










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.27 n.3 Bogotá set./dez. 2009
Gustavo A. Ligarreto M.1, 2 y Andrés R. Ospina H.1
1 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
2 Autor de correspondencia. galigarretom@unal.edu.co>
Fecha de recepción: 19 de febrero de 2009. Aceptado para publicación: 6 de noviembre de 2009
RESUMEN
En el mejoramiento del cultivo de arveja es importante estimar los factores heredables que, asociados al rendimiento y la precocidad de los diferentes materiales, tienden a incrementar la productividad. Para tal efecto, en la Sabana de Bogotá se realizó el presente estudio durante un ciclo de producción en condiciones de invernadero. Se emplearon 10 materiales de arveja en diferentes generaciones filiales, usando como parentales los cultivares Santa Isabel (SI), San Isidro (SAI), Diacol Caldas (DC) y New Era Wisconsin (NEW). Los caracteres evaluados fueron días en la floración, nudo donde aparece la primera flor, altura de la planta a floración, días a fructificación, ancho de vaina, longitud de vaina, número de vainas por planta, número de semillas por vaina, rendimiento por planta y peso de 100 semillas; todos los cuales fueron analizados por comparación de medias. Para cada variable se estimó el promedio de los parentales y sus respectivas progenies F1 y F2. Luego se calculó la heredabilidad por el método Warner. Los cuatro parentales, junto con el híbrido SIxSAI F1, presentaron los menores rendimientos, mientras que los híbridos SIxDC F1 y SIxNEW F1 mostraron los valores más altos, seguidos por sus progenies SIxDC F2 y SIxNEW F2. La familia SIxSAI presentó una heredabilidad de 0,485 para la variable rendimiento por planta, seguida por la familia SIxDC con un valor de 0,462, mientras que la familia SIxNEW registró un valor de 0,076, el más bajo de todos.
Palabras clave: fenotipo, genotipo, heredabilidad, variables de rendimiento.
ABSTRACT
Increasing productivity through plant breeding relies centrally on estimating heritable factors associated to yield and precocity of plant materials. With that purpose, the present study evaluated 10 pea genotypes resulting from subsequent generations of four original parental lines: Santa Isabel (SI), San Isidro (SAI), Diacol Caldas (DC) and New Era Wisconsin (NEW). The study was carried out under greenhouse conditions in the Bogota Plateau, throughout one production cycle. Analyzed through average comparison, the evaluated traits were: days to flowering, first flower node, plant height at flowering, days to fructification, pod breadth and length, number of pods per plant, number of seeds per pod, plant yield and weight of 100 seeds. Each trait was assessed in terms of estimating the averages of the parents and of their respective F1 and F2 progenies, in order to calculate heritability by the Warner method. The four parental materials, together with hybrid SIxSAI F1, produced the lowest yields; whereas hybrids SIxDC F1 and SIxNEW F1 attained the highest counts, followed by their respective progenies: SIxDC F2 and SIxNEW F2. Regarding plant yielding, family SIxSAI reached a heritability value of 0.485, followed by family SIxDC with 0.462. Family SIxNEW registered the lowest value (0.076).
Key words: phenotype, genotype, heritability, yield variables.
Introducción
La arveja es una importante fuente de alimento debido a su aporte proteico estimado en un 25-26%, además de ser una fuente de vitaminas, aminoácidos, carbohidratos, lípidos y minerales como calcio, hierro, fósforo, potasio y sodio. Se estima que en 100 g de parte comestible, aporta un total de 82 calorías (Lobo et al., 1989).
Es una leguminosa herbácea anual que se adapta normalmente a los climas fríos. Se han señalado como centros de origen de la especie las regiones montañosas del suroeste de Asia, en especial Afganistán, India y Etiopía. Un centro secundario de diversidad está ubicado en el Mediterráneo (Lobo et al., 1989).
