










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.27 n.3 Bogotá set./dez. 2009
Adriana Tofiño1, 6, Diana Cabal2, Hernán Ceballos3, Juan Carlos Pérez4 y Hernán Mauricio Romero5
Artículo derivado de tesis doctoral en Ciencias Agrarias de la primera de los autores.
1 Centro de Investigación Motilonia, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Codazzi (Colombia).
2 Departamento de Agroindustriales, Facultad de Ingenierias y Tecnológicas, Universidad Popular del Cesar, Valledupar (Colombia).
3 Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Palmira.
4 Programa de Mejoramiento de Yuca, Centro Internacional de Agricultura Tropical (CIAT), Palmira (Colombia).
5 Departamento de Biología, Facultad de Ciencias, Universidad Nacional de Colombia, Bogotá.
6 Autor de correspondencia. atofino@corpoica.org.co
Fecha de recepción: 12 de abril de 2009. Aceptado para publicación: 6 de noviembre de 2009
RESUMEN
En el marco de la creciente tendencia a la agroindustrialización de la yuca, y dado que las características del almidón definen el potencial industrial de estas raíces, aquellas herramientas moleculares que permitan identificar variantes de interés deben integrarse a programas de mejoramiento genético para seleccionar eficientemente nuevos materiales parentales. En este trabajo se compararon los resultados de la técnica TILLING de genética reversa (modificada en cuanto a la visualización directa de los productos de digestión en gel de agarosa), con el método tradicional que utiliza gel de acrilamida y plataforma Li-Cor. La comparación se llevó a cabo a través de la evaluación de polimorfismo en 10 genes con control metabólico mayor de la ruta del almidón. En 150 líneas endocriadas M2, derivadas de semillas irradiadas, fueron identificadas fenotípicamente como probables mutantes y analizadas por TILLING. Solo 30% de los cebadores utilizados produjo datos de buena calidad pues la mayoría amplificó más de un blanco. Adicionalmente, la detección visual de SNP en agarosa no corresponde con los registros obtenidos a partir del genotipado por Li-Cor, pues con ella no se logró detección de SNP, ni en plantillas de mezclas de ADN, ni en muestras individuales.
Palabras clave: síntesis de cebadores, polimorfismo en nucleótidos individuales (SNP), genética reversa, mutación inducida, calidad de almidón, plataformas de genotipado.
ABSTRACT
In the context set by the continuously growing agroindustrialization of cassava, and given that starch traits define tuber industrial potential, molecular tools that allow identifying variants of interest should be integrated into breeding programs for efficient selection of parental materials. The present research study compared the TILLING reverse genetics technique (modified by directly visualizing cleaved products in agarose gel) to the traditional acrylamide gel and Li-Cor platform method. Both techniques were applied to polymorphism evaluation in 10 genes with major control of the starch metabolic route. A group of 150 M2 inbred lines derived from irradiated germplasm were phenotypically identified as probable mutants and analyzed by TILLING. Only 30% of the primers used produced good quality data, for most of them amplified more than one target. Additionally, visual detection of SNPs in agarose was not achieved in mixtures of DNA templates nor in individual samples, so the results cannot be said to correspond with those obtained by Li-Cor genotyping.
Key words: primer synthesis, Single Nucleotide Polymorphism (SNP), reverse genetics, induced mutations, starch quality, genotyping platform.
Introducción
En países tropicales y subtropicales, los recursos amiláceos más importantes son la palma sago, papa y yuca (Rickard et al., 1991; Moorthy, 2004). La yuca, mandioca, tapioca, Manihot esculenta Crantz, se siembra en tierras bajas de Asia, África y Suramérica por sus raíces amiláceas cuyo contenido fluctúa entre 73,7 y 84,9% del peso seco total (Ceballos et al., 2006).
En la actualidad se diversificó su utilización, pues, además del consumo fresco, alimentación animal, comidas procesadas y producción de bioetanol, las raíces ingresaron en el mercado global del almidón y sus derivados (Tofiño et al., 2007). Los procesadores de yuca, requieren almidones novedosos como cambios en proporción de amilosa, tamaño, forma y composición química de gránulos, para mejorar la competitividad y disminuir el impacto ambiental de los procesos industriales. Este es el caso de la producción de alcohol carburante en la que se requieren raíces con mayor conversión por unidad de biomasa, relacionada con contenido alto de amilosa.
