










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.27 n.3 Bogotá set./dez. 2009
Leidy Rache C.1 y José Pacheco M.1, 2
1 Laboratorio Bioplasma, Escuela de Ciencias Biológicas, Universidad Pedagógica y Tecnológica de Colombia, Tunja.
2 Autor de correspondencia. jocpach@gmail.com
Fecha de recepción: 5 de febrero de 2009. Aceptado para publicación: 6 de noviembre de 2009
RESUMEN
Este estudio desarrolló un protocolo para micropropagación de Espeletiopsis muiska -especie de importancia ecológica, medicinal e industrial, actualmente en riesgo de extinción- en medio de cultivo Murashige y Skoog (1962). Se cultivaron embriones sexuales en medio completo y con la mitad de la concentración habitual de las macrosales; con y sin reguladores de crecimiento. Para proliferación de brotes se sembraron segmentos caulinares apicales en el medio de cultivo mencionado, con auxinas y citoquininas adicionadas individualmente o combinadas. Además se evalúo el efecto de diferentes concentraciones de caseína hidrolizada y sacarosa. Para enraizamiento se sembraron brotes en medio del cultivo con ácido 3-indol-butírico. La aclimatización de las plántulas -proceso que se inició en cuarto de incubación y finalizó en invernadero- se llevó a cabo en diferentes sustratos. El mayor porcentaje de embriones germinados (91%) se contabilizó en medio sin reguladores de crecimiento. En el medio de proliferación con ácido 3-indol-butírico 2,46 μM más 6-bencilaminopurina 0,044 μM se contabilizó la mayor cantidad de brotes de apariencia normal (5,96 brotes/explante). Durante la fase de enraizamiento el porcentaje de brotes enraizados en medio con ácido 3-indol-butírico 0,49 μM, fue del 54,1%. Después de 30 días de aclimatización, se constató la viabilidad del 60% de las plántulas transferidas a suelo de capote.
Palabras clave: citoquininas, auxinas, sacarosa, caseína hidrolizada, propagación.
ABSTRACT
The study developed a protocol for the micropropagation of Espeletiopsis muiska, an endangered species of ecological, medicinal and industrial importance. Sexual embryos were cultured in vitro in Murashige and Skoog (1962) complete and incomplete (reducing macrosalts by half) mediums, with and without growth regulators. Initial proliferation was carried out by culturing apical stem segments in medium with individual and combined addition of auxins and cytokinins; also evaluating the effect of different concentrations of hydrolyzed casein and sucrose. Rooting was promoted by growing the shoots in culture medium with 3-indole-butyric acid. Plantlet acclimatization, which started in incubation room and finished in greenhouse, made use of different substrates. The highest percentage of germinated embryos (91%) was obtained in growth regulator free medium. The largest number of normal appearance shoots per explant (5.96) was observed in proliferation medium containing 2.46 μM 3-indole-butyric acid plus 0.044 μM 6-benzylaminopurine. During the root taking phase, the medium containing 0.49 μM 3-indole-butyric acid promoted a rooting percentage of 54.1%. After 30 days of acclimatization, 60% of the plantlets transferred to soil enriched with organic matter resulted viable.
Key words: cytokinins, auxins, sucrose, hydrolyzed casein, propagation.
Introducción
Espeletiopsis muiska, frailejón nativo de los páramos colombianos, se localiza en subpáramos, 2.800 a 3.200 msnm y páramos, 3.200 a 3.800 msnm, con temperaturas de 6 a 12°C y precipitaciones de 500 a 1.000 mm año-1 (Cuatrecasas, 1976). Los frailejones son importantes industrial y medicinalmente (Correa y Bernal, 1990) y esenciales en ecosistemas de páramo debido a la gran biodiversidad que albergan, artrópodos entre otros (van der Hammen, 2002), y por contribuir a la regulación y al mantenimiento de fuentes hídricas mediante la captación de agua proveniente de procesos de condensación (van der Hammen, 2002); sin embargo, dado que las áreas de páramo están siendo intervenidas y utilizadas para el establecimiento de cultivos, la mayoría de los frailejones se encuentran en alto riesgo de extinción (Muñoz, 2002; Calderón, 2004).