La planta de arveja desarrolla un tallo primario o eje central, y puede tener tallos secundarios que nacen del nudo cotiledonar o de nudos superiores. El número de ramas está determinado por las condiciones ambientales, en especial por el fotoperiodo, pero también parte de la variabilidad observada se debe a diferencias genéticas (Hatam y Amanullah, 2001; Ligarreto, 2002).
La diversidad en arveja es amplia, con un gran número de cultivares que se han mejorado por rendimiento y por calidad del producto fresco o como materia prima para la agroindustria. Colombia cuenta con variedades de arveja mejoradas bastante antiguas, dentro de las que se encuentran: ICA Bojacá, ICA Teusacá y Diacol Caldas, entregadas en las décadas de 1960 y 1970; ICA Tominé, ICA-Corpoica Sindamanoy y San Isidro, liberadas en la década de 1990; no obstante, la variedad regional Santa Isabel es la que ocupa la mayor parte del área sembrada en el país, ya que posee buenas características comerciales como grano y vaina grandes, y es una de las variedades más rendidoras en vaina verde. Presenta un hábito de crecimiento voluble, con flores de color blanco. El grano seco es de forma redonda, liso y de color crema quemado; el hilum es negro. La altura promedio de la planta es de 1,70 m y es susceptible a patógenos como Ascochyta spp., Colletotrichum pisi Pat. y Fusarium oxysporum f. sp. pisi (Buitrago, 2006).
De las variedades mejoradas, la que más se siembra en el país es la San Isidro, que presenta un hábito de crecimiento voluble, sus flores son de color blanco, el grano es de forma redonda, liso y verde, con un hilum de color blanco. Los días hasta la floración van de 58 a 61; los días a cosecha van de 112 a 118 para grano en verde y de 140 a 147 para grano en seco. El rango de adaptación es de 2.400-2.800 msnm; la altura promedio de la planta es de 1,11 a 2,00 m. Este material presenta resistencia en campo a Ascochyta ssp. y a Colletotrichum pisi Pat. (Buitrago y Duarte, 2006).
La variabilidad ambiental afecta la relación entre genotipo y fenotipo. Este grado de asociación es medido por la heredabilidad (h2), la cual puede definirse como la proporción de la variación fenotípica total observada, debida a causas genéticas. Cuanto mayor sea la heredabilidad, el fenotipo describirá más al genotipo. De esta forma, cuando se acerca al 100%, se garantiza que los fenotipos seleccionados son los de componente genético superior. Además, es necesario tener presente que las estimaciones de este parámetro son específicas para la población referencia sobre la que se ha medido y para las condiciones ambientales en las que se realiza la evaluación (Vallejo y Estrada, 2002).
La heredabilidad tiene una estrecha relación con la selección. La selección busca obtener y aislar grupos de individuos genéticamente mejores que la población original. El éxito de la selección varía de una población a otra, dentro de una especie o de especie a especie. Si hay considerable variación genotípica de tipo aditivo intralocus e interloci y una alta heredabilidad, la selección puede causar grandes y permanentes cambios en la población. La varianza genotípica de tipo aditivo puede estar presente, pero si la heredabilidad es baja, la selección no es efectiva para cambiar la población, a menos que esta se acompañe con apareamientos adecuados y un buen control ambiental (Vallejo y Estrada, 2002).
En arveja se han formulado modelos que permiten predecir el comportamiento de caracteres de rendimiento, calidad de grano y precocidad y su interacción con el ambiente; sin embargo estos modelos han sido realizados para ser utilizados en zonas donde existen estaciones (González, 2001). De igual manera, no se tiene información sobre el cruzamiento específico de arvejas volubles con adaptación a la zona andina y variedades foráneas. Por consiguiente, en este trabajo se pretende aportar en la estimación de la heredabilidad para parámetros fenológicos y de rendimiento a partir de progenitores contrastantes por hábito de crecimiento y por susceptibilidad a Fusarium oxysporum f. sp. pisi, como un aporte inicial para generar conocimiento útil al mejoramiento genético de la arveja.
