










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.27 n.3 Bogotá set./dez. 2009
FISIOLOGÍA DE CULTIVOS
Bernardo Chaves C.1, 3, Gerardo Cayón S.1 and James W. Jones2
1 Departamento de Agronomia, Facultad de Agronomia, Universidad Nacional de Colombia, Bogota.
2 Agricultural & Biological Engineering Department, University of Florida, Gainesville, FL.
3 Corresponding author. bchavesc@unal.edu.co
Received por publication: 16 April, 2009. Accepted for publication: 6 November, 2009.
ABSTRACT
Plantain is a basic food for more than 400 million people of the tropical and subtropical regions of the world. Crop modeling has become a useful agricultural tool whose outputs not only facilitate the scientific study of crop physiological processes, but also the adaptation of farmers' crop management decisions. By using physiological and climatic data from two experiments on variety Dominico-Harton (Musa AAB Simmonds), a plantain potential production model was developed. Based on the results, Light Extinction Coefficient (k) and Light Use Efficiency (LUE) were respectively estimated as 0.2817 and 1.63 g MJ-1. Likewise, aerial dry matter results allowed estimating partition coefficients for both the vegetative and the reproductive stage. Leaf, stem and corm dry matter were observed to increase in equal proportions during the vegetative stage. During the reproductive stage, only the stem was observed to increase its dry matter content (although not as much as in the vegetative stage), while leaves and corm were found to decrease it. A sensitivity analysis established LUE as the most sensitive parameter. In consequence, research efforts should be aimed at improving this conversion of radiant energy into dry matter.
Key words: growth, dry matter, light extinction coefficient, light use efficiency, photosynthetic active radiation.
RESUMEN
El plátano es un producto básico en la alimentación para más de 400 millones de habitantes de las regiones tropicales y subtropicales del mundo. Los modelos del desarrollo de cultivos se han convertido en una herramienta de mucha utilidad para investigadores que estudian procesos fisiológicos básicos, y agricultores que los usan en la toma de decisiones para el manejo del cultivo. Con el objetivo de desarrollar un modelo de producción potencial de plátano, se usaron los datos de dos experimentos con la variedad Dominico-Hartón en Colombia, en los cuales se midieron variables fisiológicas de crecimiento y desarrollo, así como variables climáticas. Con base en los resultados se estimó el coeficiente de extinción de la luz (k) en 0,2817 y el uso eficiente de la luz (LUE) en 1,63 g MJ-1. En la etapa vegetativa, las hojas, el tallo y el cormo incrementaron su materia seca por partes iguales, pero en la etapa reproductiva las hojas y el cormo perdieron masa, mientras que el tallo continuó en aumento, aunque no tan aceleradamente como en la primera etapa. Del análisis de sensibilidad se deduce que LUE es el parámetro más sensible y, por lo tanto, los esfuerzos se deben dirigir a mejorar la conversión de luz incidente en materia seca.
Palabras clave: crecimiento, materia seca, coeficiente de extinción de la luz, eficiencia en el uso de la luz, radiación fotosintéticamente activa.
Introduction
Plantain is cultivated worldwide in approximately 2.3 million ha, which produce about 18.3 million metric tons a year. Employing about 20 million people, Latin America and the Caribbean (LAC) produce 40% of the world's total production (7.3 t year-1), which is therefore critical to food security. Production is mostly destined to local consumption, and only 1% is exported to USA and Europe (Inibap, 2001). Colombia is the biggest plantain producer in LAC. Out of the 2.97 million tons that are produced in 400,000 ha, only 120,000 are exported. Colombia is also the first plantain consumer in LAC, averaging 135 kg per person and year (Belalcázar, 1991).
