










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.27 no.3 Bogotá Sep./Dec. 2009
Luis Ernesto Rodríguez1
1 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. lerodriguezmo@unal.edu.co
Fecha de recepción: 4 de mayo de 2009. Aceptado para publicación: 6 de noviembre de 2009
RESUMEN
La presente revisión busca reunir diferentes propuestas utilizadas para clasificar las papas cultivadas y sus parientes silvestres (Solanum L. sect. Petota Dumort.), ya que históricamente se han presentado diferencias entre los taxónomos que han estudiado su clasificación. A la fecha no hay consenso, debido a que el límite entre especies no está definido, y las interrelaciones entre estas son frecuentes. Es importante generar una propuesta definitiva, coherente y práctica, que genere un consenso en la taxonomía de la papa. Esto permitiría conocer el número real de especies, el límite entre ellas y sus interrelaciones y continuos cambios. A su vez, esta información deberá facilitar un mejor aprovechamiento del germoplasma por parte de los programas de mejoramiento genético.
Palabras clave: Solanum tuberosum, especies silvestres.
ABSTRACT
The present review seeks to bring together different proposals about the taxonomy of cultivated potatoes and their tuberbearing wild relatives (Solanum L. sect. Petota Dumort.). Throughout history, taxonomists have introduced discordant criteria about the classification of this group. Up to date, no agreement has been achieved, because the limits among species are not clear, and they frequently interact. It is therefore important to generate a consensual, coherent, practical and definite proposal for potato taxonomy that allows knowing the real number of species and their limits, interrelations and continuous changes. In turn, this information is necessary to facilitate better germplasm utilization in breeding programs.
Key words: Solanum tuberosum, tuber-bearing wild species.
Introducción
La papa y sus parientes silvestres Solanum L. sect. Petota Dumort. se distribuyen a lo largo del continente americano, desde el suroeste de Estados Unidos hasta Chile, Argentina y Uruguay (Hijmans y Spooner, 2001). La papa se considera uno de vegetales más productivos que se cultiva en el mundo, y provee la mayor fuente de nutrición e ingreso para muchas sociedades (Spooner y Hetterscheid, 2005). Ocupa un lugar importante en la agricultura, economía y seguridad alimentaria, sitúandose en el cuarto lugar de los cultivos o alimentos que sustentan la nutrición a nivel mundial, después del maíz, el trigo y el arroz, con una producción para el año 2007 de 325,3 millones de toneladas y un valor estimado de la cosecha de 40 mil millones de US dólares. Se espera que para 2020 la demanda de la papa sea el doble de la consumida en 1993 (FAO, 2009).
Actualmente, la papa cultivada es conocida colectivamente bajo el nombre de S. tuberosum L. (Spooner y Salas, 2006; Andre et al., 2007), posee un rico pool de genes, constituido por 190 especies silvestres que forman tubérculos (Spooner y Salas, 2006). La papa es el único grupo que posee poliploides, aproximadamente 70% de las especies son diploides, la mayoría de los restantes son tetraploides (2n = 6x = 72), con un número reducido de triploides y pentaploides (Spooner et al., 2005a; Hijmans et al., 2007; Spooner et al., 2008). Representan una reserva de germoplasma amplia y única, parcialmente explorada y poco usada en el mejoramiento genético, a pesar de que muchas de estas especies se pueden utilizar directamente en cruzamientos compatibles con papas cultivadas o a través del uso de gametos no reducidos 2n (Estrada, 2000; Spooner y Salas, 2006).
Existe controversia sobre la conveniencia de clasificar la papa como varias especies, bajo la clasificación propuesta por Linneaus (1753) y adoptada por el Código Internacional de Nomenclatura Botánica (ICBN-International Code of Botanical Nomenclature), o como única especie con diferentes grupos cultivados ubicados dentro de S. tuberosum, utilizando la nomenclatura propuesta por el Código Internacional de Nomenclatura de Plantas Cultivadas (Icncp-International Code of Nomenclature of Cultivated Plants) (Spooner y Hetterscheid, 2005).
La papa continúa siendo objeto de intensa colección e investigación taxonómica, con el fin de determinar un número real de especies (Spooner y Salas, 2006), debido principalmente a que gran parte de las especies se relacionan entre sí (Hawkes, 1990; Ochoa, 1990; van den Berg et al., 1998; Sukhotu et al., 2006; Spooner y Salas, 2006; Spooner et al., 2007). Los límites entre diferentes series son menores y aparentemente artificiales, razón por la cual muchas especies son morfológicamente similares y usualmente se distinguen por diferencias menores como la pubescencia o la forma de la hoja (Spooner y van der Berg, 1992). La hibridación ha contribuido en la formación de muchas especies tanto a nivel diploide como poliploide. La introgresión e hibridación interespecífica han sido sugeridas como algunas de las causas que dificultan la taxonomía en la sección Petota (Spooner et al., 2007), debido a que la hibridación interespecífica natural es común para muchas especies de la sección Petota (Ugent, 1970; Hawkes, 1963, 1990; Clausen y Spooner, 1998; Miller y Spooner, 1999).
