tuvo como objetivo determinar la variación de las principales características físicas y químicas de los frutos de champa en cinco estados de madurez, con el propósito de determinar un punto óptimo de cosecha que garantice la recolección del fruto directamente del árbol, y que de esta manera se eviten altas pérdidas en poscosecha.
Materiales y métodos
Los frutos utilizados se cosecharon en cinco estados de madurez, en las fincas "El Mango" y "La Playa" de la vereda Ayata del municipio de Miraflores (Boyacá), ubicado a 1.432 msnm, 5° 11' N y a 73° 08' W, con una precipitación promedio de 2.500 mm, temperatura promedio de 20°C y humedad relativa del 80%.
Las mediciones de las características físicas y químicas de los frutos de champa fueron realizadas en el Laboratorio del Instituto de Ciencia y Tecnología ICTA, de la Universidad Nacional de Colombia, sede Bogotá.
Se determinaron las características físicas de los frutos en cinco estados de madurez, los cuales se clasificaron con base en el color, como se muestra en la Tab. 1, tomando cinco frutos por estado y por medición. La masa fresca se midió con una balanza Acculab VIC 612 (Bradford, MA) de 0,01 g de precisión; la masa seca se obtuvo después de someter los frutos en una mufla a 90°C durante 36 h; el color, mediante la tabla de colores de Munsell, y la firmeza (N, Newtons), mediante un penetrómetro Bertuzzi Fruit Tester FT327 (Facchinni, Italy).

Las determinaciones químicas realizadas fueron sólidos solubles totales (SST) como porcentaje de grados Brix con un refractómetro Carl Zeiss (Jena, Alemania). El pH se determinó con un potenciómetro Orión, modelo 420 (Boston, MA); la acidez total titulable (ATT), mediante la titulación con NaOH 0,1 N hasta pH 8,2 y se expresó como ácido málico. La relación de madurez (RM) se estableció como el cociente entre los SST y la ATT. La determinación de sacarosa, glucosa y fructosa se precisó por cromatografía líquida de alta resolución (HPLC), con un detector de índice de refracción y una columna Sugar Pack.
Para las diferentes variables medidas se realizó un análisis descriptivo y un análisis de regresión no lineal con el fin de establecer los modelos de mayor ajuste al proceso de crecimiento del fruto mediante el software SAS® v. 8.1e (Cary, NC).
Resultados y discusión
Al momento de la madurez de consumo (estado 5), se determinó la masa fresca y seca de los frutos de champa (Fig. 1), así como la masa por separado de las semillas y de la pulpa más la corteza, dado que acorde con lo mencionado por Villachica (1996), es muy dispendioso separar la corteza de la pulpa.
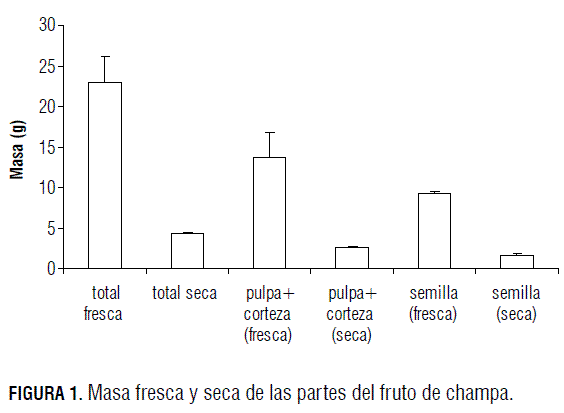
La pulpa más la corteza en fresco ocupan el 60% de la masa del fruto, el 40% restante está conformado por la masa de cuatro a seis semillas. Al secar los frutos, la relación en masa seca sigue constante, lo que indica que la pulpa y la semilla acumulan la misma cantidad de agua en porcentaje, situación que genera semillas recalcitrantes pues presentan gran contenido de humedad, de acuerdo con lo demostrado por Carvalho et al. (1997).
En la Fig. 2 se aprecia la evolución de la masa de los frutos de champa a través de los diferentes estados de madurez. La curva muestra un comportamiento sigmoideo en el cual el fruto presenta una ganancia de masa entre el estado 2 y 3 mayor que en los otros estados, proceso dominado en esta fase por el llenado de fruto y, posiblemente, por la disminución en la respiración (Kays, 2004). El modelo logístico sigmoideo es el de mayor ajuste comparado con el modelo de Gompertz pues presenta el menor cuadrado medio del error residual.