Plantains AAB are the largest subgroup of bananas, including numerous cultivars that vary in shape, size, color and taste. Plantain is cultivated in a traditional mode, often in combination with other crops (coffee, cocoa and others), thus attaining relatively low yields (10 average t ha-1). A popular feature of this crop is that it can be produced with little management care. Plantain is not involved in substantial international trade (less than 0.4% of total world agricultural exports), but is abundant in local markets. Production is mainly concentrated in Latin America (where Colombia is the first producer), the West Indies (The Dominican Republic, Haiti), and Central and West Africa (The Democratic Republic of Congo, Nigeria, Ghana, Côte d'Ivoire and Cameroon) where there is considerable diversity (Gowen 1995).
Plantain crop systems involve the complex interaction of several processes, out of which a useful model must deal with those that are relevant to its output. In order to model and simulate maximum possible dry matter production or potential yield, the effect of climatic factors such as temperature and radiation on development and growth processes must be considered. Climatic data availability allows estimating potential dry matter production for different locations.
A potential production model does not take into account water availability, soil nutrient shortage or pest and disease management (Gary et al., 1998). Thus, its dry matter calculations are always larger than field measurements. For this reason, it is difficult to validate potential production models. However, yield can be optimized by identifying those factors that limit it (Dourado-Neto et al., 1998; Meira and Guevara, 2000; Ranganathan et al., 2001). The model presented here was based on a simple and general crop growth model developed in Wageningen by Spitters and Shapendonk (1990) and Kooman (1995), involving light interception, daily conversion of light into dry matter production, and dry matter allocation to different plant organs.
A plantain potential production model not only can help growers determine the best environmental conditions to plant the crop and obtain better yields, but also facilitates taking crop management decisions (Lentz, 1998; Marcelis et al., 1998). The productive behavior of any plantain variety in a specific location depends on its genotype and on the environment where it grows, two key aspects that determine the diversity of agro climatic conditions that are usually found (Thornley and Jhonson, 1990; Uthaiah et al., 1992; Keen and Spain, 1992; Peart and Curry, 1998). Despite plantain's great adaptability to diverse environmental conditions, its productivity is still conditioned by the physiological limits imposed by such adaptation (Turner, 1994). Plentiful knowledge on plant physiological processes and on the way they are affected by environmental factors is necessary to attain good productivity in economically important species. Modeling crop growth as a function of environmental conditions allows building up efficient and sustainable production techniques, as well as deciding favorable genotypes adapted to different production zones.
Although the modeled crop can be a newly established or a previously existing plantation, the present work is specifically focused on the first possibility, and it is actually the first one in its kind. In this context, the objective of the present work was to develop a dynamic model of plantain potential growth and yield. The model allows both simulating different management strategies and estimating dry matter content of different plant organs. Its design also permits further incorporation of water, nutrient and pest limitations.
Materials and methods
A one cycle potential yield model of a new Dominico Harton plantation is presented. Total dry matter was estimated from the observed values of two experiments conducted from August 1991 to March 1993. One of them was planted in the locality of El Agrado, municipality of Montenegro, Quindío, Colombia (04°31' N; 75°49' W). The other one was carried out in the municipality of Palmira, Valle, Colombia (03°31' N; 76°19' W). Approximately 2 kg corms coming from needle type sprouts were planted at a density of 1,200 plants/ha, using a 3 x 3 m arrangement with only one plant per site. Three plants were monthly sampled during the nine months of the vegetative period; and then every two weeks during the four months of the reproductive period (after flowering). The three sampled plants were dissected into their constituting organs (roots, rhizome, pseudo stem, leaves, flower stem, and bunch) in order to measure fresh and dry matter. Climatic variables for both experiments were available from January 1991 to December 1993.
Several physiological variables were measured: plant height, perimeters, length, width and fresh and dry weight; number of current and emerged leaves, and fresh and dry weight of floral stem, bunch, fruit pulp and rind.
Non-linear regression analysis was applied to adjust the curve that describes and estimates leaf area index behavior. According to a study carried out by Turner (1994) on 30 cultivars, base temperature may vary from 10.3°C to 14.2°C. Thus, we set base temperature (Tb) at 12.5°C; and optimum and maximum temperatures at 25°C and 40°C, respectively. The model was developed in Microsoft Excel® software.