Un ejemplo práctico es el origen de S. andigena. Se ha propuesto que proviene de S. stenotomum por poliploidización sexual durante muchas veces y en múltiples lugares, con la subsiguiente hibridación interespecífica e intervarietal a través de cruces (4x x 4x) o poliploidización a través de cruces (2x x 4x ó 4x x 2x) (Sukhotu y Hosaka, 2006). La presencia de tetraploidización sexual en papa ha sido sugerida por la capacidad de producir granos de polen 2n en las papas tetraploides y especies relacionadas (Iwanaga y Peloquin, 1982; Watanabe y Peloquin, 1989; Werner y Peloquin, 1991).
La clasificación taxonómica de la papa es una labor importante, que trata de organizar la variabilidad de una forma lógica; sin embargo, el nombre de una especie no es más que una hipótesis que representa a un grupo "relacionado entre sí por algo". Si se refinan los límites entre las especies silvestres de papa y se agrupan por características similares, esto constituiría una herramienta de predicción muy importante para cualquier programa de fitomejoramiento.
Es de gran relevancia generar una propuesta final y práctica de amplia aceptación y consenso entre taxónomos, mejoradores y productores, que permita una acertada clasificación taxonómica de las especies silvestres y cultivadas de papa, para facilitar un mejor aprovechamiento del germoplasma en los programas de mejoramiento genético.
El objetivo de esta revisión fue compilar las diferentes propuestas existentes para la clasificación de la papa cultivada y sus parientes silvestres.
Taxonomía de las especies de papa
La taxonomía permite describir, nombrar y clasificar los organismos (Lincoln et al., 1998). La sistemática es una disciplina más amplia que determina relaciones filogenéticas a través de métodos experimentales modernos, basados en la anatomía comparativa, citogenética, ecología, morfología y usando datos moleculares (Stuessy, 1990).
La clasificación de las plantas cultivadas y silvestres tienen objetivos diferentes: mientras que las plantas silvestres se clasifican en un sistema que intenta clarificar relaciones evolutivas, las plantas cultivadas se clasifican (o se deberían clasificar) según criterios definidos por los usuarios de acuerdo con un propósito especial, con estabilidad en los nombres primarios, teniendo como meta un sistema de clasificación totalmente diverso (Brandenburg, 1986).
Existe controversia sobre la taxonomía de las papas cultivadas, debido a que diferentes autores han reconocido desde una hasta 20 especies (Huamán y Spooner, 2002), pero todas forman un mismo pool genético (gene pool) (Spooner et al., 2005b).
Resultados morfológicos y moleculares recientes clasifican la papa en 190 especies repartidas en cuatro clados, que presentan algunas relaciones inter específicas y gran variabilidad con la clasificación propuesta inicialmente en 1990 por Hawkes (Spooner et al., 2008). Spooner et al. (2007) ponen en tela de juicio muchas de estas series y listan únicamente 189 especies, a pesar de la descripción de diez nuevas especies desde Hawkes (1990), y predicen que en un futuro se presentará una nueva reducción en el número de especies, en la medida que continúen sus trabajos de investigación con marcadores moleculares.
La taxonomía de las especies silvestres y cultivadas de papa continúa siendo notablemente compleja (Hijmans et al., 2002; Hijmans et al., 2003; Spooner et al., 2003c; Spooner et al., 2004; Spooner y Salas, 2006; Spooner et al., 2007; Spooner et al., 2008), genera controversias y actualmente sigue siendo refinada (Hijmans et al., 2002; Sponner et al., 2007). Muchas especies de papa son iguales, aunque tengan apariencia diferente, ya que mantienen la capacidad de hibridarse cuando están en contacto. El límite entre especies es confuso y aún falta definición para entenderlo claramente (Spooner y Lara-Cabrera, 2001; Spooner y Salas 2006).
A diferencia de los demás cultivos de importancia agronómica, como el maíz, el trigo y el arroz, para la papa no se ha considerado el mismo concepto taxonómico de gene pool; solamente se ha realizado una separación en diferentes series, una de las cuales corresponde a las papas cultivadas, esto como consecuencia a que no hay barreras marcadas para el flujo de genes en las diferentes categorías (Spooner et al., 2003b; Spooner et al., 2008).
Muchos taxónomos han aplicado diversos conceptos para reconocer y agrupar las especies de papa (Spooner y van den Berg, 1992). Actualmente, la sistemática molecular que utiliza análisis de ADN genómico proporciona una herramienta de alta confiabilidad, que ha conducido a modificaciones recientes en la clasificación taxonómica, y se espera ayude a despejar muchas dudas acerca de la relación entre especies (Spooner et al., 2003c).