Hace diez años la población de individuos de E. muiska era de 350 en la Reserva Natural de la Municipalidad de Ranchería que colinda con Paipa y Sotaquirá. Actualmente existen menos de 40, lo cual representa una disminución de 90% debida a las actividades de pastoreo y cultivos agrícolas establecidos (Posada, 2009). A pesar del riesgo de extinción, no se conocen estudios sobre la propagación de esta especie; algunos reportes abordan temas como: "Un ácido labdanoico componente mayoritario en Espeletiopsis muiska" (Ramírez et al., 2000); "Estudio químico de la Espeletiopsis muiska (Cuatr.) y evaluación de la actividad antimicrobiana" (Ramírez et al., 2000) y "Estudio preliminar del cariotipo y del ciclo celular en Espeletiopsis muiska (Cuatrec)" (Rondón y Pacheco, 2006).
El objetivo de esta investigación fue establecer un protocolo de micropropagación como una alternativa para producción masiva de plántulas de E. muiska utilizables para repoblamiento de áreas de páramo intervenidas.
Materiales y métodos
Condiciones generales de cultivo
El medio base utilizado fue MS (Murashige y Skoog, 1962) suplementado con (en mg L-1): ácido nicotínico 0,5; piridoxina 0,5; glicina 2,0; tiamina 0,1; sacarosa 30.000 y agar 6.500, pH de 5,8 ajustado con KOH y/o HCl (0,5-1,0 N). Los medios se autoclavaron durante 20 min, a 1.500 kPa y 121°C. Los cultivos se mantuvieron en cuarto de incubación a 24±1°C, con iluminación continua (70-80 μmol m² s-1) suministrada por lámparas fluorescentes de 75 W.
Recolección y caracterización del material vegetal de partida
En el Parque Natural La Ranchería (Paipa, Boyacá, Colombia), ubicado a 3.200 msnm, temperatura promedio de 11°C y humedad relativa de 67%, se localizaron cinco plantas adultas de Espeletiopsis muiska; de cada planta se tomaron ocho a diez aquenios maduros (Fig. 1A) y se transportaron en bolsas plásticas; posteriormente se realizó el conteo y la descripción de semillas.

Establecimiento in vitro de cultivos
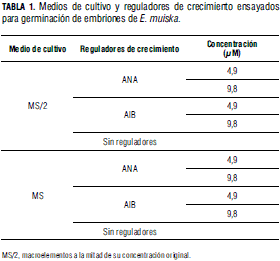
Para la asepsia de semillas, en cámara de flujo laminar, las semillas se enjuagaron con agua destilada estéril más Tween 20 (0,1% v/v) durante 5 min y luego se sumergieron consecutivamente en etanol (70%) durante 5 min y en NaOCl (5,25% p/v) al 2% (v/v) durante 10 min. Finalmente, se enjuagaron cuatro veces consecutivas (2 min cada una) con agua destilada estéril. Con ayuda de pinzas y bisturí, las semillas se escarificaron (Fig. 1B) y los embriones obtenidos se cultivaron durante 30 d en los medios indicados en la Tab. 1. Una vez determinado el medio de cultivo y el regulador de crecimiento más adecuado para germinación de embriones, se cultivaron en MS con diferentes concentraciones de AIB (0; 2,46; 4,9; 9,8; 14,7 y 24,6 μM). En cada tratamiento se cultivaron, en frascos de vidrio de 10 mL con alícuotas de 5 mL de medio, 30 embriones durante 30 d.
Proliferación de brotes axilares
Se utilizaron segmentos apicales de 1,0-1,5 cm de longitud (Fig. 1D) tomados de las plántulas obtenidas en la etapa anterior (Fig. 1C) y se cultivaron en recipientes de vidrio de 100 mL con alícuotas de 20 mL de medio. La proliferación de brotes comprendió dos fases: inducción y expresión (30 d cada una); la fase de expresión se realizó en medio sin reguladores. Se realizaron tres ensayos de proliferación cultivando 30 segmentos apicales por tratamiento y repitiendo cada ensayo dos veces. Los datos de cada ensayo se tomaron al finalizar la fase de expresión, cuantificándose número y longitud de brotes axilares desarrollados/ explante y, longitud y número de hojas del tallo principal (explante primario desarrollado). La longitud de brotes y tallo principal se midió sobre papel milimetrado estéril.