Description of the system and development of the model
As a perennial crop, plantain is featured by yield and physiological behavior of mother and daughter plants, each of them representing one production cycle. Even though the modeling process can be applied to previously existing plantations or to second or third production cycles, the complexity they imply is out of the scope of this work.
In modeling potential yield, it is assumed that the crop is only limited by temperature and radiation, implying that the plants are plenty of water and nutrients, and that no pests or diseases are present. Daily average temperature (°C) and radiation (MJ m-2 d-1), constitute the input of the model, while total dry matter (g/plant) is the output. Intercepted light estimation and light use efficiency (g MJ-1) are respectively an auxiliary variable and a parameter. Transformation of intercepted light into daily growth (g/ plant), is the main process to be modeled. Cumulative daily light interception rate, light use efficiency and daily accumulated solar radiation produce variations in daily accumulated total dry matter.
Model structure
A simple schematic representation of the model is shown in Fig. 1.
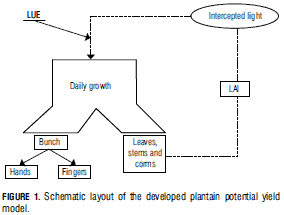
Light interception
The driving factor that determines plantain growth is light intercepted by the crop (F int), which depends on the amount of foliage and its distribution, that, in turn, vary along the growth cycle.
Depending on the specific crop tissue that is being modeled, and as it can be seen in equation 1, light interception can be described as:
where k is the extinction coefficient and LAI is leaf area index. The latter was calculated as the quotient that results from dividing total leaf area by 10,000 m2 (Spitters et al., 1989). Total leaf area is the product of leaf length (m) and width (m), times 0.80246 (Belalcázar, 1991). When light competition appears, intercepted light starts to decrease, which takes place around harvesting time. For such reason, this phase was considered relatively irrelevant to the model, and therefore excluded from it.
Growth
Expressed in dry matter production (g ha-1 d-1), daily growth was calculated as the product of intercepted radiation (MJ m-2 d-1), light use efficiency (LUE) and intercepted light fraction, which is expressed as g of dry matter per MJ of intercepted radiation. LUE integrates photosynthesis and respiration, so any factor that affects it will also affect these processes.
As it is shown below in equation 2, total daily growth is calculated as:
where DW is total daily growth as expressed in dry matter (g ha-1 d-1) and PAR is photosynthetic active radiation, which is equivalent to 50% of global radiation (Monteith, 1977; Gosse et al., 1986; Kooman, 1995; Jones and Luten, 1998).
Dry matter allocation
The model addresses a simple way to simulate daily dry matter allocation, consisting in only taking into account the reproductive organs to be harvested (the plantain bunch in this case) and the vegetative necessary organs that give the plant its strength and allow it to produce assimilates. Its approach to dry matter allocation consists in estimating each organ's fraction from total dry matter. Hence, the growth rate of each organ can be written as shown below in equations 3 to 6 (Marcelis, 1994; Kooman, 1995; Marcelis et al., 1998; Salazar et al., 2008):
where αl, αs and αf are the partition coefficients for leaves, stem and fruit, respectively.
Given that bunch biomass allocation only takes place after flowering, until then αf is equal to 0.
Dry matter simulation
Total daily dry matter was simulated through Euler's method (Van Kraalingen, 1995; Salazar et al., 2008), as presented below in equation 7:
where Wt is total dry matter (g m-2) at time t; Wt-1 is total dry matter at time t-1 (g m-2); dWt is dry matter daily growth rate (g m-2 d-1), and Δt is the time increment (1 day).
Results and discussion
Leaf Area Index (LAI)
Light interception was derived from LAI, which was calculated as total leaf area/10,000 m2 for 1,111 plants/ha. Thermal time was calculated using a Tb value of 12.5°C (Turner, 1994) and a weather database for the period between the planting date (01-02-1991) and the last measurement (25- 02-1992). A non-linear regression equation was fitted in order to estimate LAI as a function of time after planting, using a sigmoid logistic curve.