En la Tab. 1 se describe el color del fruto según la tabla de Munsell para cada estado de madurez indicado. El fruto de champa en sus estados iniciales presenta un color verde intenso que lentamente cambia a un verde más claro, para luego pasar a un verde amarillento, luego a un amarillo claro y finalmente a un amarillo intenso; este último color de la corteza es el indicativo de madurez de consumo del fruto.
Estos cambios en la coloración del fruto se deben a la degradación de la clorofila por la actividad de enzimas del tipo clorofilasa y clorofila oxidasa (Valpuesta et al., 1996), que hacen desenmascarar los colores amarillos (carotenoides) a medida que el fruto madura (Kays, 1997). Esta coloración amarilla en algunos casos se presenta con el fin de evitar la fotoinhibición (Matile et al., 1999). La degradación de la clorofila comienza con la disociación del residuo de fitol y del anillo de porfirina de la molécula de clorofila, reacción que es catalizada por la enzima clorofilasa (Kariola et al., 2005). La pulpa también cambió de color durante la maduración del fruto, y pasó de un color blanco en los estados iniciales a un color crema, lo que indica la posible existencia de xantofilas (Kays, 1997).
Los valores de firmeza se presentan en la Fig. 3. Estos muestran una disminución progresiva de la firmeza de los frutos de champa conforme la fruta madura, y oscilan entre 85,0 N para frutos en el primer estado de madurez y 9,5 N cuando la fruta se encuentra en madurez de consumo, lo que implica una reducción drástica de la firmeza.
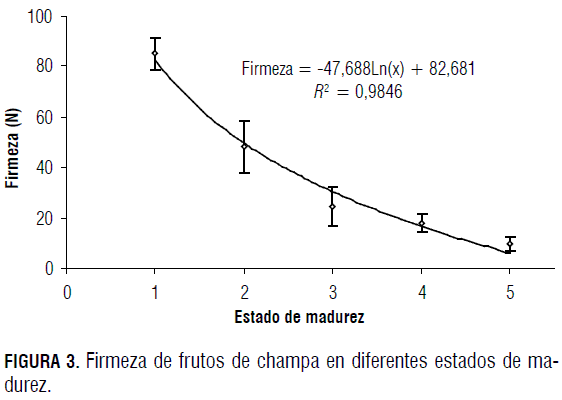
Al comparar la firmeza de la champa en estado de madurez (9,5 N) con la de frutos promisorios como la uchuva en su último estado de madurez (3,4 N) (Trinchero et al., 1999), resulta ser un poco mayor, pero al obsevar la firmeza en los estados iniciales 85 y 18 N para champa y uchuva respectivamente, se deduce que la pérdida de firmeza es mucho mayor en el fruto de champa, lo que indica que probablemente el proceso de maduración es acelerado y que la absorción de calcio y la traslocación del mismo hacia los frutos, probablemente, en este estado es insuficiente (Marschner, 2002).
Durante la maduración de los frutos ocurre pérdida de firmeza de forma logarítmica, debido principalmente a cambios en la estructura de la pared celular; estos implican la actividad de enzimas capaces de degradar los componentes de la misma. Las principales enzimas involucradas en el proceso de maduración son la poligalacturonasa, celulasa y pectinmetilesterasa (Zarra y Revilla, 1996), y en algunos casos glicosidasas (Gross et al., 1995; Trinchero et al., 1999; Minic y Jouanin, 2006).
Esta degradación de la pared celular que ocurre durante la maduración se da principalmente en los poligalacturónidos debido a la acción de las poligalacturonasas. Del mismo modo se ha observado que la pectinmetilesterasa cataliza la deesterificación del grupo carboxilo del ácido poligalacturónico e incrementa la susceptibilidad de las pectinas a la poligalacturonasa (Zarra y Revilla, 1996; Kays, 1997). También puede ocurrir la presencia de celulasas, como la endo β(1-4) D-glucanasa, la cual actúa sobre polisacáridos hemicelulósicos del tipo xiloglucano y β(1-4) (1-3)glucano (Zarra y Revilla, 1996).
La importancia de la firmeza radica en que es uno de los parámetros más importantes para determinar tanto la calidad como el posible manejo y empaque del producto (Kays, 1997); debido a que los frutos de champa pierden esta propiedad rápidamente, su manejo se convierte en un aspecto crítico en la poscosecha de este fruto.