Las especies silvestres y cultivadas de papa (Solanum sección Petota) son un desafío para los taxónomos, por la ausencia de una definición clara de las diferencias morfológicas entre algunas especies, la plasticidad fenotípica cuando se siembran en diferentes ambientes, la compatibilidad sexual entre muchas de las especies, el amplio rango en los niveles de ploidía -desde diploides hasta hexaploides-, la especiación híbrida y la hibridación introgresiva (Spooner et al., 2003c; Spooner et al., 2007).
De acuerdo con Hijmans et al. (2002), la clasificación moderna tiene sus raíces en el sistema de Linnaeus (1753), quien describió la papa cultivada común como la especie S. tuberosum L., pero no hizo lo mismo con ninguna especie silvestre. Adicionalmente, propuso la clasificación jerárquica de las especies según sus características físicas compartidas y normalizó su denominación. Linnaeus (1753) propuso la fórmula binominal, consistente en el uso de dos palabras para asignar un nombre único para cada especie: la primera es el nombre del género y la segunda es el epíteto específico (Knapp et al., 2004).
Estudios morfológicos (fenéticos), acompañados de datos moleculares utilizando isoenzimas, FLP, AFLP o SSR de ADN, se han aplicado para definir el límite entre especies silvestres y cultivadas de papa (Huamán y Spooner, 2002; Spooner et al., 2003a).
Aspectos moleculares
Actualmente se ha recurrido a los marcadores moleculares para resolver muchos interrogantes, entre estos la determinación de especies, las relaciones entre especies, la biogeografía, la confirmación de hibridaciones e introgresiones naturales y artificiales, la evolución de la nodulación, la concordancia de distintos conjuntos de datos, la determinación de marcadores diagnósticos de registros de especies particulares, la determinación de herencia auto poliploide y alo poliploide, la diversidad genética, la heterosis, el mantenimiento de bancos genéticos, la evolución de la domesticación y la evolución del genoma (Spooner y Lara-Cabrera, 2001).
La elección de métodos analíticos adecuados para investigar los límites entre especies está sujeta a un interrumpido debate aún por resolverse. Las especies se definen de distintas maneras mediante criterios morfológicos, biológicos y cladísticos (Spooner et al., 2003c).
Las especies morfológicas han sido definidas con base en criterios fenéticos intuitivos o computarizados (Sokal y Crovello, 1970). Las especies biológicas dependen de su capacidad real o potencial de cruzamiento (Mayr, 1969). Recientemente se han aplicado los conceptos cladísticos para la determinación de las especies. Los cladistas investigan el parentesco entre progenitor y derivado, con base en que estos comparten un carácter derivado determinado a partir del grado de proximidad de los taxones emparentados o grupos derivados (Hennig, 1966).
La filogenia estudia las interrelaciones entre organismos y proporciona información de cómo se relacionan especies, géneros y familias; en la actualidad se utiliza basándose en los principios de la cladística, de tal manera que un grupo de especies ubicado dentro de un clado monofilético implica que todos los descendientes provienen de un antecesor común. Sin embargo, cada filogenia no es más que una hipótesis acerca de las relaciones entre los organismos bajo estudio (Knapp et al., 2004).
La construcción de árboles filogenéticos para muchos grupos de organismos ha revolucionado el campo de la sistemática y ha conducido a nuevas interpretaciones acerca de cómo los grupos, tanto de especies silvestres como cultivadas, están relacionados, realizando un análisis de diferentes caracteres dentro de un marco evolutivo y permitiendo una unión entre la taxonomía y la genómica a través de la descripción de diferentes especímenes (Knapp et al., 2004).
La investigación en taxonomía está plenamente justificada como una herramienta predictiva en el mejoramiento de las plantas; debe proporcionar una guía útil para hacer predicciones que permitan utilizar o no germoplasma de acuerdo con su relación con otras especies, y aprovechar así fuentes de resistencia a diferentes factores bióticos, abióticos o rasgos agronómicos de importancia, que generalmente se encuentran organizados en la literatura utilizando la taxonomía y permiten un mejor aprovechamiento de los recursos genéticos (Spooner y Salas, 2006).
Recientes estudios morfológicos, fenéticos y moleculares han confirmado la ausencia de una definición clara de las diferencias entre especies de la sección Petota, y han mostrado la utilización de un número de caracteres con rangos sobrepuestos para la delimitación de especies, lo que traería como consecuencia la reducción en el número de especies de la sección Petota. Adicionalmente, estudios moleculares recientes han sugerido la necesidad de reconsiderar el concepto tradicional para mantener las series actuales (Spooner et al., 2003a).