As it can be seen in equation 8, the predictive mathematical expression is:
where 12.9454 m2 is maximum LAI (m), 0.0306 is the slope of the curve (b), and 130.6 is the time in days during which LAI growth rate is maximum (c). RMSE was 1.5145; R2 was 98.10% and standard error values for m, b and c were respectively 0.6767, 0.000879 and 130.6 (Fig. 2).
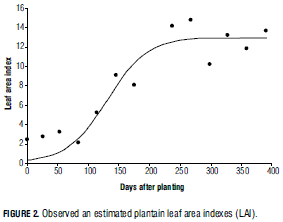
The precision of many photosynthesis models depends mostly on accurately predicting LAI, which is generally related to light interception. Two approaches have been frequently used to simulate leaf area development: (1) leaf area as a function of plant development, and (2) as a prediction from estimated leaf dry matter (Marcelis et al., 1998). LAI development is strongly influenced by radiation (Marcelis et al., 1998); for this reason, the first approach is frequently imprecise for greenhouse crops, due to fluctuations in radiation (annual crops). Nevertheless, some authors combine both methods, as is the case of Spitters et al. (1989) in annual species, De Visser (1994) in Onion, De Visser et al. (1995) in carrot, and Gijzen et al. (1998) in vegetables under greenhouse conditions.
In tomato and rose crops, Marcelis et al. (1998) have estimated LAI using constant values of specific leaf area (SLA) as a function of developmental stage or of source-sink relations. Basing LAI modeling on estimated leaf dry matter and SLA constitutes a more flexible approach, and has been applied in several crop models such as tomato (Heuvelink, 1999), lettuce (Van Henten, 1994), rose (Lieth and Pasian, 1991), chrysanthemum (Lee and Heuvelink, 2003) and oil palm (Awal et al., 2004). According to Lee and Heuvelink (2003), prior-to-canopy-closing LAI values have been frequently overestimated.
Estimation of total dry matter parameters
Light use efficiency (LUE)
An iterative non-linear optimization procedure was used to minimize the square sum of the differences between observed and predicted dry matter values, through a Microsoft Excel® solver tool. The minimum square sum was found when LUE = 1.63 g MJ-1. Regarding the American tropic, and specifically Colombia, Salazar et al. (2008) developed a cape gooseberry (Physalis peruviana) potential production model that describes dry mass production and distribution from the moment of planting to the end of the first harvest cycle. Their model presents LUE values of 0.46 g MJ-1 for the vegetative stage, and 2.62 g MJ-1 for the reproductive stage. In other studies, LUE (g MJ-1) has been observed to depend on crop management and sowing density, ranging from 2.1 to 3.2 in the case of potato (Kooman, 1995) and from 1.92 to 2.02 in peanuts, (Kiniry et al., 2005); similar measurements are 1.81±0.05 for wheat, 1.52 ±0.05 for pea and 1.92±0.12 for mustard (O'Connell et al., 2004). Kooman (1995) pointed out that LUE changes with plant development.
Intercepted light
According to equation 9, daily intercepted light was determined as:
where k = 0.2817 is the extinction coefficient and LAI was estimated by the logistic function. Fig. 3 shows intercepted light through time after planting. Approximately 150 d after planting, the foliage was intercepting close to 0,9 of global radiation.
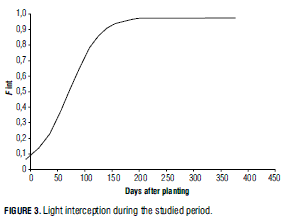
Extinction coefficient k depends on plant architecture and leaf position, angle and orientation. Several models have been developed in different crops to estimate this coefficient (Marcelis et al., 1994), whose values vary with planting density. In the case of coffee, for example, Castillo et al. (1996) reported different k values for different planting densities. For 2,500; 5,000; 7,500 and 10,000 coffee plants/ha, k values were respectively 0.46, 0.48, 0.60 and 0.61. Heuvelink (1995) reported k = 0.72 for tomato densities of 2.3 and 2.6 plants/m2. Carranza et al. (2008) found k values of 0.5120 and 0.5052 for two simple dry mass distribution models in broccoli (Brassica oleracea var. italica) and cabbage (Brassica oleracea), respectively.