Materiales y métodos
Se evaluaron 10 materiales de arveja constituidos por cuatro progenitores, tres generaciones F1 y tres F2; como parentales se utilizaron los cultivares volubles: Santa Isabel (SI), San Isidro (SAI) y Diacol Caldas (DC). Este último cultivar presenta resistencia a Fusarium oxysporum, f. sp. pisi en campo, es 15 d más precoz la floración que las otras dos variedades y su color de flor es morada. New Era Wisconsin (NEW), de porte bajo, precoz y con resistencia a las razas 1 y 2 de Fusarium oxysporum, f. sp. pisi (Kraft y Pfleger, 2000). Las progenies F1 y F2 resultaron de cruzar la variedad Santa Isabel con cada uno de los otros cultivares. Los cruces se realizaron a través de la técnica de hibridación artificial en arveja por emasculación (Ligarreto y Patiño, 2004).
Los 10 materiales fueron sembrados bajo cubierta protegida en los invernaderos de la Facultad de Agronomía de la Universidad Nacional de Colombia (Bogotá), a una altitud de 2.600 msnm y a temperatura promedio de 14°C, mínima promedio de 4°C y una máxima promedio de 20,5°C, humedad relativa de 80% y brillo solar de 4,5 h d-1. Los materiales fueron colocados en un diseño completamente al azar con tres repeticiones; para los parentales y los híbridos F1 se sembraron 10 semillas por unidad experimental y para las poblaciones F2 se sembraron 20 semillas por replicación, siempre distanciadas a 10 cm.
Se evaluaron los caracteres días hasta la floración, el número del nudo del tallo donde aparece la primera flor, la altura de la planta a floración, los días hasta la fructificación, el número de vainas por planta, el rendimiento por planta, el peso de 100 semillas, el ancho de la vaina, la longitud de la vaina y el número de semillas por vaina; las tres últimas variables fueron medidas sobre 10 vainas por planta. Con los datos generados a partir de la evaluación de los 10 tratamientos se determinó la heredabilidad en sentido amplio o genotípica, y es la que agrupa todos los factores genéticos que afectan la expresión de un determinado carácter; la varianza genética (σ2G) mide la variación debida a efectos genéticos, y es un componente necesario para calcular la heredabilidad de un determinado carácter al igual que la varianza fenotípica (σ2F), que es la variación fenotípica total observada, sea esta debida a efectos genéticos o ambientales (Poehlman y Sleper, 2003). El método utilizado fue el de Warner (Reyes, 1985), que presenta un estimador real para la varianza ambiental, al considerar esta como el promedio de las varianzas de las tres poblaciones P1, P2 y F1, que teóricamente no deben tener variación genotípica, el cálculo aplica la siguiente ecuación (1).
La comparación de promedios de parámetros fenotípicos se realizó por la prueba de Tukey, y para establecer el grado de asociación entre variables se utilizó la correlación de Pearson. Los análisis estadísticos de los datos se efectuaron con el programa SAS® versión 9 (SAS Institute Inc., Cary, NC).
Resultados y discusión
Para el componente de precocidad, variable días a floración, el material Diacol Caldas fue el más precoz con 48 d, mientras que la variedad Santa Isabel fue la más tardía con 64 d (Tab. 1). Las progenies F1 florecieron después que las progenies F2, lo que expresa el vigor de la progenie F1, haciéndola más tardía, excepto el híbrido SIxNEW F1 que floreció primero que su progenie F2. La ganancia en el proceso de selección es favorable en la precocidad a partir de los cruces de padres contrastantes, SI (tardío) y NEW (precoz). La importancia de este cruce puede ser la rápida fijación del carácter de precocidad en la progenie, posiblemente debido a la acción aditiva de los genes (Dhillon y Chanal, 1981; Bourion et al., 2002; Espinosa y Ligarreto, 2005).