Diferentes autores han propuesto la necesidad de modernizar la clasificación y nomenclatura de las plantas cultivadas, especialmente la revisión sobre la clasificación de la papa, tanto silvestre como cultivada (Spooner et al., 2003c; Spooner y Hetterscheid, 2005), buscando una clasificación moderna, ágil y estable definida bajo los parámetros de clasificación de las plantas cultivadas establecida por el International Code of Nomenclature for Cultivated Plants, Icncp (Brickell et al., 2004).
Las plantas cultivadas evolucionan mediante los efectos drásticamente diferentes de la selección artificial -que incluyen hibridación interespecífica, intergenérica, transgénesis y selección humana- hacia rasgos de interés agronómico, como el incremento en la variabilidad y el gigantismo de los órganos de interés, cambios en el hábito de crecimiento, hábitat y formas de cultivo (Spooner y Hetterscheid, 2005), que comparados con la selección natural, pueden tener un profundo efecto en su morfología y, por consiguiente, en su clasificación (Spooner y Lara-Cabrera, 2001).
La divergencia entre los objetivos de clasificación para plantas silvestres y cultivadas se ha mantenido por el uso del término taxón, utilizado en la taxonomía de plantas silvestres; se plantea un término alternativo: cultón, o grupo de usuarios definidos, para remplazar taxón, término usado hoy principalmente para organismos filogenéticamente relacionados (Spooner et al., 2003a; Hetterscheid y Branderberg, 1995).
El Icncp utiliza un sistema no jerárquico, que es una clasificación abierta incompatible con el sistema clásico para clasificar las plantas cultivadas; en este código se utilizan solamente dos rangos -cultivar y grupo cultivado-, que se pueden colocar donde quiera bajo un género, una especie, una subespecie, una variedad o una forma; además, el mismo cultivar puede pertenecer al mismo tiempo a diversos grupos cultivados (Ochsmann, 2004b).
La clasificación según el International Code of Botanical Nomenclature (ICBN) implica normalmente un fondo filogenético, mientras que el código del Icncp tiene como objetivo proporcionar una clasificación formal del uso práctico. El concepto de cultón como sistema no jerárquico es incompatible con el sistema jerárquico del ICBN, que da lugar a problemas con las convenciones conocidas. La aceptación del Icncp es baja:
i) para ciertos aspectos taxonómicos hay una carencia de categorías aceptadas;
ii) las reglas para nombrar cultivares son inmóviles, demasiado complicadas o restrictivas para el uso práctico, y
iii) un sector comercial importante con marcas registradas no es cubierto por el Icncp (Ochsmann, 2004b).
En el futuro, se espera el uso de códigos apropiados e incluir las reglas para nombrar clados, debido a la cantidad e importancia de los datos moleculares. Un gran avance en este proceso será el desarrollo y el establecimiento de bases de datos mundiales que proporcionan las herramientas para ligar y mantener la información en relación con los nombres de la planta (Ochsmann, 2004a).
Actualmente, la taxonomía de las especies silvestres y cultivadas de papa sigue siendo muy complicada, ya que usualmente muchas especies de papa presentan una apariencia muy diferente, pero mantienen la capacidad para hibridarse de forma natural cuando se encuentran en contacto, lo que genera alta variabilidad y hace difícil determinar los límites entre especies (Spooner y Salas, 2006).
A la fecha no hay consenso acerca de la clasificación de la papa (Spooner y Salas, 2006). A lo largo de la historia se han presentado diferentes puntos de vista entre los taxónomos que se han encargado de la clasificación de la papa (Tab. 1). Además, muchos investigadores han aplicado diferentes conceptos taxonómicos para reconocer grupos y especies. Los niveles de ploidía de las papas cultivadas varían en un rango que va desde las papas diploides (2n = 2x = 24), triploides (2n = 3x = 36), tetraploides (2n = 4x = 48) hasta las pentaploides (2n = 5x = 60) (Hijmans y Spooner, 2001; Spooner et al., 2004; Spooner y Salas, 2006).
Las papas cultivadas han sido clasificadas como especies, según el ICBN, inicialmente por Bukasov (1971) y Lechnovich (1971) y luego por Hawkes (1990) y Ochoa (1990 y 1999), mientras que Dodds (1962), Huamán y Spooner (2002) y Spooner et al. (2007) han utilizado simultáneamente la clasificación como especies, y dentro de estas, grupos cultivados utilizando la nomenclatura propuesta por el Icncp.
Hawkes (1990) proporcionó el último esfuerzo por clasificar las papas silvestres formalmente y reconoció 21 series, que incluían especies tuberosas y no tuberosas. Estudios hechos por Spooner y Sytsma (1992), Spooner et al. (1993) y Spooner y Castillo (1997) mostraron que las especies no tuberosas no pertenecen a la sección Petota y que toda la serie de Hawkes no es monofilética.
