










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.27 no.3 Bogotá Sep./Dec. 2009
PROTECCIÓN DE CULTIVOS
Diana Arias1, Fernando Cantor1, 2, José Ricardo Cure1 y Daniel Rodríguez1
1 Programa de Biología Aplicada, Facultad de Ciencias, Universidad Militar Nueva Granada, Bogotá (Colombia).
2 Autor de correspondencia. ecologia@unimilitar.edu.co
Fecha de recepción: 8 de abril de 2009. Aceptado para publicación: 6 de noviembre de 2009
RESUMEN
Trabajos preliminares realizados en cultivos comerciales de rosa en la Sabana de Bogotá han permitido evidenciar la presencia de avispas del género Praon parasitando áfidos. Con el fin de explorar su potencial como controlador biológico, el presente estudio caracterizó la biología y el ciclo reproductivo de Praon sp. en condiciones de laboratorio bajo seis temperaturas constantes: 15, 17, 20, 25, 28 y 30°C. Áfidos parasitados fueron mantenidos en incubadoras a la temperatura deseada con 12 horas luz y 12 horas de oscuridad. Diariamente se realizaron disecciones de los áfidos parasitados para determinar el estado de desarrollo del parasitoide, lo que hizo posible determinar la duración promedio en días de cada estado, desde huevo hasta adulto. A todas las temperaturas, excepto a 15°C, se observó un desarrollo completo de los diferentes estadios, para los cuales se ajustaron modelos de tasa de desarrollo en función de la temperatura. El estado de huevo presentó el mayor umbral de temperatura mínima requerida para completar su desarrollo. Tanto hembras como machos son más longevos a temperaturas bajas. A temperaturas de 17 y 20°C se encontró mayor número de machos, mientras que a 25, 28 y 30°C se halló mayor número de hembras.
Palabras clave: control biológico, áfido, tomate, avispa.
ABSTRACT
Previous research studies conducted in commercial rose crops in the Bogota Plateau have shown the presence of aphids parasitized by wasps of the genus Praon. The present research studied the biology and reproductive cycle of Praon sp. under laboratory conditions, at six constant temperatures (15, 17, 20, 25, 28 and 30°C). The parasitized aphids were kept in incubators at the selected temperatures, under a regime of alternate light and darkness 12 hour periods. They were periodically dissected to determine the development stage of the parasitoid, which allowed determining the average duration of all such stages, from egg to adult. At all temperatures, except for 15°C, the results showed complete development of all biological stages, for which a mathematical model of development rate as a function of temperature was fitted. The eggs were observed to need the highest minimum temperature threshold to complete their development. Both males and females were observed to live longer at low temperatures. Regarding sex effect, females were found to live longer than males at all temperatures. A larger number of males was observed at 17 and 20°C, while females were more numerous at 25, 28 and 30°C.
Key words: biological control, aphid, tomato, wasp.
Introducción
Macrosiphum euphorbiae es una plaga con numerosos hospederos, entre los que sobresalen varias especies de flores y hortalizas (Eastop, 1983). Para el control de M. euphorbiae y de otros insectos considerados plaga en cultivos de flores de la Sabana de Bogotá, investigadores del Centro de Investigaciones y Asesorías Agroindustriales (CIAA-UJTL) y de la Universidad Militar Nueva Granada (UMNG) realizaron liberaciones de enemigos naturales para evaluar el potencial regulador de organismos biológicos introducidos desde Holanda. En dicho ensayo se evaluó la acción reguladora de Aphidius ervi sobre M. euphorbiae, pero su porcentaje de control fue muy inferior al de avispas del género Praon pos. occidentale que se presentaron de forma espontánea en los invernaderos donde se realizó el ensayo (Torrado et al., 2001). En trabajos anteriores a dicho ensayo, Lee et al. (1999) reportaron avispas del género Praon atacando áfidos que se encontraban infestando árboles presentes en los alrededores de invernaderos comerciales. El objetivo de este estudio fue el de evaluar algunos parámetros básicos de la biología de P. pos. occidentale con el fin de generar herramientas que permitan contribuir en el conocimiento de los requerimientos biológicos básicos de la especie.
En la literatura ya se ha reportado el potencial de avispas del género Praon para regular poblaciones de áfidos en programas de control biológico (Schlinger y Hall, 1960; Sigsgaard, 2000).