Ensayo 1
Efecto de auxinas y citoquininas. Para la fase de inducción, los segmentos caulinares se cultivaron en presencia de los reguladores indicados en la Tab. 2.

Ensayo 2
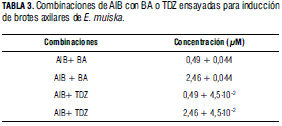
Efecto del AIB combinado con BA o TDZ. Con base en los resultados obtenidos en el ensayo anterior, durante la fase de inducción los segmentos caulinares se cultivaron en presencia de las combinaciones de reguladores indicadas en la Tab. 3.
Ensayo 3
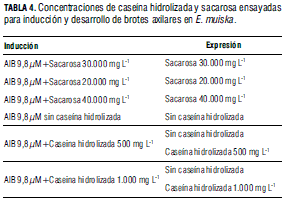
Efecto de diferentes concentraciones de caseína hidrolizada y de sacarosa. Los segmentos caulinares se cultivaron en medio suplementado con AIB, caseína hidrolizada o sacarosa en las concentraciones indicadas en la Tab. 4.
Enraizamiento de brotes
Brotes de longitud >2,0 cm, obtenidos en la etapa de proliferación (Fig. 1E), fueron individualizados y cultivados en medio con AIB (0; 4,9; 0,49 y 0,049 μM). Se cultivaron 30 brotes por tratamiento y cada tratamiento se realizó por duplicado. Después de 30 d de cultivo se cuantificó el número de brotes enraizados y el número de raíces/brote.
Aclimatización de plántulas
Una vez extraídas las plántulas de los frascos, las raíces se enjuagaron con agua corriente para eliminarles el agar. Posteriormente, las plántulas se transfirieron a vasos plásticos de 100 mL con los sustratos arena: tierra: cascarilla de arroz: tierra de capote (mantillo de bosque) en una proporsión 1:1:1:2; arena: tierra: cascarilla de arroz: tierra de capote (1:2:1:3) y tierra de capote.
Los vasos se colocaron en recipientes de vidrio de 50 x 30 x 10 cm cerrados con plástico extensible y se mantuvieron en cuarto de incubación (23°C y 97% de humedad relativa) durante 30 d, con riego por aspersión cada 8 d. Posteriormente, se trasladaron a invernadero y, de forma progresiva, durante 15 d, se eliminó el plástico extensible.
La aclimatización concluyó a 75-85% de humedad relativa, temperatura media diurna 18°C y nocturna 12°C. En invernadero las plántulas se regaron por nebulización durante 6 s cada 24 h. En cada uno de los sustratos utilizados se sembraron 20 plántulas, el ensayo se realizó dos veces y se cuantificó el porcentaje de plántulas viables.
Análisis estadístico
Los datos de número y longitud de brotes axilares desarrollados y longitud y número de hojas del tallo principal fueron analizados mediante Anava, con un nivel de confiabilidad de 95%. Para los factores que resultaron estadísticamente significativos se realizó la prueba LSD. Los datos fueron procesados con el paquete estadístico StatGraphic versión 2.0.
Resultados y discusión
Caracterización del material vegetal de partida
Las semillas presentaron forma espatulada, testa delgada y 4 a 5 mm de longitud. El embrión está recubierto por dos envolturas y su eje embrionario está entre los dos cotiledones. Como resultado de la observación de 500 semillas se encontró que 56% tenía embrión, 35% no tenía embrión y 9% estaba infestado con larvas de himenópteros.
Establecimiento in vitro de cultivos
La asepsia de semillas con NaOCl durante 10 min fue efectiva para eliminar los contaminantes superficiales y permitió la germinación de los embriones. Después de 30 d los embriones cultivados en MS y MS/2 suplementados con ANA, así como los embriones cultivados en MS/2 sin reguladores de crecimiento no germinaron, mientras que 85% de los embriones cultivados en MS sin reguladores de crecimiento y 74% de los cultivados con AIB germinaron y desarrollaron el eje embrionario. Con respecto a germinación in vitro de embriones, George (1996) indica que muchos compuestos de los medios de cultivo, así como los suplementos hormonales, pueden afectar de varias maneras los procesos morfogenéticos de crecimiento y desarrollo. Además, cuando los explantes son cultivados en medios con bajas concentraciones de sales minerales, se pueden producir alteraciones metabólicas que, a su vez, pueden retrasar e incluso interrumpir los procesos de crecimiento y desarrollo. Teniendo en cuenta el desarrollo de los embriones cultivados en MS, se evidenció que el medio proporcionó los requerimientos necesarios para el desarrollo progresivo normal de los embriones hasta la formación de plántulas; cabe señalar que la alta concentración de sacarosa que contiene el MS actúa no solo como fuente de energía sino que mantiene la osmolaridad del medio, factor de gran importancia en el cultivo de embriones (Montoya, 1991).