Parameters for dry matter distribution to leaves, stem, corm and bunch
Plant developmental stage determines the pattern of dry matter distribution to different organs, and leaf area growth determines the light interception pattern (Kooman, 1995). As a simple way of simulating total dry matter distribution, the present model considers two stages: vegetative and reproductive. The vegetative stage corresponds to the production of leaves, corm and stem, which are necessary to form the plant and produce assimilates. In calculating biomass partition during the reproductive stage, flowers and bunch need also be considered, in addition to leaves, corm and stems. In doing so, the different organs of the plant were assumed to compete for assimilates, each of them representing a fraction of total dry matter, and varying with the physiological stages. This led to calculating the fractions independently for each organ at each stage.
In order to estimate the partition coefficient (αo), each organ's observed and estimated dry matter square sum was calculated and integrated into a single expression that assigns them all the same weight. Thus, the objective function to be minimized (equation 10) was:
where SSf, SSl, SSs and SSc are the respective square sums of the differences between observed and estimated bunch, leaf, stem and corm dry matter values; and S2 f, S2 l, S2 s , S2 c are the variances of the observed data.
As mentioned above, before flowering αf> equals 0. Consequently, the OF function was divided in two objective functions to be minimized: one for the vegetative stage, and another one for the reproductive stage (Salazar et al., 2008). For the vegetative stage, the OF function was minimized using the values of the estimated parameters αl=0.3387, αs=0.3550 and αc=0.3058. Except for the root, dry matter of leaves, stem and corm was found to increase in equal proportions during the vegetative stage, indicating that there was no selectivity for assimilates on the part of the plant organs. Even though during the reproductive stage almost all assimilates went to the bunch (implying that αf=1), the stem still exhibited some growth. The parameters of the reproductive stage were estimated with the same methodology.
For the reproductive stage, αl=-0.0121, αs=0.2009, αc=-0.2526 and αf=1.0639. The negative values of leaf and corm fractions imply that these organs lost mass because part of their assimilates were transferred to the bunch, and possibly to daughter plants and roots as well. Fig. 4 shows the amount of dry matter of each modeled organ. A negative relation can be clearly observed between dry matter accumulation in the bunch and loss of corm mass. While the leaves did not gain mass, the stem continued to increase. At the end of the experiment, bunch dry matter represented 45.28% of total aerial dry matter, to support which stem and root structures need to be sufficiently strong. The corm was observed to start losing dry matter before bunch formation, probably at flowering (Fig. 4). It is important to consider and measure the flowering process in order to include this organ in the model's explanation of the gap between the beginning of dry matter loss by the corm and the formation of the bunch.
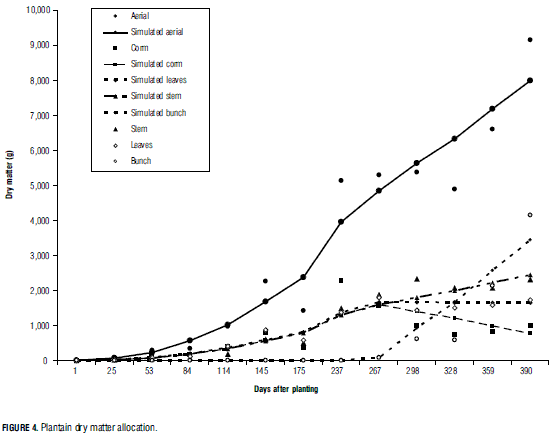
Although flowering usually marks the limit between the vegetative and reproductive stages, in the present work such limit was set at bunch inception. This date changeover resulted from the environmental conditions of the experiment sites, and is likely influenced by differences in stage of growth at the moment of planting. Salazar et al. (2008) found respective stem and leaf dry matter partition coefficients of 0.72 and 0.28 during the vegetative stage; and of 0.09, 0.23 and 0.69 for leaves, stem and fruits during the reproductive stage. Likewise, Carranza et al. (2008) reported respective partition coefficients of 0.4662, 0.4513 and 0.0825 for leaves, stems and root in the vegetative stage of broccoli; and of 0.1418, 0.3643, 0.2709, and 0.2230 for leaves, stems, root and flowers in the reproductive stage. In cabbage, such coefficients were 0.6530, 0.2841 and 0.0630 for leaves, stems and root, respectively.