Los áfidos que han sido parasitados por Praon presentan una coloración blanca. Además, la pupa del parasitoide se desarrolla sobre un "pedestal" que lo separa de la base de la hoja y que lo diferencia de otros parasitoides de áfidos (Malais y Ravensberg, 1992). Las avispas del género Praon presentan tres estados larvales y su reproducción es biparental; de huevos sin fertilizar emergen machos, mientras que de huevos fertilizados emergen hembras (arrenotoquia). El macho del género Praon es a menudo mucho más oscuro que la hembra (Muesebeck, 1956).
Materiales y métodos
Establecimiento de la cría de Praon pos. occidentale
Se utilizaron plantas de tomate (Solanum lycopersicum var. Rocío) de ocho semanas de edad colocadas en jaulas de madera de 80 × 90 × 60 cm para el establecimiento de M. euphorbiae como hospedero de Praon sp. Posteriormente las plantas infestadas fueron trasladadas a cámaras de parasitismo de las mismas dimensiones, en donde se liberaban 15 avispas de P. pos. occidentale obtenidas de cultivos semicomerciales de tomate y de rosa mantenidos bajo invernadero. Cuando las hojas de tomate presentaban áfidos momificados de color blanco, se retiraban de la planta y se transferían a recipientes de plástico con 5 cm por cada lado y 12 cm de alto. Estas cámaras tenían una ventana de aireación en la cara superior, la cual estaba sellada con malla metálica. En estos recipientes se introducía un foliolo de ocho semanas de edad con el pecíolo inmerso en un tubo plástico ("aquapin"con capacidad de 7 mL) que contenía solución acuosa con preservante (acido cítrico más sacarosa en una concertación de 5 g L-1). Adicionalmente, la cámara tenía adherida dentro de una de sus paredes una tira de plástico amarillo impregnada de miel como fuente de carbohidratos para los parasitoides que emergían. En cada recipiente se ubicaron como máximo 100 áfidos por foliolo. Así mismo, se liberaban 40 avispas que eran retiradas después de 24 h.
Ensayos preliminares
a) Se identificaron categorías de edad de áfidos adultos para identificar la edad más susceptible al parasitismo por Praon sp.
b) Se determinó el tiempo en el cual P. pos. occidentale alcanza su madurez sexual. En este ensayo se utilizaron áfidos de edad adulta que fueron parasitados por avispas de 0-1, 1-2, 2-3 y 3-4 d de edad para comparar los diferentes porcentajes de parasitación.
c) Se reconocieron los diferentes estados biológicos de la avispa, desde huevo hasta adulto, por medio de la observación y disección de áfidos con diferentes tiempos de parasitación.
Las edades de los áfidos se determinaron de acuerdo con las características propuestas por Berlandier (2005), mientras que la determinación de desarrollo sexual y sexo de los individuos de P. pos. occidentale se efectuó mediante el análisis de las características del ovipositor y la genitalia del macho, como se describen en Schlinger y Hall (1960) y Marsh et al. (1987).
Estimación de parámetros de crecimiento de P. pos. occidentale en función de temperaturas
Para poder generar un modelo de tasas de desarrollo, es necesario evaluar el tiempo de desarrollo de los diferentes estados fenológicos en función de diferentes temperaturas, razón por la cual se emplearon temperaturas de 15, 17, 20, 25, 28 y 30°C en cámaras de cría. Se emplearon estas temperaturas por estar dentro del rango en que el parasitoide habita naturalmente. Previamente a la colocación de los parasitoides en estas cámaras, se realizó el proceso de parasitación en condiciones de laboratorio (22°C y HR de 55%). Para la parasitación se utilizó un recipiente de plástico rectangular de 17 cm de largo por 11 cm de alto y 14 cm de ancho, en el cual se colocaron dos foliolos de ocho semanas. A cada foliolo se añadieron 150 áfidos de edad adulta y se expusieron a 15 avispas con 24 h de emergidas en una proporción de siete hembras y ocho machos. Se adhirió dentro del recipiente una tira de plástico amarillo con solución azucarada. Este procedimiento se realizó en cinco repeticiones.