Después de 30 d de cultivo en MS suplementado con AIB (2,46-24,6 μM) se cuantificó 72-83% de embriones germinados; sin embargo, el mayor porcentaje de germinación, 91%, se cuantificó en MS sin reguladores de crecimiento. Proliferación de brotes axilares
Ensayo 1
En presencia de auxinas y citoquininas. Comparando la producción de brotes/por explante en medio sin reguladores de crecimiento (testigo) con la producción en medios suplementados con auxinas (Fig. 1 H, I, J), la mayor cantidad (5,7) se cuantificó (Tab. 5) en presencia de 14,7 μM AIB, mientras que en presencia de ANA se cuantificó la producción más baja. Los datos sobre longitud de brotes axilares desarrollados presentaron diferencias significativas, observándose que en presencia de 14,7 μM de AIB los brotes fueron de mayor longitud promedio (2,13 cm), mientras que en presencia de ANA (9,8 y 14,7 μM) los brotes fueron de menor longitud con respecto a los producidos y desarrollados en presencia de AIB, AIA y en MS sin reguladores. Meyer y Staden (1991) también estudiaron el efecto del AIB en Aloe barbadensis y obtuvieron resultados similares a los de este ensayo, mayor cantidad de yemas axilares en desarrollo en medio con AIB, seguido del medio suplementado con AIA y, en menor cantidad, en medio con ANA.
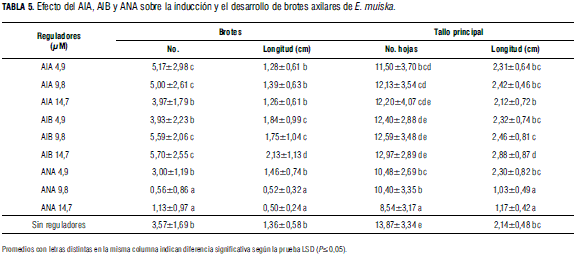
Durante la fase de proliferación todos los explantes cultivados desarrollaron un tallo principal, observándose que la mayor longitud promedio (2,88 cm) la presentaron los explantes cultivados en presencia de 14,7 μM de AIB. Con respecto al efecto de las auxinas sobre el número de hojas desarrolladas, la mayor cantidad de hojas/explante se cuantificó en los brotes cultivados en MS sin auxinas. Además, la longitud de brotes, número de hojas y longitud del tallo principal fueron semejantes en presencia de las diferentes concentraciones de AIA, mientras que el aumento de la concentración del AIB mostró un efecto positivo y progresivo sobre dichos parámetros. Estos resultados pueden ser explicados en razón de que las auxinas no solo tienen capacidad para inducir agrandamiento y alargamiento celular, sino que, además, se ha encontrado que promueven división celular en tejidos cultivados in vitro (Krikorian, 1991).
En este ensayo de proliferación se observó que de las auxinas ensayadas, ANA fue la menos efectiva, cuantificándose una disminución progresiva y significativa tanto en el número y la longitud de brotes axilares como en el número de hojas y la longitud del tallo principal a medida que aumentó la concentración (Tab. 5). Andrade et al. (1999) en Lavandula vera, observaron una reducción significativa en el número de brotes desarrollados por explante al suplementar con ANA el medio de proliferación.
Los datos de proliferación de brotes en presencia de citoquininas (Tab. 6, Fig. 1 K, L, M) indicaron que la mayor producción de brotes en desarrollo/explante, se produjo con 0,44 μM de BA; en contraste, el más bajo se cuantificó en el medio sin reguladores (Tab. 6).