Total dry matter sensitivity analysis
In order to determine parameter sensitivity for k, LUE and k*LUE, the model was run nine times, changing the values of the parameters to make up a 3*3 factorial array. As a result of the analysis of variance, LUE comes up as the most sensitive parameter, whose square sum represents 98.1% of the total sum squares; in turn, k represents 1.77%, and k*LUE, 0.13%. As well as in the case of total dry matter, LUE was observed to be the most important parameter accounting for fruit yield. In fact, within the variation range of LUE, yield was found to be directly proportional to it. In practice, all this means that it is important to conduct efforts aimed at improving CO2 capture through breeding and crop management, paying special attention to factors such as population density, planting system, planting date, appropriate cultivation zones and cultural management.
Great part of the assimilates accumulated in the corm until flowering supply bunch formation during the reproductive stage, so a timely cultural management aimed at eliminating organ competition should be carried out. Furthermore, stem partition coefficient was observed to increase from the vegetative to the reproductive stage, probably in order to support the bunch. In contrast, leaf dry matter proportion decreased considerably from the first to the second stage. Total dry matter response and uncertainty were linear with respect to K and LUE. Although larger values are desirable for these parameters, a suitable balance must be found between K, LUE, total dry matter and their variability.
The model presented here does not take into account heterogeneous development and growth. However, it predicts biomass trough time, and must be understood as representing the growth of a single plant during its first production cycle. In this sense, Tixer et al. (2004) report a probability based SIMBA-POP model, calibrated and validated to predict banana harvest, but not involving plant growth or development anyway.
Conclusions
Modeling plantain potential yield allowed determining and estimating some of the parameters that affect it, namely k, LUE, and total dry matter partition coefficients. The distributive model clearly shows the dynamics and processes of the crop, particularly focusing on the translocation of assimilates from the vegetative organs (especially the corm) to the bunch. In estimating the parameters, two stages were considered, which led to minimizing two objective functions, one for the vegetative stage, and another one for the reproductive stage. At the beginning of the reproductive stage (bunch inception), assimilate dynamics undergo a remarkable change when leaf dry matter stops increasing. Stem dry mater continues to increase, but corm dry matter decreases to transfer assimilates to the bunch, and possibly to the roots and daughter plants. Given that it contributes to understanding assimilate distribution dynamics, it is advisable to include flower dry matter in the model. Among the evaluated model's parameters, LUE comes up as the one that most sensitively affects yield. Further research should be aimed at improving CO2 capture and assimilate formation via crop management as well as plant breeding.
It is advisable to include more components in the model and to continue evaluating them through sensitivity analysis and validation processes. Once the complete potential growth model is ready, it shall be possible to include soil, water and nutrient limitations, as well as mass reduction caused by pests and diseases. Finally, with the aim of maximizing the profit, the economic component should also be included.