Se introdujeron las avispas en recipientes, que a su vez fueron distribuidos en cámaras climatizadas, las cuales permanecían con un fotoperiodo de 12 h luz y 12 h de oscuridad.
Después de la emergencia de los parasitoides, se hizo el seguimiento de su longevidad en cada una de las temperaturas.
Parámetros evaluados
A partir de los datos registrados, se cuantificaron las siguientes variables:
a) tiempo de desarrollo de huevo, larva, pupa y adulto en cada temperatura a través de una variación de la metodología de individuos-día (Carey, 1993);
b) tasa de desarrollo estimada como el inverso del tiempo promedio de desarrollo;
c) tiempo de desarrollo fisiológico, obtenido a partir del modelo del tiempo fisiológico;
d) temperatura mínima de desarrollo, estimada a partir del modelo del tiempo fisiológico (Campbell y Mackauer, 1975);
e) sobrevivencia y longevidad de adultos, determinada a cada temperatura, y
f) proporción sexual definida como la proporción de hembras en la progenie (Mackauer, 1983).
Análisis de datos
Se ajustaron dos modelos propuestos en la literatura para la predicción de las tasas de desarrollo en función de la temperatura. Se utilizo el modelo lineal (Gutiérrez, 1996) con el fin de obtener el umbral mínimo de temperatura y los grados día acumulados en cada estado de desarrollo, como se plantea en la ecuación (1).
donde R corresponde a la tasa de desarrollo, generalmente expresada en d-1 y T es la temperatura. Los parámetros a y b deben estimarse mediante regresión lineal. El tiempo requerido para que un individuo alcance un determinado estado de desarrollo a una temperatura especificada se puede obtener al calcular el inverso de la tasa de desarrollo para dicha temperatura. El umbral mínimo de temperatura requerido para el desarrollo del individuo se puede obtener como -a/b (Gutiérrez, 1996).
El modelo propuesto por Lactin et al. (1995) permitió obtener el umbral máximo de temperatura y la temperatura óptima de desarrollo para cada uno de los estados biológicos (ecuación 2).
donde r(T) corresponde a la tasa de desarrollo en función de la temperatura, ρ se interpreta como un valor Q10 compuesto para un conjunto hipotético de enzimas que son relevantes en el desarrollo del insecto (Logan et al., 1976), Tmax representa el umbral máximo de temperatura, por encima del cual el individuo no puede regular los procesos biológicos por periodos de tiempo prolongados, y Δ representa el rango de temperatura desde el óptimo termal hasta el umbral máximo de temperatura.
El ajuste de los modelos fue valorado mediante la prueba F del análisis de varianza para el modelo general, las pruebas de t para la significancia de cada uno de los parámetros y el R2; para los diferentes análisis fue utilizado el lenguaje estadístico R versión 2.6.2. Las comparaciones de longevidad se realizaron empleando análisis de varianza y pruebas de Tukey. Los datos de la proporción sexual fueron comparados mediante la prueba de Kruskal-Wallis (Walpole y Myers, 1992).
Resultados y discusión
Ensayos preliminares
Identificación de las categorías de edad de M. euphorbiae
Existen dos categorías de edad claramente reconocibles: los áfidos inmaduros (0-3 d de edad) en los cuales la longitud máxima de los cornículos apenas sobrepasa el extremo posterior de la cauda, y los áfidos adultos (4 d en adelante) cuyos cornículos sobrepasan la punta de la cauda (Berlandier, 2005).
Tiempo a la madurez sexual en Praon pos. occidentale
Al comparar los tratamientos (porcentajes de parasitismo) no se encontraron diferencias significativas (F(3,11) = 2,19, P = 0,145) en el número de huevos ovipositados entre el segundo a cuarto día de edad de la avispa. Es por ello que para los experimentos posteriores se usaron avispas con edad máxima de 2 d después de emergidas.