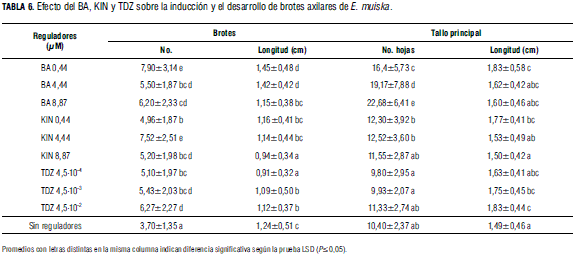
Con respecto a longitud de brotes axilares desarrollados, los resultados de los diferentes tratamientos presentaron diferencias estadísticamente significativas, cuantificándose la más alta, en los explantes cultivados con 0,44 μM de BA. La mayor cantidad de hojas por tallo, se encontró en presencia de 8,87 μM de BA, mientras que la menor cantidad se cuantificó en presencia de 4,5.10-4 μM de TDZ. El tallo principal presentó mayor longitud, 1,83 cm, en presencia de 0,44 μM de BA y 4,5.10-2 μM de TDZ (Tab. 6).
Los datos registrados en este ensayo indican que en los medios de cultivo con 0,44 μM de BA se obtuvieron los mayores promedios de brotes axilares, longitud de brotes y número de hojas; Hossain et al., 2008, obtuvieron 30 a 50 tallos por explante de Stevia en medio con BA. En los medios con TDZ la apariencia de los brotes fue anormal, presentando tallos y hojas notoriamente alargadas. También se evidenció que el BA fue la citoquinina más efectiva para inducir desarrollo de brotes axilares. Observando, en orden secuencial, la efectividad de las citoquininas sobre la formación de brotes, los resultados concuerdan en parte con el orden de eficacia citado por Sujatha y Ranjitha (2007) en Artemisia vulgaris: BA>Kn>Zeatina>Adenina. Además, se observó un efecto negativo del TDZ sobre el desarrollo de brotes de E. muiska.
Comparando los resultados obtenidos en los diferentes tratamientos de este ensayo, se observó que en los cultivos de proliferación de brotes en MS con auxinas o citoquininas (Tab. 5 y 6), mientras el AIB favoreció más la elongación, el BA estimuló más la proliferación de brotes. Además, en presencia de BA se cuantificó la mayor cantidad de hojas/tallo principal, 22,68, mientras que en el ensayo con auxinas, el mayor número se cuantificó en el testigo (medio sin reguladores de crecimiento) 13,87. Con base en estas comparaciones se infirió que una apropiada combinación de auxina con citoquinina favorecería una adecuada elongación y un aumento del número de brotes desarrollados. Respecto a esta inferencia, Sotolongo et al. (2003) sugieren la utilización de auxinas con el fin de superar y reducir los efectos residuales de las citoquininas favoreciendo la elongación de los brotes.
Ensayo 2
Efecto del AIB combinado con BA o TDZ (Fig. 1 N, O). Los datos registrados para número de hojas, longitud del tallo principal y longitud de brotes desarrollados (Tab. 7) no mostraron diferencias estadísticas significativas entre los tratamientos realizados. Mientras que para número de brotes/explante se observaron diferencias significativas, cuantificándose el mayor número de brotes/explante (6,93) en presencia de 2,46 μM de AIB y 4,5.10-2 μM de TDZ, pero estos brotes presentaron hojas anormalmente alargadas.
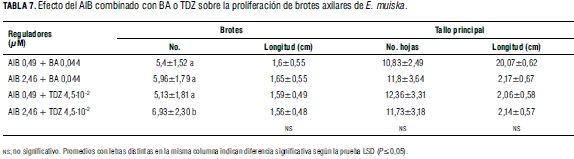
Aunque en presencia de 2,46 μM de AIB y 0,044 μM de BA se cuantificó 5,96 brotes/explante, los brotes presentaron apariencia normal y adecuada para ser transferidos a medio de enraizamiento y posterior aclimatización. Estos resultados corroboran el efecto positivo de la combinación auxina-citoquinina sobre la multiplicación celular indicado por Suárez et al. (2006) y la importancia de establecer balances adecuados auxina/citoquinina para elevar las tasas de proliferación (Brucato et al., 2006).