En este sentido, una limitante del aprovechamiento industrial es el desconocimiento de la variabilidad genética del almidón, ya que la alta heterocigosidad de la especie dificulta la identificación de variantes alélicas de interés (Ceballos et al., 2004; Ceballos et al., 2006). El avance en la comprensión de la estructura genética del cultivo y la secuenciación completa del genoma posibilitarán análisis de rasgos complejos en el acervo genético y posterior introgresión en variedades comerciales mediante ingeniería genética, mejoramiento convencional y mutagénesis (Tofiño et al., 2007).
Adicionalmente, se ha difundido la mutagénesis debido al acople entre tecnologías de secuenciamiento de bajo costo con colecciones de secuencias expresables (EST) y estrategias de genética reversa como TILLING (Bouck y Vision, 2007). Esta técnica deriva su nombre de las siglas (Targeting Induced Local Lesions in Genomes). Este método, desarrollado en Arabidopsis, consiste en la inducción e identificación de mutaciones puntuales en genes específicos, y se emplea exitosamente en especies animales y vegetales (Sood et al., 2006; Wang et al., 2006; Cooper et al., 2008). Generalmente, en TILLING se utilizan mutagénicos químicos como etilmetanosulfonato (EMS) que presenta efectos alquilantes sobre el ADN como transiciones G:C a A:T. Sin embargo, se registran investigaciones con radiaciones ionizantes como rayos gamma y neutrones rápidos, que producen sustituciones de un nucleótido, deleciones de 2-4 pb y de más de 6 Mb, no transmisibles a la progenie.
Aunque la tasa de mutación observada depende de la especie y el órgano expuesto, se ha sugerido que la densidad de mutaciones puntuales producidas por EMS es superior a gamma, mientras que las mutaciones inactivantes de genes "Knockout" generados por gamma, son superiores a las de EMS (Ahloowalia et al., 2004; Wu et al., 2005; Otani et al., 2006; Sato et al., 2006; Tofiño et al., 2007; Xin et al., 2008). Datos más precisos corroboran esta tendencia, pues se ha registrado con gamma densidades de mutación de 1/6190 kb en arroz; mientras que EMS produce 1/526 kb y, en general, mayores densidades de mutación en la mayoría de cultivos (Till et al., 2007); en Arabidopsis 1/300 kb (Greene et al., 2003), soya 1/140 kb (Cooper et al., 2008) y sorgo 1/526 kb (Xin et al., 2008).
El TILLING se adaptó para la identificación de diversidad natural mediante EcoTILLING (Wang et al., 2006). Ambas técnicas, acopladas a herramientas de haplotipado, se utilizan en mejoramiento genético, identificación de variación alélica en genes de expresión correlacionable con fenotipos, estudios de caracterización de germoplasma, identificación de genes candidatos y series alélicas en loci de diferentes rasgos; robustecimiento de bases de estudio en familias de genes, mapeo en plantas diploides y poliploides (Weil, 2005; Wang et al., 2006; Wang et al., 2007; Balvan et al., 2008; Cooper et al., 2008).
El TILLING aprovecha la actividad endonucleasa de CEL I, que digiere heterodúplex en posiciones de un solo nucleótido o bucles de inserciones pequeñas debido a mutaciones (Gilchrist y Haughn, 2005; Wang et al., 2007). En la detección de polimorfismos de un solo nucleótido (SNP) dentro de una población, se generan mezclas de ADN en las que el locus blanco se amplifica mediante PCR, se somete a la formación de heterodúplex y al corte con CEL I. Recientemente, además de CEL I se identificaron, en Brassica y Arabidopsis, endonucleasas con mayor especificidad de corte (Sato et al., 2006; Triques et al., 2007).
Actualmente, la electroforesis en poliacrilamida acoplada al uso de plataformas de genotipificación de alta resolución como Li-Cor constituye el método usual para identificar productos de digestión. El uso masivo del TILLING en países del tercer mundo depende de la detección de productos de digestión en gel de agarosa, sin genotipado por Li-Cor o marcaje de cebadores con fluorescencia (Raghavan et al., 2007).
En respuesta a las demandas de la industria almidonera, se planteó un proyecto de mejoramiento basado en mutagénesis, cuyo objetivo es producir e identificar variables de interés en la calidad de raíces, especialmente del almidón. Sin embargo, no existen reportes de TILLING en germoplasma de yuca, y el desarrollo de la técnica es fundamental para el análisis rápido de secuencias de interés en poblaciones mutantes grandes. El programa de mejoramiento de yuca de CIAT trabajó en la estandarización de la técnica TILLING para validarla como alternativa eficiente, barata y aplicable en países productores, en la identificación de mutantes comercialmente útiles, en genes de la ruta del almidón. Esta iniciativa cobró relevancia, pues la identificación del mutante de gránulo pequeño (Ceballos et al., 2008); otros mutantes del almidón aún no registrados, como gránulo hueco, bajo almidón, resistencia al deterioro fisiológico y posible apomíxis, se lograron con técnicas microscópicas acopladas a pruebas fisicoquímicas, en una población M2 pequeña cuya inversión en tiempo, recursos humanos y financieros ha sido considerable. Por tanto, la sostenibilidad de la mutagénesis en yuca dependerá de la estandarización e implementación del TILLING.