Influencia de la temperatura en diferentes parámetros de crecimiento de una población de P. pos. occidentale
Tiempo y tasas de desarrollo
La temperatura ejerce una influencia en la disminución de los tiempos de desarrollo de huevo (F(5,24) = 25,26; P = 7,84·10-9), larva (F(5,24) = 314,66; P = 4,70·10-21), pupa (F(5,24) = 65,58; P = 3,31·10-21) y adulto (F(5,24) = 345,36; P = 3,80·10-18) (Fig. 1). Además, no se alcanzó el desarrollo completo de las avispas a 15°C. Estas llegaron, como máximo y apenas en algunas de las repeticiones, hasta el estado de pupa. En relación con las tasas de desarrollo, tanto el modelo lineal como el modelo de Lactin permitieron obtener ajustes satisfactorios a los datos experimentales. En general se obtuvo un mayor coeficiente de determinación (R2) para el modelo de Lactin, cuyos valores estuvieron entre 0,93 y 0,98, mientras para el modelo lineal los valores de R2 variaron entre 0,76 y 0,98. De acuerdo con los resultados de Schlinger y Hall (1960), el tiempo desarrollo de P. palitans oscila entre 10,5 a 14,0 d a 21,0°C, sobre Therioaphis maculata, mientras que en este experimento Praon pos. occidentale requirió 20,26 d para completar su desarrollo a 20°C. En el trabajo de Campbell y Mackauer (1975) se reportan 26,71 d para la obtención de adultos de P. pequodorum a 14,8°C, a partir de áfidos Acyrthosiphon pisum, mientras en el presente trabajo no se obtuvieron individuos a 15°C. Según Hafez (1961), existe generalmente una relación parabólica entre la tasa de desarrollo y la temperatura para varias especies de himenópteros parasitoides de áfidos. Esa tendencia pudo ser observada en P. pos. occidentale , en donde la tasa de desarrollo aumenta progresivamente en 17, 20 y 25°C hasta alcanzar su máximo valor a 28°C, y en 30°C disminuye.
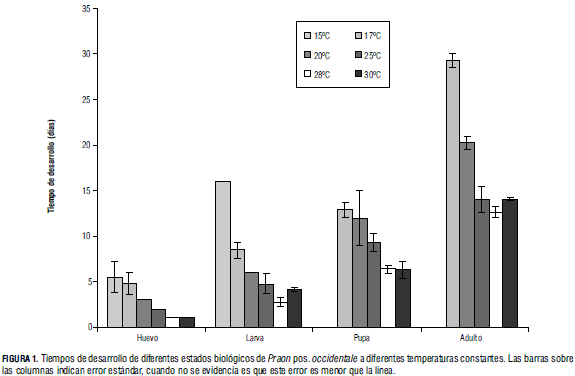
Parámetros del modelo lineal
Los parámetros estimados para el modelo lineal del estado adulto, que son el tiempo fisiológico y la temperatura mínima de desarrollo de huevo a adulto, se presentan en la Tab. 1. Los estados de pupa y adulto exhiben umbrales mínimos relativamente bajos (sin superar los 10°C), mientras que para los huevos y las larvas los umbrales son mayores (12°C aproximadamente). Por tanto, el establecimiento del parasitoide en la Sabana de Bogotá podría dificultarse, pues si bien la temperatura media de la región es de aproximadamente 13°C, se presentan oscilaciones desde 3°C hasta 23°C, e incluso temperaturas inferiores a cero en caso de heladas, lo que afectaría negativamente a los estados de huevo y larva, que presentaron una alta mortalidad asociada a temperaturas inferiores a 12°C.

En cuanto al requerimiento de unidades térmicas o grados día, se incrementa conforme mayor es el estado de desarrollo del insecto, como es de esperarse. Esto puede verse también en la pendiente del modelo lineal, que se reduce a medida que avanzan los estados de desarrollo, indicando menores tasas y por lo tanto mayores tiempos de desarrollo (Fig. 2).