Según Khvostova (1983), en arveja se pueden establecer cinco clases del tiempo a floración: las arvejas de día neutro (DN), las de iniciación temprana (EI), las tardías (L), las tardías con respuesta aumentada (LHR) y las muy tardías (VL), las cuales se explican por diferentes combinaciones de los genes: Lf, Sn, E y Hr (Larrain y Alcalde, 2003). Los alelos lfa, lf, Lf y Lfd confieren nudos de iniciación floral mínimos de 5, 8, 11 y 15, respectivamente. En este sentido, los cuatro parentales -incluido Diacol Caldas y sus progenies- pueden poseer el gen Lfd. De igual manera, dada la precocidad de Diacol Caldas, es posible que esté presente en su combinación genética el alelo E (dominante) o Hr que es epistático sobre E y adelanta la floración (Larrain y Alcalde, 2003).
La variedad Santa Isabel no presentó diferencias significativas en el número del nudo de aparición de la primera flor con ninguna de sus progenies F1 y F2, pero sí mostró diferencias con los parentales San Isidro, New Era Wisconsin y Diacol Caldas. El material Diacol Caldas presentó el valor medio más bajo con 13,21 nudos a la aparición de la primera flor, sin exhibir diferencia estadística con New Era Wisconsin. Diacol Caldas es voluble pero comienza a florecer de manera temprana incluso 11 d antes de New Era Wisconsin (Tab. 1).
Los cuatro parentales presentaron promedios más bajos en altura de la planta a floración que sus respectivas progenies (Tab. 1), lo cual se puede explicar por el valor heterótico de las progenies; se resalta el comportamiento de la progenie SIxNEW F2 y del híbrido SIxNEW F1, que presentaron las medias más altas, expresando la herencia trasgresiva hacia el progenitor SI de hábito indeterminado que es dominante sobre el hábito determinado de NEW. Cuando los progenitores no fueron contrastantes en el hábito de crecimiento, como SIxSAI y SIxDC, sus híbridos F1 mostraron vigor en la altura de la planta presentando valores medios más altos que sus respectivas progenies segregantes F2. Estos resultados están acordes con lo sugerido por Bourion et al. (2002) para quienes la expresión de la altura de la planta se atribuye principalmente a efectos genéticos sobre los cuales es eficiente un proceso de selección.
El comportamiento de la variable días hasta la fructificación para cada uno de los materiales, tanto parentales como híbridos F1 y progenies F2, coinciden con el orden presentado para días a floración; sin embargo se presentó una excepción con la variedad SI que floreció después que el híbrido SIxDC F1, pero en la variable días hasta la fructificación, el híbrido SIxDC F1 mostró una media mayor a la variedad SI, la cual fue de 66 d.
Para el componente de rendimiento, la variable número de vainas por planta (Tab. 2), la variedad Diacol Caldas y sus progenies presentaron los valores más altos, respectivamente. Se resalta el alto potencial de rendimiento que puede tener la variedad Diacol Caldas y sus progenies del cruce con Santa Isabel, originado por el mayor número de vainas por planta en compensación con los otros componentes de acción poligénica, como peso de la vaina y número de semillas por vaina (Timerman-Vaughan et al., 2004). San Isidro y sus progenies registraron los valores medios más bajos y no presentaron diferencias significativas entre ellas.