Ensayo 3
Efecto de la sacarosa y la caseína hidrolizada. En los tratamientos con sacarosa y caseína hidrolizada, la longitud y el número de hojas del tallo principal no presentaron diferencias estadísticas significativas. Sin embargo, la longitud de brotes desarrollados en los tratamientos con sacarosa o caseína hidrolizada mostraron diferencias estadísticamente significativas, cuantificándose los valores medios más elevados, 1,83 y 2,13 cm de longitud de brotes en los segmentos caulinares cultivados en MS suplementado con 30.000 mg L-1 de sacarosa o en MS sin caseína hidrolizada. En general, el promedio de brotes por explante en los diferentes tratamientos de este ensayo fue alto, 5,26 a 7,22 (Tab. 8); sin embargo, al aumentar o disminuir la concentración de sacarosa o al utilizar caseína hidrolizada en los medios de proliferación, los brotes desarrollados presentaron apariencia anormal, con tallos delgados muy ramificados y con hojas deformes muy alargadas (Fig. 1P, Q). Cabe recordar que los carbohidratos sirven como fuente de carbono o energía para el metabolismo de las plantas cultivadas in vitro y actúan como agentes que modifican las condiciones osmóticas (Gemas y Bessa, 2006). Además, aunque la caseína hidrolizada ha sido utilizada para aumentar el crecimiento y la proliferación, en este ensayo se observó un efecto negativo sobre la proliferación de brotes. Además de los resultados descritos anteriormente, en los cultivos realizados en los diferentes ensayos de proliferación también se observó:
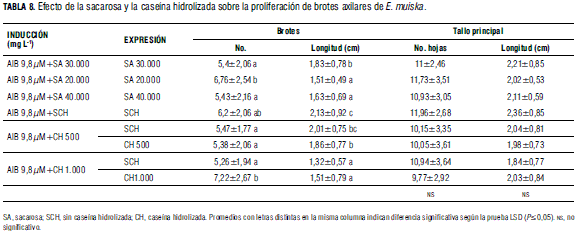
-Un mayor porcentaje de explantes necróticos se cuantificó a medida que aumentó la concentración de AIB y ANA, 2,4 a 16,9 y 4,8 a 24,05, respectivamente (datos no mostrados). Los menores porcentajes de explantes necróticos 7,0 y 8,3; 2,4, 2,5 y 4,1, se cuantificaron cuando los explantes se estimularon con 0,44 y 4,44 μMdeKIN, con sacarosa a diferentes concentraciones y cuando los explantes fueron estimulados en presencia de caseína hidrolizada. En los demás tratamientos no se presentó necrosis de tejidos.
-Al estimular segmentos caulinares apicales con AIA se cuantificaron bajos porcentajes de explantes con callo basal, 4,9 a 9,8% y de brotes hiperhidratados 3,35 a 10,00%. Al estimular con AIB, el porcentaje de explantes con callo basal aumentó, 30,9 a 71,5%, y se cuantificaron, en general, porcentajes similares de explantes hiperhidratados, 13,8; a 16,9, mientras que al estimular con ANA, 92,8 a 100% de los explantes formaron callo compacto y de color marrón (Fig. 2).
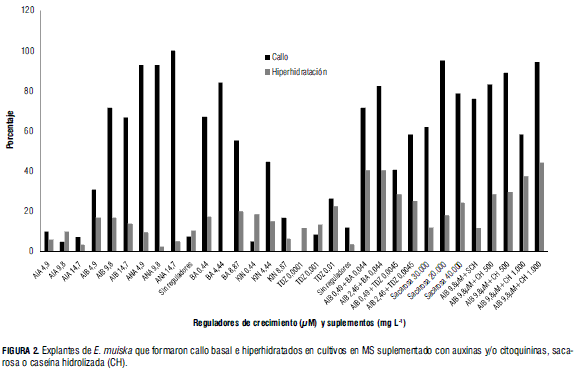
Todos los explantes que formaron callo posteriormente murieron. Al igual que en este ensayo, el ANA estimuló el crecimiento de callo en Stevia rebaudiana (Bondarev et al., 1997).
- En general, más de 50% de los explantes formó callo basal en presencia de las combinaciones AIB-BA o AIB-TDZ, sacarosa a diferentes concentraciones y caseína hidrolizada, excepto el tratamiento 23 (AIB 0,49 μM más TDZ 0,0045 μM) en el cual 40,8% de los explantes desarrollaron callo. El número de explantes hiperhidratados fue mayor cuando se cultivaron en MS con AIB-BA que cuando se cultivaron con AIB-TDZ. Al aumentar o disminuir la concentración de sacarosa y al incrementar la de caseína hidrolizada aumentaron los porcentajes de explantes hiperhidratados (Fig. 2).