La variación de la cantidad de SST se presenta en la Fig. 4. Se aprecia un aumento constante de la cantidad de azúcares a medida que el fruto madura, al pasar de 9,5 a 11,2%, aunque en el estado 4 se presenta una cantidad de 11,9%, lo que indica, probablemente, que este es el punto óptimo de cosecha, y que después de este estado el fruto comienza a incrementar su respiración y a utilizar sustratos como la glucosa en el proceso metabólico.
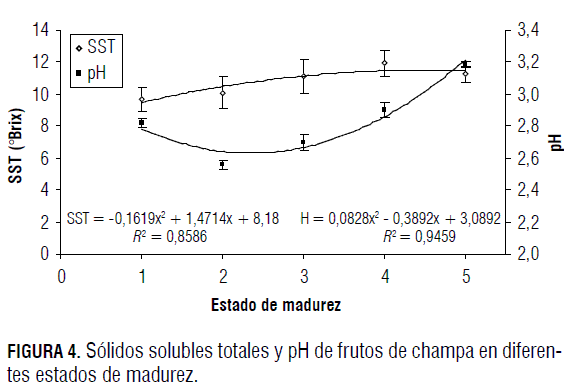
La cantidad de SST se incrementa hasta el estado de madurez 4, debido a la hidrólisis del almidón almacenado en vacuolas y espacios intercelulares durante el crecimiento del fruto. Este proceso ocurre debido al marcado incremento en la actividad de las enzimas a-amilasa, β-amilasa y almidón fosforilasa (Kays, 1997). Al parecer sólo la a-amilasa puede degradar gránulos de almidón intactos, por lo que la actividad de la β-amilasa y almidón fosforilasa se limitan a actuar sobre los primeros productos liberados del almidón por la a-amilasa (Salisbury y Ross, 1994).
El almidón en las células es almacenado en pequeños gránulos y a temperatura ambiente (20°C); es de difícil degradación tanto por el agua como por las enzimas hidrolíticas, debido a la formación de puentes de hidrógeno dentro de las mismas moléculas de almidón. Una vez se incrementa la temperatura, los puentes de hidrógeno se rompen y el agua es absorbida por las moléculas de almidón, lo que produce la ruptura y gelatiniza los gránulos (Zobel, 1984).
En la Fig. 4 se observa la variación del pH en los frutos de champa; a medida que maduran, este disminuye del estado 1 al estado 2, y posteriormente se incrementa desde el estado 2 hasta el estado 5.
La disminución del pH en los primeros estados puede estar relacionada con el aumento que se da en la concentración de ácidos orgánicos (Fig. 5). El incremento en el pH a partir del estado 2 se da porque durante el llenado de frutos gran parte de la actividad de acumulación se da por simporte, en donde los iones H+ desempeñan un papel importante; estos hacen parte de la formación de sustratos como la sacarosa y la glucosa, y hacen que su concentración a nivel vacuolar disminuya durante las últimas fases de la maduración, por lo que el pH se ve ligeramente aumentado (Marschner, 2002).
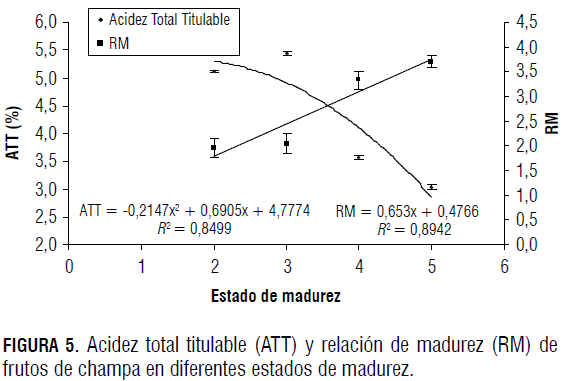
La ATT disminuye a medida que el fruto de champa madura (Fig. 5), como ocurre en la mayoría de los frutos (Hobson, 1996; Kays, 1997). Esta disminución se da debido a la actividad de las deshidrogenasas y a que los ácidos orgánicos son utilizados como sustratos de la respiración para la síntesis de nuevos componentes durante la maduración. Este descenso coincide con el inicio de la maduración y la acumulación de azúcares (Kays, 1997).