Bukasov (1971) y Lechnovich (1971) clasificaron la papa bajo el sistema del ICBN y reconocieron 21 especies, incluyendo por separado las especies S. andigena y S. tuberosum. Dodds (1962) fue el primero en clasificar las papas cultivadas como grupos cultivados, usando las categorías propuestas por el Icncp y reconoció tres especies: S. curtilobum, S. juzepczukii y S. tuberosum; adicionalmente, sugirió que existía un pobre soporte morfológico para la mayoría de las especies cultivadas. Hawkes (1990) reconoció siete especies cultivadas y siete subespecies, mientras que Ochoa (1990 y 1999) lo hizo con nueve especies.
El conteo de cromosomas demuestra que los cultivares con ausencia de dormancia en el tubérculo pueden ser diploides o tetraploides (Ghislain et al., 2006). Esto proporciona una ayuda adicional a la carencia de rasgos coherentes, que mantienen la clasificación de las papas cultivadas como especies diferentes, y apoyan fuertemente la clasificación de las papas cultivadas como grupos diferentes (Ghislain et al., 2006).
Huamán y Spooner (2002), mediante estudios morfológicos combinados con una serie de cruzamientos previos, datos moleculares, y considerando el probable origen híbrido, los orígenes múltiples, la dinámica evolutiva, la hibridación continua y la forma de clasificación, clasificaron todas las papas cultivadas como una sola especie S. tuberosum L., con ocho grupos cultivados (Tab. 1). Adicionalmente, reconocieron los grupos cultivados como apropiados para responder a los propósitos prácticos de los usuarios, pero no como taxas naturales que merezcan el estado de especie.
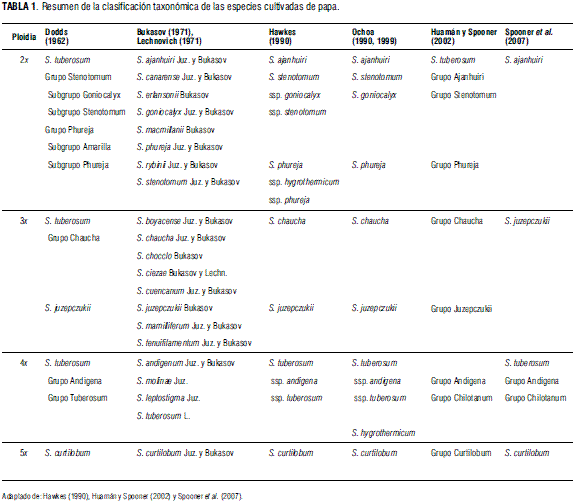
Huamán y Spooner (2002) compararon morfológicamente los cultivares nativos de la papa cultivada -todas las especies cultivadas, excepto los cultivares modernos producidos en tiempos poscolombinos-, utilizando representantes de las siete especies y subespecies propuestas por Hawkes (1990). Estas especies son: S. ajanhuiri, S. chaucha, S. curtilobum, S. juzepczukii, S. phureja ssp. phureja, S. stenotomum ssp. stenotomum, S. stenotomum ssp. goniocalyx, S. tuberosum ssp. andigena y S. tuberosum ssp. tuberosum. Los resultados mostraron algún soporte fenético para las especies S. ajanhuiri, S. chaucha, S. curtilobum, S. juzepczukii y S. tuberosum ssp. tuberosum, pero un pequeño o ningún soporte para los otros seis taxa. Como en las especies silvestres descritas arriba, el mayor soporte morfológico para estas especies fue únicamente por el acompañamiento de sobreposición de algunos caracteres. El conteo de cromosomas no estuvo disponible para Hawkes (1990) y Ochoa (1990), quienes identificaron originalmente estas accesiones y asumieron que todas fueron diploides, una condición incorrecta en algunos casos (Ghislain et al., 2006).
Huamán y Spooner (2002) interpretaron estos resultados a la luz de su probable origen híbrido, orígenes múltiples, dinámica evolutiva por hibridación continua y filosofía en su clasificación, para clasificar a todos los cultivares nativos de las papas cultivadas como una sola especie -S. tuberosum-, con ocho grupos cultivados: Ajanhuiri, Andigena, Chaucha, Chilotanum, Curtilobum, Juzepczukii, Phureja y Stenotomum.
Finalmente, Spooner et al. (2007) clasificaron las papas nativas en cuatro especies: S. ajanhuiri (diploide), S. juzepczukii (triploide), y S. curtilobum (pentaploide) y S. tuberosum L., con dos grupos cultivados: Andigena (Andiegenum) y Chilotanum; en el primero se incluyen todas las papas andinas, tanto diploides como tetraploides, que crecen a lo largo de los Andes desde Venezuela hasta Argentina, y el segundo está conformado por las papas adaptadas a día largo, que crecen en las tierras bajas del centro y sur de Chile, incluyendo las islas del archipiélago de los Chonos (Huamán y Spooner, 2002; Ames y Spooner, 2008; Ghislain et al., 2009).