Enraizamiento
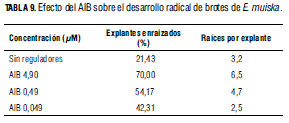
Después de 30 d de cultivo en medio de enraizamiento, se observó que a medida que se incrementó la concentración de AIB, también aumentó el porcentaje de brotes enraizados y la media de raíces por explante, mientras que en estudios realizados por Suárez et al. (2006) en Stevia rebaudiana, a medida que se aumentó la concentración de AIB en el medio de cultivo la formación de raíces disminuyó; sin embargo, en otros estudios con Stevia (Debnath, 2008 y Hossain et al., 2008), el mayor número, 2 a 8, de raíces/ explante se registró en presencia de 1,0; 1,5 y 2,0 mg L-1 de AIB. En este ensayo, aunque en presencia de 4,9 μM de AIB se cuantificó el mayor porcentaje, 70%, de brotes enraizados y el mayor promedio, 6,5, de raíces por explante (Tab. 9), el AIB también indujo formación y desarrollo de brotes en la base del explante, dificultando la aclimatización de estas plántulas. De otro lado, aunque en el medio con 0,49 μM de AIB se cuantificó 54,17% brotes enraizados y 4,7 raíces/ explante (Tab. 9, Fig. 1F), la inducción y el desarrollo de brotes axilares en la base del explante disminuyó, pero el porcentaje de plántulas viables en la fase de aclimatización aumentó.
Aclimatización
Después de 90 d de aclimatización, el mayor porcentaje, 60%, de plántulas viables se cuantificó en el sustrato tierra de capote (Fig. 1G), observándose que la textura y estructura de este sustrato favorecen el desarrollo caulinar y radical de las plántulas. Teniendo en cuenta que la tierra de capote presenta alta porosidad, elevada retención de agua, buena aireación, pH bajo y textura fina (Valencia y Ramírez, 1993), se convierte en un sustrato apropiado para la aclimatización de E. muiska. El control inicial de temperatura, humedad e iluminación fue imprescindible durante la primera etapa de aclimatización, observándose que las plántulas transferidas directamente del cuarto de cultivo a condiciones de invernadero no fueron viables (resultados no mostrados).
Conclusiones
A través de los ensayos realizados se estableció un protocolo de micropropagación de Espeletiopsis muiska que permite la producción masiva de plántulas utilizables para repoblamiento de áreas de páramo intervenidas.
El protocolo de micropropagación de E. muiska más adecuado comprende: germinación de embriones en MS sin reguladores de crecimiento; proliferación de brotes axilares en medio con 2,46 μM de AIB más 0,044 μM de BA; enraizamiento de brotes en medio con 0,49 μM de AIB y aclimatización de plántulas en condiciones controladas utilizando como sustrato suelo de capote.
Agradecimientos
Los autores agradecen al Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología "Francisco José de Caldas" (Colciencias) y a la Dirección de Investigaciones de la Universidad Pedagógica y Tecnológica de Colombia (UPTC), Tunja, el apoyo económico para el desarrollo del trabajo, así como a los integrantes del Grupo de Investigación Bioplasma, UPTC, por su valiosa colaboración.