Literature cited
Awal, M.A., I. Wan, J. Endan, and M. Haniff. 2004. Determination of specific leaf area and leaf area-leaf mass relationship in an oil palm plantation. Asian J. Plant Sci. 3(3), 264-268. [ Links ]
Belalcázar, S. 1991. El cultivo del plátano en el trópico. Manual de Asistencia Técnica 50. ICA; Comité de Cafeteros de Colombia; CIID; INIBAP, Cali, Colombia. [ Links ]
Carranza, C., O. Lanchero, D. Miranda, M.R. Salazar, and B. Chaves. 2008. Modelo simple de simulación de distribución de masa seca en brócoli (Brassica sp.) variedad Coronado y repollo (Brassica oleracea) híbrido Delus cultivado en la Sabana de Bogotá. Agron. Colomb. 26(1), 23-31. [ Links ]
Castillo, E., A. Jaramillo, J. Arcila, and J. Sanabria. 1996. Coeficientes de extinción de la radiación fotosintéticamente activa en Coffea arabica L. Atmósfera 24, 65-70. [ Links ]
Dourado-Neto, D., D.A. Teruel, K. Reichardt, D.R. Nielsen, J.A. Frizzone, and O.O.S. Bacchi. 1998. Principles of crop modeling and simulation: I. Uses of mathematical models in agricultural science. Sci. Agric. 55, 46-50. [ Links ]
De Visser, C.L.M. 1994. ALCEPAS, an onion growth model based on SUCROS87: I. Development of the model. J. Hort. Sci. 69, 501-518. [ Links ]
De Visser, C.L.M., J.A. Schoneveld, and M.H. Zwart-Roodzant. 1995. Ontwikkeling van een gewasgroeimodel voor peen op basis van SUCROS87. PAGV-Verslag, Lelystad, The Netherlands. [ Links ]
Eckstein K., J.C. Robinson, and S.J. Davie. 1995. Physiological responses of banana (Musa AAA, Cavendish sub-group) in the subtropics. III. Gas exchange, growth analysis and source-sink interaction over a complete crop cycle. J. Hort. Sci. 70, 169-180. [ Links ]
Gary, C., J.W. Jones, and M. Tchamitchian. 1998. Crop modeling in horticulture: state of the art. Scientia Hort. 74, 3-20. [ Links ]
Gijzen, H., E. Heuvelink, L.F.M. Marcelis, E. Dayan, S. Cohen, M. Fuchs, and H. Challa. 1998. HORTISIM: A model for greenhouse crops and greenhouse climate. Acta Hort. 476, 441-450. [ Links ]
Gowen, S. 1995. Bananas and plantain. Chapman and Hall, London. [ Links ]
Heuvelink, E. 1995. Dry matter production in a tomato crop: measurements and simulation. Ann. Bot. 75, 369-379. [ Links ]
Gosse, G., C. Varlet-Grancher, R. Bonhomme, M. Chartier, J. Allirand, and G. Lemaire. 1986. Production maximale de matiére seche et rayonnement solaire intercepté par un couvert vegetal. Agronomie 6, 47-56. [ Links ]
INIBAP. 2001. Annual Report. Internacional Network for the Improvement of Banana and Plantain, Montpellier, France. pp. 53-56. [ Links ]
Jones, J.W. and J.C. Luyten. 1998. Simulation of biological processes. pp. 19-62. In: Peart, R.M. and R.B. Curry (eds.). Agricultural Systems. Modeling and Simulation, Marcel Dekker, New York, NY. [ Links ]
Keen, R.E. and J.D. Spain. 1992. Computer simulation in biology: a basic introduction. Wiley-Liss, New York, NY. [ Links ]
Kiniry, J.R., C.E. Simpson, A.M. Schubert, and J.D. Reed. 2005. Peanut leaf area index, light interception, radiation use efficiency, and harvest index at three sites in Texas. Field Crops Res. 91(2-3), 297-306. [ Links ]
Kooman, P.L. 1995. Yielding ability of potato crops as influenced by temperature and daylength. Ph.D. thesis. Landbouw Universiteit, Wageningen, The Netherlands. [ Links ]
Lentz, W. 1998. Model applications in horticulture: a review. Scientia Hort. 74, 151-174. [ Links ]