Materiales y métodos
Material vegetal
En 2003, se irradiaron con rayos γ de 200 Gy (a partir de cobalto 60) 1.400 semillas de cinco familias de hermanos completos y medios hermanos; los parentales se referencian en la Tab. 1. Las plántulas M1 se trasplantaron al campo, y en 2004 inició la floración. Seguidamente, se realizó autopolinización en plantas M1, para obtener semillas M2. Las plantas M2 se trasplantaron en 2005, y se cosecharon en 2006.
Evaluación plantas M2 para selección del subgrupo a analizar por TILLING
Se realizaron evaluaciones fenotípicas para seleccionar posibles mutantes, mediante identificación en M2, de quimeras registradas en M1. Esta información se complementó con microscopía óptica para identificar cambios en la forma y el tamaño del gránulo y pruebas de tinción con yodo para identificar cambios importantes en el contenido de amilosa.
A partir de los resultados, se identificaron 150 genotipos para análisis TILLING modificado, por presentar características inusuales en variables de calidad de raíces con respecto a valores máximos y mínimos registrados en análisis del banco de germoplasma de yuca, que incluyen los parentales no irradiados (CIAT, 2005). Dentro de ese grupo, 30 individuos con alteraciones del gránulo también se evaluaron por TILLING convencional (Tab. 1).
Extracción de ADN y diseño de cebadores
Se extrajo ADN de 150 individuos M2, parentales M1, individuos no irradiados de las familias evaluadas, genotipos control como TAI 8, en el que se han evaluado un gran número de marcadores SSR y yuca Waxy. En la Tab. 1 se relacionan 30 genotipos de este subgrupo enviados a IAEA para el desarrollo del método estándar de TILLING, con electroforesis en acrilamida, lectura por Li-Cor y secuenciación de productos de amplificación de cebadores. Se diseñaron y sintetizaron cebadores a partir de secuencias disponibles de 10 enzimas relacionadas con la biosíntesis del almidón en especies de plantas superiores, disponibles en la web del Genebank como Sintetasa del almidón ligada al gránulo I (GBSSI), con tamaño variable entre especies desde 2.800 a 3.572 pb; enzima ramificadora del almidón II (SBEII); enzima deramificadora del almidón II (DBEII); sacarosa sintetasa (SUSY); sintetasa del almidón soluble II (SSII); sacarosa fosfato sintetasa (SPS); Invertasa; transportador de sacarosa 1 (SUT1) y transportador de maltosa1 (MEX1). Las enzimas se seleccionaron por la tasa de control elevada sobre el flujo metabólico de la ruta (Tetlow et al., 2004) (Tab. 2).

Posteriormente, se comparó la similitud entre secuencias de enzimas conocidas con secuencias expresadas EST de la genoteca de yuca, disponibles en el sitio web de CIAT y el Institute of Chinese Academy of Tropical Agricultural Science. Una vez identificadas cuáles secuencias ortólogas del gen de interés en las plantas consideradas presentaban mayor homología con fragmentos EST de yuca, se procedió a diseñar cebadores, con apoyo del programa Coddle (Greene et al., 2003) y Primer3 (Rozen y Skaletsky, 2000). En la extracción de ADN se utilizó tejido foliar y el método de mini extracción Dellaporta, modificado. La calidad de ADN se verificó en agarosa al 3%, tinción con bromuro de etidio y cuantificación de concentración mediante fluorómetro.
Desarrollo del TILLING
La PCR se desarrolló en volumen de 20 μL consistente en dH2O, 1 × PCR buffer (Promega Corp., Madison, WI), 1,5 mM MgCl2 (Promega Corp.), 0,2 mM dNTPs (Promega Corp.), 0,0625 U Taq polymerase (Promega Corp.), 0,2 μM de oligonucleótidos en sentido y antisentido sin marcar con fluorescencia (Operon Biotechnologies, Inc., Huntsville, AL), 3,125 mg mL-1 PVP, 0,125 mg mL-1 BSA (New England BioLabs, Ipswich, MA), y 0,875 - 1,25 ng de ADN.