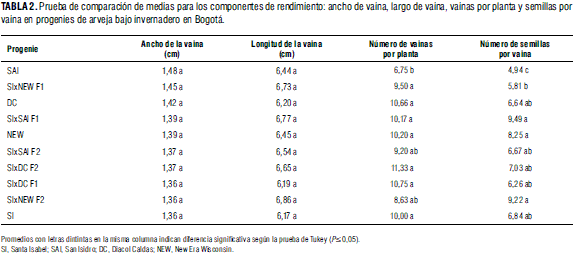
El número de semillas por vaina (Tab. 2) evidenció diferencias significativas entre materiales. Así el híbrido SIxSAI F1 y la progenie SIxNEW F2 presentaron, respectivamente, las medias más altas, consecuente con lo observado para la variable largo de vaina, donde los mismos materiales registraron los valores medios más altos, y sin efecto alguno por parte del ancho de la vaina, dada su alta uniformidad en tamaño de la vaina entre los progenitores, como también porque esta variable depende de muchos factores genéticos, como lo expresa Khvostova (1983). La variedad San Isidro con valor medio de semillas por vaina de 4,94 presentó diferencias significativas con todos los materiales. De igual forma, el híbrido SIxNEW F1 estuvo muy por debajo del valor registrado por los parentales New Era Wisconsin y Santa Isabel (Tab. 2). Por lo general, en esta variable se expresa el vigor híbrido de la F1 y la máxima expresión de la segregación en la F2 originando comportamiento superior de las filiales con respecto a los progenitores, situación que coincide con lo reportado por Gupta et al. (1986) y Espinosa y Ligarreto (2005).
El número de semillas por vaina es uno de los componentes de rendimiento más importantes en el mejoramiento de la arveja para el caso de Colombia, donde más de 95% de la producción de arveja se destina al mercado de vaina verde. Lo deseable es un número igual o superior a seis granos por vaina, pero la mejora genética es difícil dado que hay mecanismos de compensación de los componentes del rendimiento, y si se aumentan las semillas por vaina, se puede disminuir el número de vainas por planta y el peso de la semilla; una alternativa es desarrollar trabajos en mapeo de ligamiento de QTL para rendimiento y realizar su aplicación en la mejora genética de la especie (Timerman- Vaughan et al., 2004).
Timmerman-Vaughan et al. (2004) encontraron cinco QTL en donde los alelos marcadores asociados con los loci tienen efectos contrarios para peso de semilla y número de semillas, avanzando en la explicación genética de la compensación de los componentes de rendimiento. El número de semillas y el rendimiento de las semillas están altamente correlacionados; sin embargo los QTL tan solo coinciden en dos regiones genómicas.
Para el rendimiento por planta (Tab. 3) se registraron diferencias significativas entre materiales, en donde las F1 y F2 de los híbridos SIxDC y SIxNEW presentaron los valores más altos, respectivamente. Tanto en rendimiento por planta como en la variable peso de 100 semillas los progenitores registraron los valores medios más bajos (Tab. 3), al igual que en rendimiento la variedad New Era Wisconsin evidenció el menor peso de 100 granos, resultado esperado porque es un material precoz y tiene menor tiempo para el llenado del grano.

Se puede observar que una de las consecuencias de cruzar la variedad Santa Isabel con las variedades San Isidro, Diacol Caldas y New Era Wisconsin es la obtención de híbridos F1 con un alto valor agronómico en la descendencia, haciendo que estos tengan un promedio más alto que el de sus padres, en especial en las variables altura de la planta a floración, rendimiento por planta y el peso de 100 semillas. Algunas progenies F2 mostraron un comportamiento similar, regisregistrando un promedio superior no solo al de los parentales sino también al de sus F1. Entre los resultados que ilustran este caso se tiene que los dos valores más altos registrados en las variables altura de la planta a floración fueron para las progenies SIxNEW F1 y SIxNEW F2; en longitud de la vaina fueron SIxNEW F2 y SIxSAI F1; en vainas por planta fueron los cruzamientos SIxDC F2 y SIxDC F1; en semillas por vaina fueron SIxSAI F1 y SIxNEW F2; en rendimiento por planta fueron las progenies SIxDC F1 y SIxNEW F1, y en peso de 100 semillas alcanzaron el mayor valor los híbridos SIxNEW F2 y SIxSAI F1.