La mayoría de los frutos contienen grandes concentraciones de ácidos derivados del ciclo de los ácidos tricarboxílicos y otros ciclos metabólicos, los cuales se acumulan en la vacuola, en algunos frutos en forma de ácido málico, el cual se oxida a CO2, o se convierte en ácido cítrico o glutámico, lo que explica el descenso del ácido málico durante la maduración (Hobson, 1996).
La relación de madurez expresada como SST/ATT mide la calidad organoléptica de los frutos. Esta relación presenta una tendencia a aumentar conforme el fruto madura (Fig. 5), con un mayor incremento entre los estados 3 y 4. La RM en los frutos de champa está relacionada principalmente con un aumento de los SST y una disminución de la ATT. En los frutos climatéricos el aumento de la RM posiblemente ocurre cuando alcanzan la tasa respiratoria máxima y desdoblan rápidamente sus reservas (ácidos orgánicos) como respuesta al incremento de su metabolismo y, en consecuencia, la RM se incrementa (Kays, 2004). Sin embargo, hay que tener en cuenta que cuando el fruto tiene un contenido alto de azúcares, el nivel de los ácidos debe ser suficientemente elevado para satisfacer el gusto del consumidor (Osterloh et al., 1996).
Los valores de RM oscilan entre 2 y 4, aproximadamente; estos se pueden considerar bajos al comparar las RM de frutos feijoa (Acca sellowiana Berg) reportadas por Rodríguez et al. (2006), las cuales se encuentran entre 4,0 y 11,6. Esta situación se puede presentar probablemente porque la champa presenta una mayor acidez titulable (3-5%) en todos los estados de madurez, mientras que la ATT de la feijoa varía desde 2,5 hasta 1,0.
En la Fig. 6 se observa la variación del contenido de sacarosa, glucosa y fructosa durante el proceso de maduración del fruto de champa. En esta se aprecia una acumulación de sacarosa durante todo el proceso, mientras que la glucosa y la fructosa sólo aumentan hasta la fase de maduración 4, por lo que se supone que en esta fase comienzan a ser sustrato en el proceso respiratorio del fruto (Hobson, 1996).
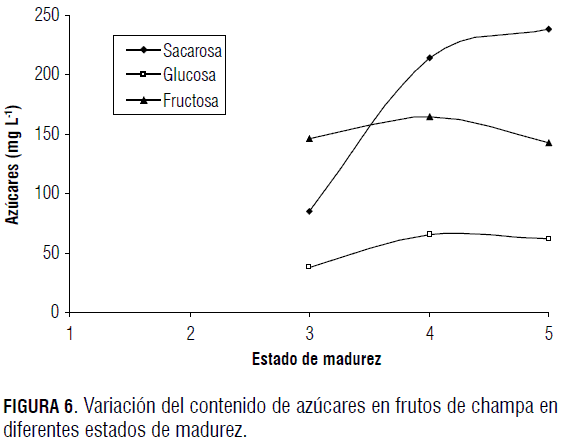
La sacarosa presentó una mayor concentración en comparación con los demás azúcares, al igual que en la mayoría de los frutos (Wills et al., 1998). El incremento de sacarosa corresponde a la traslocación de fotoasimilados que se da desde las hojas y hacia el fruto durante el crecimiento del mismo y que está directamente relacionado con el aumento en la actividad de la sacarosa fosfato sintetasa y la disminución de la invertasa (Lowell et al., 1989; Kubo et al., 2001).
Según Forney y Breen (1986), en fresas el proceso de entrada de sacarosa al fruto se da vía apoplasto y la mayoría del sustrato se hidroliza antes de su asimilación, proceso que se da debido a que al final del crecimiento del fruto todavía existe traslocación de sacarosa desde las hojas más cercanas (Fischer y Lüdders, 1997). En guayaba y feijoa el azúcar predominante es la fructosa (Rodríguez et al., 2006), mientras que para champa el azúcar con mayor concentración es la sacarosa, al igual que en Eugenia stipitata (Galvis y Hernández, 1993).
La glucosa y la fructosa se incrementan en las primeras fases de maduración del fruto, debido a que la mayoría de estos acumulan almidón en sus estados iniciales (Dinar y Stevens, 1981), y se originan azúcares más sencillos durante la maduración por efecto de la marcada actividad de las enzimas a-amilasa, β-amilasa y almidón fosforilasa (Kays, 1997).