Las condiciones del termociclador fueron 95°C por 5 min para desnaturalización inicial, 40 ciclos de 95°C por 1 min, 63-69°C por 1 min, 72°C por 1 min, un ciclo de 72°C por 10 min y 4°C. Los productos PCR se visualizaron en agarosa al 3% con tinción con bromuro de etidio. Los productos PCR se calentaron y enfriaron para formar heterodúplex a través de un ciclo de 99°C por 10 min para inactivar la Taq polimerasa, 70 ciclos de 20 s a partir de 70°C disminuyendo 0,3°C. Una vez formados los heterodúplex, los productos PCR se trataron con CEL I (Transgenomic, Inc. Omaha, NE) y limpieza para evitar el exceso de primer/dimer con polietilenglicol y etanol 250 μL. Los productos de digestión se incubaron a 42°C por 20 min, para digestión de secuencias de baja alineación en los bucles. Se aplicó solución de parada (Transgenomic, Inc.) y se visualizaron los productos de digestión en geles de agarosa de 1,5 y 3,0%.
Resultados y discusión
La técnica TILLING se caracteriza por su exigencia elevada en calidad del ADN molde y cebadores empleados en amplificación por PCR (Till et al., 2007). Se sintetizaron cebadores de genes de nueve enzimas del almidón y tres pares para amplificar GBSSI, debido a su gran tamaño. Posteriormente, se estandarizaron las condiciones de amplificación por PCR en todos los genes. En la Tab. 2 se relaciona la lista de genes evaluados y sus respectivos cebadores para TILLING. Se evaluó la amplificación de cebadores sintetizados en germoplasma mutante y se realizaron ajustes para garantizar amplificación en cada uno de ellos.
En la Fig. 1 se presentan productos de amplificación por PCR en condiciones estandarizadas. La estandarización de la técnica en germoplasma de yuca se inició con la evaluación de cebadores del inicio, medio y final de GBSSI, para verificar calidad de amplificación en los individuos CRA- 013, CW14-11, MCOL-1508 y MCOL-2269. Se observan bandas bien definidas e intensas en la amplificación de cada cebador.

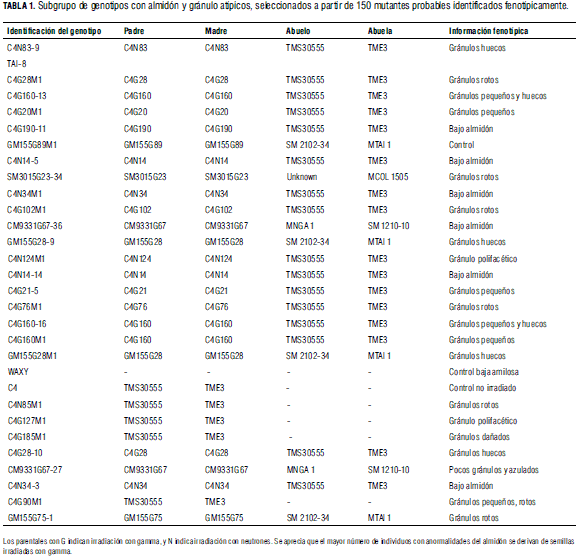
Una vez realizados los ajustes a las especificaciones del kit de invitrogen, se verificó la estandarización de la técnica en germoplasma de yuca en GBSSI. En la Fig. 2 se presentan los resultados de la hibridación, considerando diferentes controles del kit, genotipos cultivados y silvestres de yuca. Se puede observar la formación de heterodúplex tanto en los controles como en los individuos de yuca.
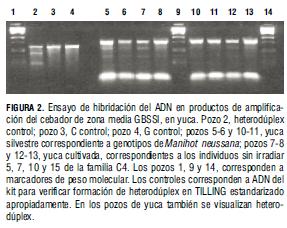
Se identificó en la población M2, mediante caracterización fenotípica, mutantes de características del almidón como gránulo pequeño (Ceballos et al., 2008). Igualmente, la caracterización fenotípica de M2 permitió identificar individuos con otras anormalidades del gránulo y, por tanto, con probabilidad elevada de polimorfismo en genes del almidón. A partir de este grupo, se evaluó el sistema TILLING modificado, pues la probabilidad de formación de bucles detectables por CEL I era mayor. Uno de los aspectos sensibles del TILLING fue la obtención de ADN de alta calidad. En este sentido, se evaluó la amplificación de cebadores sintetizados en mutantes putativos (Tab. 2). Posterior a la verificación de la calidad del ADN, se evaluó la amplificación de cebadores de genes de interés, previamente a la aplicación del TILLING. Durante la aplicación de la técnica en los genes seleccionados, se observó ausencia de productos de digestión, indicadores de cambio en la secuencia de ADN (Fig. 3).