Conclusiones
La diferencia fundamental entre las formas de clasificación propuestas por el Icncp y el ICBN es que el ICBN utiliza el termino taxón, para denominar a un grupo similar de plantas, mientras que el Icncp usa los términos cultivar y grupo cultivado para clasificar las plantas cultivadas.
Esta diferencia se plantea teniendo en cuenta que las plantas cultivadas se originaron como consecuencia de actividades humanas mediante procesos de selección de manera accidental o deliberada por hibridación y posterior selección en campos de agricultores, y han manteniendo en algunos casos su identidad por sucesivos ciclos de propagación vegetativa o generado nueva variabilidad como consecuencia de la adaptación a diferentes ambientes e hibridación espontánea con especies silvestres o cultivadas.
El análisis de datos a partir de las nuevas tecnologías de la sistemática molecular y el uso de programas avanzados de computador han servido para revisar el conocimiento del germoplasma y definir los límites entre especies y su relación entre ellas. Sin embargo, se mantiene la controversia entre los taxónomos de plantas, debido a un sinnúmero de variantes originadas como consecuencia de factores asociados con eventos de polinización cruzada, autopolinización, apomixis, propagación clonal y poliploidización, lo que favorece diferentes patrones de variación y dificulta una subdivisión en unidades fácilmente reconocibles.
La taxonomía de las plantas cultivadas trae consigo una complicación adicional por la influencia de los factores de domesticación que generan nuevos patrones de variación, debido a que la domesticación favorece mayores y más rápidos efectos sobre caracteres morfológicos usados tradicionalmente en la clasificación de las plantas.
Se hace necesario establecer un acuerdo entre taxónomos para desarrollar un sistema de clasificación estable y único que no solo permita la identificación de los diferentes taxa, sino que facilite la clasificación de grupos de plantas con cualidades similares que sirvan como una herramienta de predicción efectiva para los programas de fitomejoramiento.
Literatura citada
Ames, M. y D.M. Spooner. 2008. DNA from herbarium specimens settles a controversy about origins of the European potato. Amer. J. Bot. 95(2), 252-257. [ Links ]
Andre, C.M., M. Ghislain, P. Bertin, M. Oufir, M.R. Herrera, L. Hoffmann, J.F. Hausman, Y. Larondelle y D. Evers. 2007. Andean potato cultivars (Solanum tuberosum L.) as a source of antioxidant y mineral micronutrients. Agric. Food Chem. 55, 366-378. [ Links ]
Brandenburg, W.A. 1986. Classification of cultivated plants. Acta Hort. 182, 109-115. [ Links ]
Brickell, C.D., B.R. Baum, W.L.A. Hetterscheid, A.C. Leslie, J. McNeill, P. Trehane, F. Vrugtman y J.H. Wiersema. 2004. International code of nomenclature of cultivated plants. Acta Hort. 647, 1-123. [ Links ]
Bukasov, S.M. 1971. Cultivated potato species. pp. 5-40. En: Bukasov, S.M. (ed.). Flora of cultivated plants. Vol. 9. Kolos, Leningrad, Rusia. [ Links ]
Clausen, A.M. y D.M. Spooner. 1998. Molecular support for the hybrid origin of the wild potato species Solanum rechei (Solanum sect. Petota). Crop Sci. 38, 58-865. [ Links ]
Dodds, K.S. 1962. Classification of cultivated potatoes. pp. 517-539. En: Correll, D.S. (ed.). The potato and its wild relatives. Series of Botanical Studies 4. Texas Research Foundation, Renner, TX. [ Links ]
Estrada, N. 2000. La biodiversidad en el mejoramiento genético de la papa. Centro de Información para el desarrollo (CID), Lima, Perú. [ Links ]
FAO. 2009. FAOSTAT. El mundo de la papa. En: http://www.potato2008. org/es/mundo; consulta: noviembre de 2009. [ Links ]
Ghislain, M., D. Andrade, F. Rodríguez, R.J. Hijmans y D.M. Spooner. 2006. Genetic analysis of the cultivated potato Solanum tuberosum L. Phureja Group using RAPDs and nuclear SSRs. Theor. Appl. Genet. 113(8), 1515-1527. [ Links ]