Luego de que el fruto se encuentra en su última fase de madurez (estado 4 en adelante), la concentración de glucosa y fructosa disminuye (Fig. 6) debido a dos posibles causas: la primera, a que pasan a ser sustrato respiratorio, y la segunda, a que el proceso de síntesis de estos azúcares disminuye, pero la cantidad de agua que entra al fruto no, por lo que la concentración de estos monosacáridos decrece.
La disminución de la fructosa puede ir directamente relacionada con el aumento de la sacarosa, ya que la fructosa fosforilada y retirada como hexosa libre facilita la formación del gradiente de sacarosa entre el floema y las células en los vertederos (Renz et al., 1993), en este caso el fruto; también puede ocurrir un proceso de reconversión en el que las hexosas libres mediante la acción de la sacarosa fosfato sintasa sean sintetizadas a sacarosa, la cual es transportada por medio de un mecanismo antiporte H+/sacarosa a través del tonoplasto (Fieuw y Willenbrink, 1990).
Cabe resaltar que dentro de los cromatogramas de azúcares analizados, además de los picos registrados para sacarosa, glucosa y fructosa, se presenta otro pico de refracción de mayor intensidad, lo que indicaría la posible presencia de otro azúcar específico para la champa en mayor concentración y por identificar.
Conclusiones
Con base en los resultados obtenidos se puede concluir que los frutos de champa pueden ser cosechados en el estado de madurez 4, que corresponde a 60% verde - 40% amarillo claro. En este estado los frutos presentan una firmeza de 15,55 N, un pH de 2,9, un contenido de SST de 11,93oBrix, 3,57% de ATT y una RM de 3,33. El azúcar con mayor concentración en los frutos de champa es la sacarosa.
Literatura citada
Bonilla, A., C. Duque, C. Garzón, Y. Takaishi, K. Yamaguchi, N. Hara e Y. Fujimoto. 2005. Champanones, yellow pigments from the seeds of champa (Campomanesia lineatifolia). Phytochem. 66, 1736-1740. [ Links ]
Carvalho, J.E.U., N.V.M. Leão y C.H. Muller. 1997. Sensibilidade de sementes de gabiroba (Campomanesia lineatifolia Ruiz et Pav. - Myrtaceae) ao dessecamento e à baixa temperatura. En: X Congresso Brasileiro de Sementes, Associaão Brasileira de Tecnologia de Sementes. Foz de Iguazú, Brasil. Abrates 7(1/2), 252. [ Links ]
Dinar, M. y M. Stevens. 1981. The relation between starch accumulation and soluble solids content of tomato fruits. J. Amer. Soc. Hort. Sci. 106, 415-418. [ Links ]
Fieuw, S. y J. Willenbrink. 1990. Sugar transport and surgar-metabolizing enzymes in sugar beet storage roots (Beta vulgaris ssp. Altissima). J. Plant Physiol. 137(2), 216-223. [ Links ]
Fischer, G. y P. Lüdders. 1997. Developmental changes of carbohydrates in cape gooseberry (Physalis peruviana L.) fruits in relation to the calyx and the leaves. Agron. Colomb. 14(2), 95-107. [ Links ]
Forney, C. y P. Breen. 1986. Sugar content and uptake in the strawberry fruit. J. Amer. Soc. Hort. Sci. 111, 241-247. [ Links ]
Galvis, J. y M. Hernández. 1993. Análisis del crecimiento del fruto y determinación del momento de cosecha del arazá (Eugenia stipitata). Colombia Amazónica 6(2), 107-121. [ Links ]
Garzón, C. y F. Barreto. 2002. Campomanesia lineatifolia Ruíz & Pav. en Colombia. p. 315. En: Libro de Resúmenes del Octavo Congreso Latinoamericano y Segundo Congreso Colombiano de Botánica. Etnobotánica y botánica económica. Asociación Latinoamericana de Botánica. 13-18 de octubre. Cartagena, Colombia. [ Links ]
Gross, K., D. Scarrett y H. Chen. 1995. Rhamnogalacturonase, a-galactosidase and B-galactosidase: potential role in fruit softening. Acta Hort. 398, 121-130. [ Links ]