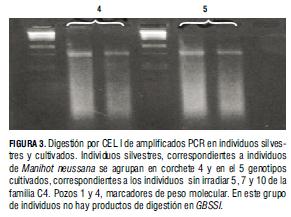
Entonces, se diseñó un ensayo para definir si la ausencia de polimorfismo en genes de las enzimas de la ruta biosintética del almidón se debía a carencia de mutaciones, anomalías en diseño de cebadores o número bajo de individuos evaluados (Fig. 4). En la figura se aprecia la amplificación del ADN de genotipos M2 con cuatro cebadores. Las muestras se ubicaron entre marcadores invitrogen de peso molecular de precisión elevada. Los productos cercanos a 400 pb son apropiados para TILLING; por tanto, el primero y el segundo par de cebadores exhiben productos inapropiados para la técnica (Raghavan et al., 2007).
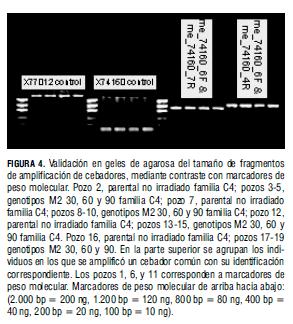
Este ensayo develó la posibilidad de cebadores inapropiados para TILLING; por tanto, se enviaron a la IAEA muestras del germoplasma y las secuencias de cebadores para realizar análisis complementarios que señalaran los aspectos críticos que impidieron el análisis exitoso de mutantes de yuca mediante el protocolo modificado de TILLING (Tab. 1).
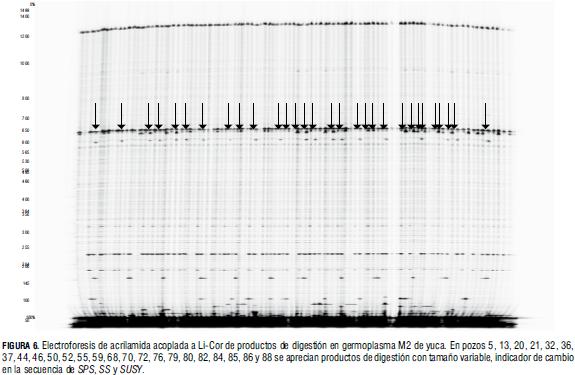
En la IAEA se desarrollaron análisis de calidad de ADN, diseño de cebadores y productos de amplificación, mediante pruebas de secuencia de amplificaciones y validación de productos de digestión por Li-Cor (Figs. 5 y 6). Esta metodología se utilizó en la identificación de cebadores que amplificaron más de un blanco y ocasionaron ineficacia en la identificación de mutaciones con TILLING en soya.
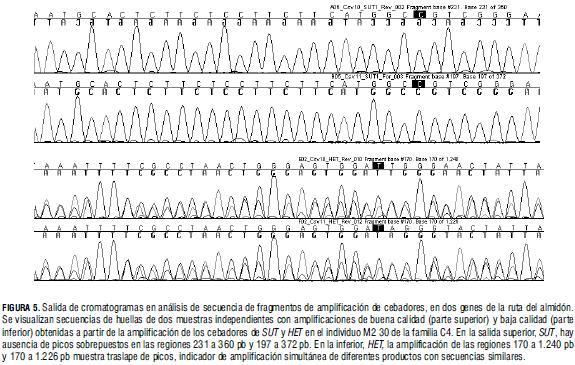
En soya, se evitó redundancia de blancos con la aplicación de pretratamiento del ADN total con una enzima de restricción (Cooper et al., 2008). La superposición de picos, en la salida inferior de cromatogramas de secuencias amplificadas en yuca, surgió debido a la amplificación por el cebador de diferentes fragmentos en el gen objetivo (Fig. 5).