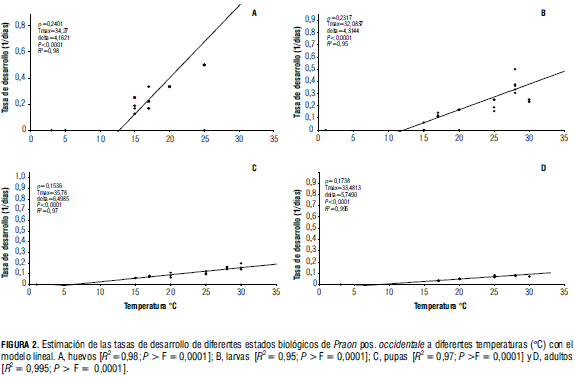
Por otro lado, en el experimento de Sigsgaard (2000), el tiempo fisiológico de huevo a adulto de P. volucre fue de 276,00 grados día, mientras que en este trabajo fue de 235,85 grados día para adultos de P. pos. occidentale . Aunque no hay una diferencia marcada entre estos dos valores, se podría explicar posiblemente por la biología diferente de ambos parasitoides, que hace posible que sus requerimientos térmicos sean distintos. Al comparar la temperatura mínima hallada en este trabajo con el obtenido por Campbell y Mackauer (1975), que fue de 6,9°C, hay una diferencia de 2°C aproximadamente, lo que se explica por las diferencias que existen en los requerimientos de temperatura entre las distintas especies de áfidos y sus parasitoides, como entre las poblaciones de algunas especies de diferentes áreas geográficas (Campbell et al., 1974). En cuanto al requerimiento térmico, Campbell y Mackauer (1975) reportan 199,60 grados día para llegar a adulto de P. pequodorum, mientras en este trabajo se estiman 235,85 grados día para alcanzar el estado adulto de P. pos. occidentale .
Parámetros del modelo de Lactin
Los valores de los tres parámetros estimados para el modelo de Lactin et al. (1995) se muestran en la Tab. 2. Los valores de ρ son un indicativo de la sensibilidad en términos de desarrollo en respuesta a cambios de temperatura. El mayor valor de ρ se presenta para los estado de huevo y larva, indicando que estos muestran una mayor respuesta en términos de tasa de desarrollo, al incrementarse la temperatura. Por el contrario, el parámetro ρ disminuye notablemente para los estados de pupa y adulto, por lo que se puede considerar que su desarrollo es afectado en menor medida por los cambios de temperatura, en el intervalo creciente de la curva.
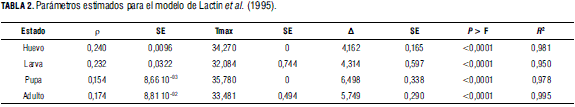
Los máximos termales oscilan entre 32 y 35°C para los diferentes estados de desarrollo (Fig. 3). Si se tiene en cuenta que en las condiciones de la Sabana de Bogotá las temperaturas en los invernaderos pueden fácilmente superar los 35°C, es factible que las poblaciones del parasitoide presenten mortalidad significativa en los diferentes estados de desarrollo, lo que compromete su función como controlador eficiente de los áfidos. Finalmente, al efectuar la resta entre los parámetros Tmáx -ΔT, se obtiene una aproximación de la temperatura óptima para el desarrollo de los diferentes estados. Dicho óptimo oscila entre 30,1°C para los huevos y 27,7°C para los adultos. Puesto que en la Sabana de Bogotá estas temperaturas solo se alcanzan durante ciertos momentos del día dentro de un invernadero y nunca a libre exposición, se puede suponer que para los diferentes estados de desarrollo del parasitoide las tasas estarán casi siempre por debajo del óptimo, en la fase creciente de la curva. Sin embargo, las amplias diferencias que se presentan entre los momentos más cálidos del día y los momentos más fríos de la noche pueden exponer al parasitoide a temperaturas bajas que pueden afectar negativamente el desarrollo y causar mortalidad en la población.
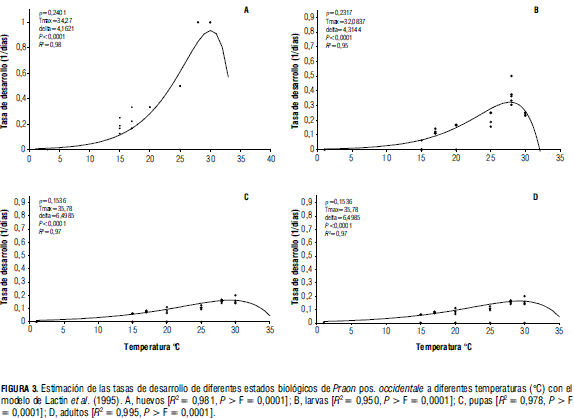
Aunque la temperatura que permite obtener máximas tasas de desarrollo está entre 27,7°C y 30,1°C, no necesariamente es esta la temperatura óptima para el establecimiento y crecimiento de las poblaciones del parasitoide, pues tales temperaturas pueden afectar negativamente otros procesos, como la fecundidad, la sobrevivencia de las hembras y la proporción sexual (Gutiérrez, 1996).