Hobson, G.E. 1996. Maduración del fruto. pp. 463-478. En: Azcón- Bieto, J. y M. Talón (eds.). Fisiología y bioquímica vegetal. Interamericana McGraw-Hill, Bogotá. [ Links ]
Kariola, T., G. Brader, J. Li y T. Palva. 2005. Chlorophyllase 1, a damage control enzyme, affects the balance between defense pathways in Plants. Plant Cell 17, 282-294. [ Links ]
Kays, S. 1997. Postharvest physiology of perishable plant products. Exon Press. Athens, GA. [ Links ]
Kays, S. y R.E. Paull. 2004. Postharvest biology. Exon Press, Athens, GA. [ Links ]
Kubo, T., I. Hohjo y S. Hiratsuka. 2001. Sucrose accumulation and its related enzyme activities in the juce sacs of satua mandarin fruit from trees with different crop loads. Scientia Hort. 91, 215-225. [ Links ]
López, M. y J. Rodríguez. 1995. Diagnóstico del mercadeo de la champa en el municipio de Miraflores, Boyacá. Trabajo de grado. Instituto de Educación Abierta y a Distancia, UPTC, Tunja. [ Links ]
Lowell, C., P. Tomlinson y K. Koch. 1989. Sucrose-metabolizing enzymes in transport tissues and adjacent sink structures in developing citrus fruit. Plant Physiol. 90, 1394-1402. [ Links ]
Marschner, H. 2002. Mineral nutrition of higher plants. 2a ed. Academic Press, London. [ Links ]
Matile, P., S. Hörtensteiner y H. Thomas. 1999. Chlorophyll degradation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50, 67-95. [ Links ]
Minic, Z. y L. Jouanin. 2006. Plant glycoside hydrolases involved in cell wall polysaccharide degradation. Plant Physiol. Biochem. 44, 435-449. [ Links ]
Osorio, C., M. Alarcón, C. Moreno, A. Bonilla, J. Barrios, C. Garzón y C. Duque. 2006. Characterization of odor-active volatiles in Champa (Campomanesia lineatifolia R. & P.). J. Agric. Food Chem. 54(2), 509-516. [ Links ]
Osterloh, A., G. Ebert, W.H. Held, H. Schulz y E. Urban. 1996. Lagerung von Obst und Südfrüchten. Verlag Ulmer, Stuttgart, Alemania. [ Links ]
Renz, A., L. Merlo y M. Stitt. 1993. Partial purification from potato tubers of three fructokinases and three hexokinases which show differing organ and developmental specificity. Planta 190, 156-165. [ Links ]
Rodríguez, M., H. Arjona y J. Galvis. 2006. Maduración del fruto de feijoa (Acca sellowiana Berg) en los clones 41 (Quimba) y 8-4 a temperatura ambiente a condiciones de Bogotá. Agron. Colomb. 24(1), 68-76. [ Links ]
Salisbury, F. y C. Ross. 1994. Fisiología vegetal. 4a ed. Grupo Editorial Iberoamérica, México. [ Links ]
Trinchero, G., G. Sozzi, A. Cerri, F. Vilella y A. Franschina. 1999. Ripening-related changes in ethylene production respiration rate and cell-wall enzyme activity in golden berry (Physalis peruviana L.) a solanaceous species. Postharv. Biol. Technol. 16, 139-145. [ Links ]
Valpuesta, V., M.A. Quesada y M.S. Reid. 1996. Senescencia y abscisión. pp. 479-492. En: Azcón-Bieto, J. y M. Talón (eds.). Fisiología y bioquímica vegetal. Interamericana McGraw-Hill, Bogotá. [ Links ]
Villachica, H. 1996. Frutales y hortalizas promisorios del Amazonas. pp. 181-185. Tratado de Cooperación Amazónica. Secretaría Pro Tempore, Lima. [ Links ]
Wills, R., B. McGlasson, D. Graham y D. Joyce. 1998. Postharvest - an introduction to the physiology and handling of fruit, vegetables and ornamentals. CAB International, Wallingford, UK. [ Links ]
Zarra, I. y G. Revilla. 1996. Pared celular. Estructura y función. pp. 1-24. En: Azcón-Bieto, J. y M. Talón. Fisiología y bioquímica vegetal. Interamericana McGraw-Hill, Bogotá. [ Links ]
Zobel, H. 1984. Gelatinization of starch and mechanical properties of starch pastes. pp. 285-311. En: Whistler, R., J. BeMiller y E. Paschall (eds.). Starch: chemistry and technology. Academic Press, London. [ Links ]