Los resultados indican que únicamente cebadores de SPS, SS III, 4F/1R de SS II y F/7R de SUSY presentaron la especificidad en amplificación requerida por TILLING. Los otros cebadores analizados son inespecíficos. Los datos obtenidos muestran eficiencia de 30% en la síntesis de cebadores. Resultados similares se identificaron en soya, donde se alcanzó 37% de efectividad en síntesis de cebadores con alta especificidad (Cooper et al., 2008). Este problema podría estar asociado con genes blanco con copias cercanas, lo cual es típico en especies poliploides o miembros de familias de genes. Lo anterior, en concordancia con el conocimiento actual de la evolución del cultivo y de las características genéticas de algunas enzimas de la ruta biosintética del almidón en yuca, como SBE, GBSSI y SS. Específicamente, en SS y GBSS, se han identificado dos isoformas, y en DBE cuatro (Tetlow et al., 2004; Sakurai, et al., 2007; Cooper et al., 2008). La secuenciación completa del genoma de las especies facilitaría la definición de cebadores específicos para un solo gen, pues el conocimiento de las secuencias de genes homólogos encaminaría el diseño de cebadores hacia regiones menos conservadas o identificación de enzimas de restricción que solo detecten uno de los homólogos, evitando redundancia de blancos (Cooper et al., 2008).
En aquellas especies que no cuentan con esa facilidad, se hace necesaria la implementación de pruebas previas de amplificación en geles de agarosa y posterior secuenciación para eliminar cebadores inespecíficos y así mejorar la eficiencia en la detección de mutaciones (Slade et al., 2005). En este sentido, se han presentado iniciativas importantes para alcanzar la secuenciación completa de yuca, como la planteada por el Departamento de Energía en asocio con el Genome Institute de Estados Unidos (Sakurai et al., 2007).
Algunas técnicas moleculares más simples, como microsatélites (SSR) o conformación en la secuencia de hebras simples de ADN (SSCP), se han utilizado con éxito en yuca en selección asistida por marcadores y mapeo genético con alta eficiencia, bajo costo y requerimientos mínimos de laboratorio, por lo cual se han convertido en herramientas clave en investigación en países en vías de desarrollo (Fregene et al., 2001; Okogbenin et al., 2006; Ayeh et al., 2008). Sin embargo, estas estrategias moleculares no alcanzan la eficiencia del TILLING en la identificación de mutantes inducidos en genes específicos. La efectividad alta en detección de SNP por TILLING ha estado apoyada en el uso de primers marcados con fluorescencia, electroforesis en geles de acrilamida y acople al Li-Cor para detección de productos de digestión (Greene et al., 2003; Triques et al., 2007; Till et al., 2007; Cooper et al., 2008; Xin et al., 2008).
Las pruebas realizadas en Li-Cor con los cebadores diseñados apropiadamente, sugieren que la calidad del ADN mutante es adecuada para la técnica, pues al aplicar TILLING bajo el protocolo convencional en 17 genotipos mutantes, se observaron productos de digestión de tamaño diferente (Fig. 6).
De acuerdo con lo anterior, el análisis directo del gel de agarosa resultó ineficiente, pues los individuos evaluados con Li-Cor en IAEA se habían analizado previamente en agarosa, sin identificación de productos de digestión. El sobrecosto de la electroforesis de acrilamida disminuye las posibilidades de financiación del análisis de más de 10.000 individuos, como se reporta en proyectos de mutagénesis en otras especies (Balvan et al., 2008).
Por otro lado, a diferencia de yuca, en arroz se evaluó exitosamente la detección de SNP en agarosa mediante TILLING (Raghavan et al., 2007). En arroz, la modificación de TILLING mostró eficiencia en detección de productos de digestión, similar a la exhibida con genotipificación por Li-Cor; con nivel de resolución en productos de digestión hasta de 3 kb en mezclas de ADN de ocho individuos. La disponibilidad de la secuenciación completa del genoma de arroz favorece la síntesis de cebadores específicos aun para amplificación de isoformas, comunes en enzimas de la ruta del almidón (Balvan et al., 2008). Por tanto, el desarrollo de esta técnica compleja en una especie cuya arquitectura genética no se conoce plenamente es un gran reto.