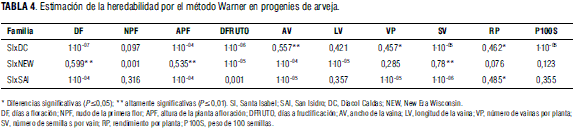
De otra parte, se tiene que los materiales evaluados de arveja tienen capacidad diferencial para heredar caracteres de rendimiento a sus progenies. Hay casos extremos (Tab. 4) como en la variable semillas por vaina, en donde la heredabilidad para la familia SIxNEW es alta, con un valor de 0,78, mientras que en la familia SIxSAI fue nula con valor de 1·10-06. Esta misma familia en la variable rendimiento por planta presentó la mayor heredabilidad, alcanzando el valor de 0,485, considerada relativamente alta por ser un carácter de naturaleza poligénica y estar influido por las condiciones ambientales (Dhillon y Chanal, 1981; Sarawat et al., 1994). También la familia SIxNEW presentó heredabilidades media altas para las variables días floración y altura de la planta a floración, con valores de 0,599 y 0,535, respectivamente. La generalidad en el estudio fue que la heredabilidad de los caracteres cambió drásticamente de una familia a otra.
La matriz de correlación entre los diferentes componentes de rendimiento (Tab. 5) deja ver una alta correlación entre las variables altura de planta a floración, días hasta la fructificación y vainas por planta; también la variable altura de planta a floración presenta una correlación alta con la variable rendimiento por planta, mientras, que la variable días hasta la fructificación exhibe una asociación alta con la variable vainas por planta. Todo esto es posible porque las variedades de hábito voluble indeterminadas predominantes en el estudio son tardías y presentan mayores rendimientos que las de hábito determinado como NEW.
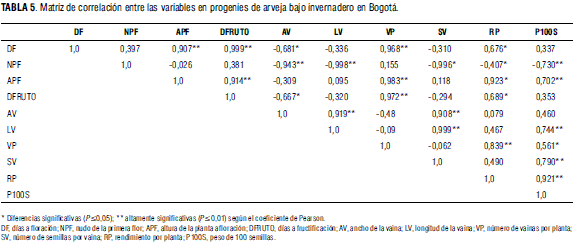
La variable vainas por planta presenta una alta correlación con la variable rendimiento por planta, la cual a su vez se correlaciona altamente con la variable peso de 100 semillas. Existe una alta correlación entre la altura de la planta a floración con el número de vainas por planta y el rendimiento por planta. Es lógico esperar este tipo de relaciones positivas entre estas variables, ya que una es dependiente de las otras para la compensación del rendimiento final, hecho que coincide con lo reportado por González y Ligarreto (2006). Lo anterior refuerza la teoría de que el rendimiento del grano en arveja tiene pocas posibilidades de mejorarse si se seleccionan componentes de rendimiento individuales (Sarawat et al., 1994).
Los resultados anteriores para precocidad y rendimiento permiten vislumbrar que para las familias en estudio la efectividad en la mejora genética puede aumentar de manera considerable cuando se selecciona por caracteres clave en generaciones tempranas, tal como lo sugieren Dhillon y Chanal (1981) y Sarawat et al. (1994). Cousin (1997) expresa que hay gran variabilidad genética para componentes de rendimiento, pero al hacer cruces para mejorar estos caracteres individualmente hay a menudo reducción del rendimiento.
Debido a los resultados obtenidos en este estudio, es necesario evaluar los materiales en diferentes condiciones ambientales para tener un mejor estimador de la heredabilidad, como también poder conocer la estabilidad de los mismos dado su comportamiento agronómico. Esto permitirá desarrollar nuevas variedades de arveja que expresen un alto potencial de rendimiento para las diferentes regiones agroecológicas del país.