Lee, J.H. and E. Heuvelink. 2003. Simulation of leaf area development based on dry matter partitioning and specific leaf area for cut chrysanthemum. Ann. Bot. 91, 319-327. [ Links ]
Lieth, J.H. and C.C. Pasian. 1991. A simulation model for the growth and development of flowering rose shoots. Scientia Hort. 46, 109-128. [ Links ]
Marcelis, L.M.F. 1994. A simulation model for dry matter partitioning in cucumber. Ann. Bot. 74, 43-52. [ Links ]
Marcelis, L.F.M., E. Heuvelink, and J. Goudriaan. 1998. Modeling biomass production and yield of horticultural crops: a review. Scientia Hort. 74, 83-111. [ Links ]
Meira, S. and E. Guevara. 2000. Uso de modelos de simulación de cultivos como herramienta para la toma de decisiones en el cultivo de soja. In: Potash & Phosphate Institute, http://www.ppi-ppic.org; consulted: November 2009. [ Links ]
Monteith, J.L. 1977. Climate and the efficiency of crop production. Britain. Philos. Trans. R. Soc. Lond. 281, 277-294. [ Links ]
O'Connell, M.G., G.J. O'Leary, D.M. Whitfield, and D.J. Connor. 2004. Interception of photosynthetically active radiation and radiation-use efficiency of wheat, field pea and mustard in a semi-arid environment. Field Crops Res. 85(2-3), 111-124. [ Links ]
Peart, M.R. and R.B. Curry. 1998. Agricultural systems modeling and simulation. Marcel Dekker, New York, NY. [ Links ]
Ranganathan, R., Y.S. Chauhan, D.J. Flower, M.J. Robertson, C. Sanetra, and S.N. Silim. 2001. Predicting growth and development of pigeonpea: leaf area development. Field Crops Res. 69, 163-172. [ Links ]
Salazar, M.R., J. Jones, B. Chaves, and A. Cooman. 2008. A model for the potential production and dry matter distribution of Cape gooseberry (Physalis peruviana L.). Scientia Hort. 115, 142-148. [ Links ]
Spitters, C.J.T., H. van Keulen, and D.W.G. van Kraalingen. 1989. A simple and universal crop growth simulator: SUCROS87. pp. 147-181. In: Rabinge, R., S.A. Ward, and H.H. van Laar (eds.). Simulation and systems management in crop protection simulation monographs. Pudoc, Wageningen, The Netherlands. [ Links ]
Spitters, C.J.T. and A.H.C.M. Schapendonk. 1990. Evaluation of breeding strategies for drought tolerance in potato by means of crop growth simulation. Plant Soil 123, 193-203. [ Links ]
Thornley, J.H.M. and I.R. Johnson. 1990. Plant crop modeling: a mathematical approach to plant and crop physiology. Clarendon Press, Oxford, UK. [ Links ]
Tixer, P., E. Melézieux, and M. Dorel. 2004. SIMBA-POP: a cohort population model for long-term simulation of banana crop harvest. Ecol. Model. 180, 407-417. [ Links ]
Turner, D.W. 1994. Bananas and plantains. pp. 37-64. In: Schaffer, B. and P.C. Andersen (eds.). Handbook of enviromental physiology of fruit crops. Vol II: Sub-tropical and tropical crops. CRC Press, Boca Raton, FL. [ Links ]
Uthaiah, B.C., M. Indiresh, R. Jayarama, and K. Balakrisma. 1992. Performance of banana cultivars under indian west coast conditions. Agric. Res. J. Kerala 30(2), 84-88. [ Links ]
Van Henten E.J. 1994. Validation of a dynamic lettuce growth model for greenhouse climate control. Agr. Syst. 45, 55-72. [ Links ]
Van Kraalingen, D.W.G. 1995. The FSE System for Crop Simulation, Version 2.1. Quantitative approaches in system analysis No. 1. AB DLO/PE, Wageningen, The Netherlands. [ Links ]