Conclusiones
Los avances obtenidos muestran las posibilidades del TILLING para evaluar germoplasma mutante de yuca. Sin embargo, se requieren técnicas complementarias, apoyo de avances en la secuenciación y saturación EST para desarrollar eficientemente la técnica, y así implementar proyectos de mejoramiento basado en mutagénesis inducida en países en vías de desarrollo. En mayor detalle, se observó que la metodología de extracción de ADN aplicada es adecuada para el TILLING. De igual forma, los resultados indican que el sistema Li-Cor es fundamental especialmente para visualizar productos de digestión en genes con isoformas en yuca. Adicionalmente, debido a la complejidad de la arquitectura genética del cultivo, los estándares en síntesis de cebadores son exigentes y demandan pruebas de amplificación y secuenciación complementarias al uso de Coddle y primer3 en el diseño de cebadores. Así mismo, para optimizar el TILLING en yuca se requiere la utilización de endonucleasas con mayor especificidad que CEL I o que reconozcan secuencias específicas de genes homólogos. También se evidenció que gamma es fuente de mutación apropiada para yuca, pues el mayor porcentaje de individuos seleccionados para análisis con TILLING y los mutantes identificados por Li-Cor en IAEA descienden de semillas irradiadas con gamma. Finalmente, es importante precisar que una limitante en el análisis de resultados de la técnica en yuca está asociada con el efecto de la reproducción sexual y la endogamia en M2, sobre el genoma heterocigoto, lo cual permite la expresión de genes recesivos, no expresados durante la reproducción vegetativa; por tanto en TILLING no puede diferenciarse si los mutantes identificados corresponden a variaciones por alelos múltiples recesivos o al efecto de la mutación inducida en loci de interés.
Literatura citada
Ahloowalia, B., M. Maluszynski y K. Nichterlein. 2004. Global impact of mutation derived varieties. Euphytica 135, 187-204. [ Links ]
Ayeh, K.O. 2008. Expressed sequence tags (ESTs) and single nucleotide polymorphisms (SNPs): merging molecular marker tools for improving agronomic traits in plant biotechnology. Afr. J. Biotechnol. 7(4), 331-341. [ Links ]
Balvan, N., O. Sreenivasulu., P. Riera y S. Azhaguvel. 2008. Mutagenesis and high-throughput functional genomics in cereal crops: current status. Adv. Agron. 98, 357-417. [ Links ]
Bhat, R.S., N.M. Upadhyaya, A. Chaudhury, Ch. Raghavan, F. Qiu, H. Wang, J. Wu, K. McNally, H. Leung, B. Till, S. Henikoff y L. Comai. 2007. Chemical- and irradiation-induced mutants and TILLING. pp. 148-180. En: Upadhyaya, N.M. (ed.). Rice functional genomics challenges, Progress and prospects. Springer, New York, NY. [ Links ]
Bouck, A. y T. Vision. 2007. The molecular ecologist's guide to expressed sequence tags. Mol. Ecol. 16(5), 907-924. [ Links ]
Ceballos, H., M.A. Fregene, Z. Lentini, T. Sánchez, Y.J. Puentes P., J.C. Pérez V., A. Rosero y A.P. Tofiño. 2006. Development and identification of high-value cassava clones. Acta Hort. 703, 63-70. [ Links ]
Ceballos, H., C. Iglesias, J. Pérez y A. Dixon. 2004. Cassava breeding: opportunities and challenges. Plant Mol. Biol. 56(4), 503-516. [ Links ]
Ceballos, H., T. Sánchez, K. Denyer, A.P. Tofiño, E.A. Rosero, D. Dufour, A. Smith, N. Morante, J.C. Pérez y B. Fahy. 2008. Induction and identification of a small-granule, high-amylose mutant in cassava (Manihot esculenta Crantz). J. Agric. Food Chem. 56 (16), 7215-7222. [ Links ]
CIAT. 2005. Project IP3, Improved cassava for the developing world. Annual Report. Centro Internacional de Agricultura, Cali, Colombia. [ Links ]
Cooper, J.L., B.J. Till, R.G. Laport, M.C. Darlow, J.M. Kleffner, A. Jamai, T. El-Mellouki, S. Liu, R. Ritchie, N. Nielsen, K.D. Bilyeu, K. Meksem, L. Comai y S. Henikoff. 2008. TILLING to detect induced mutations in soybean. BMC Plant Biol. 8, 9. [ Links ]