Literatura citada
Bourion, V., G. Fouilloux, C. Le Signor e I. Lejeune-Hénaut. 2002. Genetic studies of selection criteria for productive and stable peas. Euphytica 127, 661-273. [ Links ]
Buitrago, J. 2006. Obtención de progenies de arveja (Pisum sativum L) por cruzamientos convergentes y su reacción a Fusarium oxysporum f. sp. pisi. Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. [ Links ]
Buitrago, J. y C. Duarte. 2006. El cultivo de la arveja en Colombia. Fenalce; Produmedios, Bogotá. [ Links ]
Cousin, R. 1997. Peas (Pisum sativum L.). Field Crops Res. 53, 111- 130. [ Links ]
Dhillon, G. y G. Chanal. 1981. An analysis of combining ability and reciprocal effects in garden pea (Pisum sativum L.). J. Res. Punjab Agric. Univ. 18(4), 359-364. [ Links ]
Espinosa, N. y G. Ligarreto. 2005. Evaluación de la habilidad combinatoria y heterosis de siete progenitores de arveja Pisum sativum L. Agron. Colom. 23(2), 197-206. [ Links ]
González, M.R. 2001. Interacción genotipo x ambiente en guisante proteaginoso (Pisum sativum L.). Tesis de doctorado. Universidad de Valladolid, España. [ Links ]
González, F. y G. Ligarreto. 2006. Rendimiento de ocho genotipos promisorios de arveja arbustiva (Pisum sativum L.) bajo sistemas de agricultura protegida. Fitotecnia Colombiana 6(2), 52-61. [ Links ]
Gupta, K., B. Dahiya y K. Singh. 1986. Combining ability studies over environments in pea. Crop Improv. 13(2), 134-137. [ Links ]
Hatam, M. y Amanullah. 2001. Grain yield potential of garden peas (Pisum sativum L.) germplasm. J. Biol. Sci. 1(4), 242-244. [ Links ]
Khvostova, V. 1983. Genetics and breeding of peas. USDA, Springfield, IL. pp. 12-96. [ Links ]
Kraft, J.M. y F.L. Pfleger. 2000. Compendium of pea diseases and pests. APS Press, St. Paul, MN. [ Links ]
Larrain, M. y J. Alcalde. 2003. Determinación de las fases de sensibilidad e insensibilidad al fotoperíodo en la etapa de prefloración en arveja (Pisum sativum L.). Cien. Inv. Agr. 30(1), 15-25. [ Links ]
Ligarreto, G. 2002. El cultivo de arveja: manejo agronómico. pp. 13-20. En: Memorias Primer Seminario Taller del Cultivo de la Arveja. Bayer CropScience, Bogotá. [ Links ]
Ligarreto, G. y P. Patiño. 2004. Hibridación artificial de arveja. Centro de Investigación y Extensión Rural (CIER). Boletín Divulgativo 1. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. [ Links ]
Lobo, M., C. Medina y M. Escobar. 1989. Manejo y conservación de los recursos genéticos de arveja (Pisum sativum). Investigación en los cultivos de arveja y haba. Boletín Técnico. IICA; Prociandino, Quito. [ Links ]
Poehlman, J.M. y D.A. Sleper. 2003. Mejoramiento genético de las cosechas. 2a ed. Ed. Limusa, México D.F. [ Links ]
Reyes, P. 1985. Fitogenotecnia básica y aplicada. ACT Editor, México DF. [ Links ]
Sarawat, P., F. Stoddar, D. Marshall y S. Ali. 1994. Heterosis for yield and related characters in pea. Euphytica 80, 39-48. [ Links ]
Timerman-Vaughan, G., A. Mills, T. Frew, R. Butler, J. Mccallum, S.Murray, C. Whitfield, A. Rusell y D. Wilson. 2004. Linkage mapping of QTLs for seed yield, yield components and developmental traits in pea (Pisum sativum L.). En: 4th Intl. Crop Sci. Congr. Brisbane, Australia. [ Links ]
Vallejo, F. y E. Estrada. 2002. Mejoramiento genético de plantas. Universidad Nacional de Colombia, Palmira, Colombia. [ Links ]