Ghislain, M., J. Núñez, M.R. Herrera y D.M. Spooner. 2009. The single Andigenum origin of Neo-tuberosum-Tuberosum potato materials is not supported by microsatellite and plastidios marker analices. Theor. Appl. Genet. 118, 963-969. [ Links ]
Hawkes, J.G. 1963. A revision of the tuber-bearing Solanums. 2a ed. Scottish Plant Breeding Station Record, Roslin, UK. pp. 76, 181. [ Links ]
Hawkes, J.G. 1990. The potato: evolution, biodiversity and genetic resources. Belhaven Press, Washington DC. [ Links ]
Hennig, W. 1966. Phylogenetic systematic. Univ. Illinois Press, Urbana, IL. [ Links ]
Hetterscheid, W.L.A. y W.A. Brandenberg. 1995. Culton vs. taxon: conceptual issues in cultivated plant systematics. Taxon 44, 161-175. [ Links ]
Hijmans, R., T. Gavrilenko, S. Stephenson, J. Bamberg, A. Salas y D.M. Spooner. 2007. Geographic and environmental range expansion through polyploidy in wild potatoes (Solanum section Petota). Global Ecol. Biogeogr. 16, 485-495. [ Links ]
Hijmans, R.J., M. Jacobs, J.B. Bamberg y D.M. Spooner. 2003. Frost tolerance in wild potato species: assessing the predictivity of taxonomic, geographic, and ecological factors. Euphytica 130, 47-59. [ Links ]
Hijmans, R.J. y D.M. Spooner. 2001. Geographic distribution of wild potato species. Amer. J. Bot. 88, 2101-2112. [ Links ]
Hijmans, R.J., D.M. Spooner, A.R. Salas, A. Guarino y J. de la Cruz. 2002. Atlas of wild potatoes. Systematic and ecogeographic studies on crop gene pools 10(I-IX). International Plant Genetic Resources Institute, Roma. [ Links ]
Huamán, Z. y D.M. Spooner. 2002. Reclassification of landrace populations of cultivated potatoes (Solanum sect. Petota). Amer. J. Bot. 89, 947-965. [ Links ]
Iwanaga, M. y S.J. Peloquin. 1982. Origin and evolution of cultivated tetraploid potatoes via 2n gametes. Theor. Appl. Genet. 61, 161-169. [ Links ]
Knapp, S., L. Bohs, M. Nee y D.M. Spooner. 2004. Solanaceae - a model for linking genomics with biodiversity. Comp. Funct. Genom. 5, 285-291. [ Links ]
Lechnovich, V.S. 1971. Cultivated potato species. pp. 41-304. En: Bukasov, S.M. (ed.). Flora of cultivated plants. Vol. 9. Kolos, Leningrad, Rusia. [ Links ]
Lincoln, R.G., G. Boxshall y P. Clark. 1998. A dictionary of ecology, evolution and systematics. 2aed. Cambridge University Press, Cambridge, MA. [ Links ]
Linnaeus, C. 1753. Species plantarum. Holmiae, Estocolmo. [ Links ]
Mayr, E. 1969. The biologoical meaning of species. Biol. J. Linn. Soc. 1, 311-320. [ Links ]
Miller, J.T. y D.M. Spooner. 1999. Collapse of species boundaries in the wild potato Solanum brevicaule complex (Solanaceae, S. sect. Petota): molecular data. Plant Syst. Evol. 214, 103-130. [ Links ]
Ochoa, C.M. 1990. The potatoes of South America: Bolivia. Cambridge University Press, Cambridge, UK. [ Links ]
Ochoa, C.M. 1999. Las papas de Sudamérica: Pert. CIP, Lima, Perú. [ Links ]
Ochsmann, J. 2004a. Current problems in nomenclature and taxonomy of cultivated plants. Acta Hort. 634, 53-61. [ Links ]
Ochsmann, J. 2004b. Some notes on problems of taxonomy y nomenclature of cultivated plants. Schriften Genet. Res. 22, 43-50. [ Links ]
Sokal, R.R. y T.J. Crovello. 1970. The biological species concept: a critical evaluation. American Naturalist 104, 127-153. [ Links ]
Spooner, D.M., G.J. Anderson y R.K. Jansen. 1993. Chloroplast DNA evidence for the inter relationship of tomatoes, potatoes, and pepinos (Solanaceae). Amer. J. Bot. 80, 676-688. [ Links ]
Spooner, D.M., G.J. Bryan, R.G. van den Berg y A. del Rio. 2003a. Species concepts and relationships in wild and cultivated potatoes. Acta Hort. 619, 63-75. [ Links ]
Spooner, D.M., W.L.A. Hetterscheid, R.G. van den Berg y W. Brandenburg. 2003b. Plant nomenclature and taxonomy: an horticultural and agronomic perspective. Hort. Rev. 28, 1-60. [ Links ]
Spooner, D.M., R.G. van den Berg, G. Bryan y A. del Rio. 2003c. Species concepts and relationships in wild and cultivated potatoes. Acta Hort. 619, 63-75. [ Links ]
Spooner, D.M. y T.R. Castillo. 1997. Reexamination of series relationships of South American wild potatoes (Solanaceae: Solanum Sect. Petota): evidence from chloroplast DNA restriction site variation. Amer. J. Bot. 84, 671-685. [ Links ]