Literatura citada
Andrade, L.B., S. Echeverrygaray, F. Fracaro, G.F. Pauletti y L. Rota. 1999. The effect of growth regulators on shoot propagation and rooting of common lavender (Lavandula vera DC). Plant Cell Tiss. Org. Cult. 56(2), 79-83. [ Links ]
Barcelló, J., G. Nicolás, B. Sabater y R. Sánchez. 1993. Fisiología vegetal. 6a ed. Ediciones Pirámide, Madrid. [ Links ]
Brucato, G., M. Graziella, I.E. Trujillo D. y M. Oropeza C. 2006. Micropropagación de Lepidium virginicum L. a partir de microesquejes. Agron. Trop. 56(4), 651-656. [ Links ]
Calderón S., E. 2004. Lista preliminar de plantas fanerógamas: familia Asteraceae (=Compositae). En: Instituto Humboldt, http://www.humboldt.org.co/conservacion/asteraceae.html; consulta: octubre de 2009. [ Links ]
Correa, J. y H. Bernal. 1990. Especies vegetales promisorias de los países del convenio Andrés Bello. Tomos XII. Secretaría Ejecutiva del Convenio Andrés Bello (SECAB); Ministerio de Educación y Ciencia de España; Corporación Andina de Fomento (CAF), Bogotá. [ Links ]
Cuatrecasas, J. 1976. A new subtribe in the Heliantheae (Compositae): Espeletiinae. Phytologia 35(1), 43-61. [ Links ]
Debnath, M. 2008. Clonal propagation and antimicrobial activity of an endemic medicinal plant Stevia rebaudiana. J. Med. Plant Res. 2, 45-51. [ Links ]
Gemas, V. y A. Bessa. 2006. Influence of various carbohydrates in shoot development in nodal culture of Guinean Anacardium occidentale genotypes. Plant Cell Tiss. Org. Cult. 85(1), 103-108. [ Links ]
George, E.F. 1996. Plant propagation by tissue culture. Parte 2 Practice. Exegetics, Edington, UK. [ Links ]
Hossain, M.A., A.H.M. Shamim-Kabir, T.A. Jahan y M.N. Hasan. 2008. Micropropagation of Stevia. Int. J. Sustain. Crop Prod. 3(4), 1-9. [ Links ]
Krikorian, A.D. 1991. Capítulo 3: Medios de cultivo: generalidades, composición y preparación. pp. 41-77. En: Roca, W.M. y L.A. Mroginski (eds.). Cultivo de tejidos en la agricultura. Fundamentos y aplicaciones. Editorial Centro Internacional de Agricultura Tropical (CIAT), Cali, Colombia. [ Links ]
Meyer, H.J. y L. van Staden. 1991. Rapid in vitro propagation of Aloe barbadensis Mill. Plant Cell Tiss. Org. Cult. 26, 167-171. [ Links ]
Montoya H., L.M. 1991. Cultivo de tejidos vegetales. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Medellín, Colombia. [ Links ]
Muñoz B., F. 2002. Propagación de flora endémica de páramo o en peligro de extinción en el Parque Nacional Natural El Cocuy. pp. 842-847. Memorias Congreso Mundial de Páramos. Tomo I. Paipa, Colombia. [ Links ]
Murashige, T. y F. Skoog. 1962. A revised medium for rapid growth and bio-assays with Tobacco tissue cultures. Physiol. Plant 15, 473-497. [ Links ]
Posada, J. 2009. Protección urgente para los páramos en Boyacá. En: Generación invisible, http://www.generacioninvisible.com/ index.php?option=com_content&task=view&id=135&Item id=2; consulta: octubre de 2009. [ Links ]
Ramírez, M.B., A.N. Téllez y R.D. Torrenegra. 2000. Estudio químico de la Espeletiopsis muiska Cuat (Cuatr) y evaluación de su actividad antimicrobiana. Rev. Latinoam. Quim. 28(3), 119-124. [ Links ]
Rondón B., H.I. y J.C. Pacheco M. 2006. Estudio preliminar del cariotipo y del ciclo celular en Espeletiopsis muiska (Cuatrec). Trabajo de grado. Facultad de Ciencias Básicas, Universidad Pedagógica y Tecnológica de Colombia, Tunja. [ Links ]
Sotolongo S., R., M. García L., L. Junco C., G. Geada L. y E. García. 2003. Micropropagación de Psidium salutare (Myrtaceae). Rev. Jard. Bot. Nac. 24(1-2), 245-250. [ Links ]
Suárez, I., M. Espitia e I. Pertuz. 2006. Efecto de auxinas y citocininas en la multiplicación y enraizamiento in vitro de Stevia rebaudiana Bertoni. Fitotecnia Colombiana 6(2), 1-8. [ Links ]
Sujatha, G. y B.D. Ranjitha K. 2007. Effect of phytohormones on micropropagation of Artemisia vulgaris L. Acta Physiol. Plant. 29(3), 189-195. [ Links ]
Valencia, M.L.C. de y F. Ramírez. 1993. Notas sobre la morfología, anatomía y germinación del agraz (Vaccinium meridionale Sw). Agron. Colomb. 10(2), 151-159. [ Links ]
Van der Hammen, T. 2002. Diagnóstico, cambio global y conservación. pp. 60-71. Memorias Congreso Mundial de Páramos. Tomo I. Paipa, Colombia. [ Links ]