Fregene, M., E. Okogbenin, C. Mba, F. Ángel, M. Suárez, J. Gutiérrez, P. Chavarriaga, W. Roca, M. Bonierbale y J. Tohme. 2001. Genome mapping in cassava improvement: challenges, achievements and opportunities. Euphytica 120(1), 159-165. [ Links ]
Gilchrist, E.J. y G.W. Haughn. 2005. TILLING without a plough: a new method with applications for reverse genetics. Curr. Opin. Plant Biol. 8(2), 211-215. [ Links ]
Greene, E.A., C.A. Codomo, N.E. Taylor, J.G. Henikoff, B.J. Till, S.H. Reynolds, L.C. Enns, C. Burtner, J.E. Johnson, A.R. Odden, L. Comai y S. Henikoff. 2003. Spectrum of chemically induced mutations from a large-scale reverse-genetic screen in Arabidopsis. Genetics 164, 731-740. [ Links ]
Moorthy, S.N. 2004. Tropical sources of starch. pp. 321-359. En: Eliasson, A.C. (ed.). Starch in food. CRCPress, Boca Ratón, FL. [ Links ]
Okogbenin, E., J. Marín y M. Fregene. 2006. An SSR-based molecular genetic map of cassava. Euphityca 147(3), 433-440. [ Links ]
Otani, M., H. Saito, T. Abe y T. Shimada. 2006. Induction of mutations in sweetpotato plants by heavy-ion beam irradiation. Acta Hort. 703, 171-174. [ Links ]
Rickard, J.E., M. Asaoka y J.M.V. Blanshard. 1991. The physicochemical properties of cassava starch. Trop. Sci. 31, 189-207. [ Links ]
Rozen, S. y H. Skaletsky. 2000. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 132, 365-386. [ Links ]
Sakurai, T., G. Plata, F. Rodríguez-Zapata, M. Seki, A. Salcedo, A. Toyoda, A. Ishiwata, J. Tohme, Y. Sakaki, K. Shinozaki y M. Ishitani. 2007. Sequencing analysis of 20,000 full-length cDNA clones from cassava reveals lineage specific expansions in gene families related to stress response. BMC Plant Biol. 7, 66. [ Links ]
Sato, Y., K. Shirasawa, Y. Takahashi, M. Nishimura y T. Nishio. 2006. Mutant selection from progeny of gamma-ray -irradiated rice by DNA heteroduplex cleavage using Brassica petiole extract. Breed. Sci. 56, 179-186. [ Links ]
Slade, A.J., S.I. Fuerstenberg, D. Loeffler, M.N. Steine y D. Facciotti. 2005. A reverse genetic, nontransgenic approach to wheat crop improvement by TILLING. Nat. Biotechnol. 23(1), 75-81. [ Links ]
Sood, R., M.A. English, M. Jones, J. Mullikin, D.M. Wang, M. Anderson, D. Wu, S.C. Chandrasekharappa, J. Yu, J. Zhang y P. Liu P. 2006. Methods for reverse genetic screening in zebrafish by resequencing and TILLING. Methods 39(3), 220-227. [ Links ]
Tetlow, I., M.K. Morell y M.J. Emes. 2004. Recent developments in understanding the regulation of starch metabolism in higher plants. J. Exp. Bot. 55(406), 2131-2145. [ Links ]
Till, B.J., J. Cooper, T.H. Tai, P. Colowit, E.A. Greene, S. Henikoff y L. Comai. 2007. Discovery of chemically induced mutations in rice by TILLING. BMC Plant Biol. 7, 19. [ Links ]
Tofiño R., A.P., H.M. Romero, M.A. Fregene y A. Rosero. 2007. Posibilidades y alcances del mejoramiento genético basado en mutación inducida en yuca (Manihot esculenta Crantz). Fitot. Colomb. 7(1), 1-12. [ Links ]
Triques, K., B. Sturbois, S. Gallais, M. Dalmais, S. Chauvin, C. Clepet, S. Aubourg, C. Wang, Z.-X. Sun e Y.-Z. Tao. 2006. Aplication of TILLING in plant improvement. Acta Genetica Sinica 33(11), 957-964. [ Links ]
Wang, G.X., M.K. Tan, S. Rakshit, H. Saitoh, R. Terauchi, T. Imaizumi, T. Ohsako y T. Tominaga. 2007. Discovery of single-nucleotide mutations in acetolactate synthase gene by Ecotilling. Pest. Biochem. Physiol. 88(2), 143-148. [ Links ]
Weil, C. 2005. Single base hits score a home run in wheat. Trends Biotechnol. 23(5), 220-222. [ Links ]
Wu, J.-L., C. Wu, C. Lei, M. Baraoidan, A. Bordeos, M.R.S. Madamba, M. Ramos-Pamplona, R., Mauleon, A. Portugal, V.J. Ulat, R. Bruskiewich, G. Wang, J. Leach, G. Khush y H. Leung. 2005. Chemical- and irradiation-induced mutants of indica rice IR64 for forward and reverse genetics. Plant Mol. Biol. 59(1), 85-97. [ Links ]
Xin, Z., M.L. Wang, N.A. Barkley, G. Burow, C. Franks, G. Pederson y J. Burke. 2008. Applying genotyping (TILLING) and phenotyping analyses to elucidate gene function in a chemically induced sorghum mutant population. BMC Plant Biol. 8, 103. [ Links ]