Spooner, D.M., D. Fajardo y G.J. Bryan. 2007. Species limits of Solanum berthaultii Hawkes and S. tarijense Hawkes and the implications for species boundaries in Solanum sect. Petota. Taxon 56(4), 987-999. [ Links ]
Spooner, D.M. y W.L.A. Hetterscheid. 2005. Origins, evolution, and group classification of cultivated potatoes. pp. 285-307. En: Motley, T.J., N. Zerega y H. Cross (eds.). Darwin's harvest: new approaches to the origins, evolution, y conservation of crops. Columbia University Press, NY. [ Links ]
Spooner, D.M. y S. Lara-Cabrera. 2001. Sistemática molecular y evolución de plantas cultivadas. pp. 57-114. En: Hernández, H.M., A. García-Aldrete, F. Álvarez y M. Ulloa (eds.). Enfoques contemporáneos para el estudio de la biodiversidad. Instituto de Biología, UNAM; Fondo de Cultura Económica, México DF. [ Links ]
Spooner, D.M., K. McLean, G. Ramsay, R. Waugh y G. Bryan. 2005b. A single domestication for potato based on multilocus amplified fragment length polymorphism genotyping. Proc. Natl. Acad. Sci. 102, 14694-14699. [ Links ]
Spooner, D.M., J. Núñez, G. Trujillo, M. del R. Herrera, F. Guzmán y M. Ghislain. 2007. Extensive simple sequence repeat genotyping of potato landraces supports a major reevaluation of their gene pool structure and classification. PNAS 104(49), 19398-19403. [ Links ]
Spooner, D., F. Rodríguez, Z. Polgár, H.E. Ballard y S.H. Jansky. 2008. Genomic origins of potato polyploids: GBSSI gene sequencing data. The plant genome. A Supplement to Crop Science 48(S1), S27-S36. [ Links ]
Spooner, D.M. y A. Salas. 2006. Structure, biosystematics, and genetic resources. pp. 1-39. En: Gopal, J. y S.M.P. Khurana (eds.). Handbook of potato production, improvement, and postharvest management. Haworth's Press, Inc., Binghampton, NY. [ Links ]
Spooner, D.M. y K.J. Sytsma. 1992. Reexamination of series relationships of Mexican and Central American wild potatoes (Solanum sect. Petota): evidence from chloroplast DNA restriction site variation. Syst. Bot. 17, 432-448. [ Links ]
Spooner, D.M. y R.G. van den Berg. 1992. An analysis of recent taxonomic concepts in wild potatoes (Solanum section Petota). Genet. Res. Crop Evol. 39, 23-37. [ Links ]
Spooner, D.M., R.G. van den Berg, A. Rodriguez, J. Bamberg, R.J. Hijmans y S. Lara-Cabrera. 2004. Wild potatoes (Solanum section Petota) of North and Central America. Syst. Bot. Monogr. 68, 1-209. [ Links ]
Spooner, D.M., R.R. van Treuren y M.C. de Vicente. 2005a. Molecular markers for germplasm y genebank management. Techn. Bull. 10. International Plant Genetic Resources Institute, Roma. [ Links ]
Stuessy, T.F. 1990. Plant Taxonomy: the systematic evaluation of comparative data. Columbia University Press, New York, NY. [ Links ]
Sukhotu, T. y K. Hosaka. 2006. Origin and evolution of Andigena potatoes revealed by chloroplast y nuclear dna markers. Genome 49, 636-647. [ Links ]
Sukhotu, T., O. Kamijima y K. Hosaka. 2006. Chloroplast dna variation in the most primitive cultivated diploid potato species Solanum stenotomum Juz. et Buk. and its putative wild ancestral species using high-resolution markers. Genet. Resour. Crop Evol. 53, 53-63. [ Links ]
Ugent, D. 1970. The potato: what is the origin of this important crop plant, and how did it first become domesticated? Sci. 170, 1161-1166. [ Links ]
Van den Berg, R.G., J.T.Miller, M.L. Ugarte, J.P. Kardolus, J. Nienhuis y D.M. Spooner. 1998. Collapse of morphological species in the wild potato Solanun Brevicaule complex (Solanaceae: sect. Petota). Amer. J. Bot. 85, 92-109. [ Links ]
Watanabe, K. y S.J. Peloquin. 1989. Ocurrence of 2n pollen and ps gene frequencies in cultivated groups and their related wild species in tuber-bering Solanums. Theor. Appl. Genet. 78, 329-336. [ Links ]
Werner, J.E. y S.J. Peloquin. 1991. Occurrence and mechanisms of 2n egg formation in 2x potato. Genome 34, 975-982. [ Links ]