Longevidad
En la Fig. 4 se observa que la longevidad promedio tanto de hembras como de machos disminuye con el aumento de la temperatura. Al realizar el análisis estadístico se encuentra que la longevidad no difiere significativamente entre los dos sexos (F(4,4) = 2,65; P = 0,1838). Por el contrario, la longevidad difiere de forma estadísticamente significativa por efecto de la temperatura (F(4,8) = 6,17; P = 0,0144).
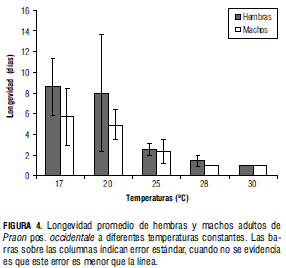
De acuerdo con el análisis de la Fig. 4, se puede interpretar que en condiciones frías tanto machos como hembras son más longevos que bajo temperaturas más altas, y que similar a lo que se presenta en el trabajo de Nealis y Fraser (1988) con la avispa Apanteles fumiferanae, la longevidad de las hembras es mayor que la de los machos en todas las temperaturas evaluadas. Sin embargo, esto no sucede siempre. Tal es el caso de Aphidius megourae, en el que las hembras fueron más susceptibles que los machos cuando se evaluaron bajas temperaturas (Hofsvang y Hagvar, 1975).
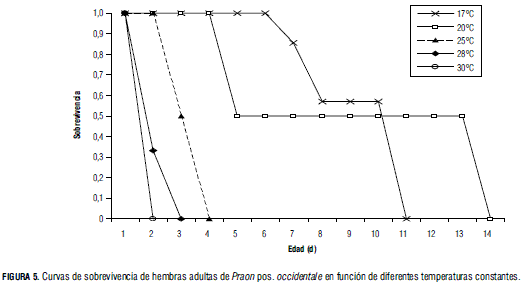
Sobrevivencia
En la Fig. 5 se encuentra que en la temperatura de 17°C se obtuvo una sobrevivencia superior a 0,8 hasta los 7 d, seguida por la sobrevivencia a 20°C, que cae a 0,5 a partir del día 5. A temperaturas superiores (25, 28 y 30°C) la sobrevivencia disminuye notablemente, siendo menor a un día en 30°C. Por tanto, la longevidad de las hembras es inversamente proporcional a la temperatura. Este fenómeno ya ha sido reportado en otras oportunidades por varios autores (Hofsvang y Hagvar, 1975; Jervis y Copland, 1996).
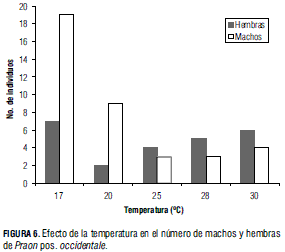
Número de machos y hembras
Al analizar la Fig. 6 se observa que no hay diferencias significativas entre el número de hembras emergidas en cada temperatura, aunque se presenta una mayor emergencia de machos en las temperaturas bajas. Estas observaciones fueron soportadas por la prueba de Kruskal-Wallis (P > X2 = 0,484) para hembras y (P > X2 = 0,0313) para machos. En la Fig. 6 se puede observar que en las temperaturas 17 y 20°C hay un mayor número de machos, mientras que en las temperaturas 25, 28 y 30°C hay un mayor número de hembras. Estos resultados coinciden con los resultados obtenidos por Sigsgaard (2000), quien registra que cuando se incrementa la temperatura de 16 a 25°C se obtiene un mayor número de hembras de P. volucre.
Consideraciones generales
Aunque se estimó el tiempo fisiológico del parasitoide y la temperatura mínima de desarrollo para conocer aspectos básicos de la biología de la avispa, es necesario tener en cuenta los requerimientos térmicos del hospedero, porque influirán en el éxito del control biológico realizado por P. pos. occidentale . De acuerdo con la bibliografía consultada, se ha encontrado que los requerimientos de temperatura de los parasitoides asociados con áfidos difieren de una manera similar, pero por regla general tienen requerimientos de temperatura más altos que sus hospederos (Campbell y Mackauer, 1975). Esta afirmación influye en que el parasitoide tenga un tiempo largo de desarrollo y sus generaciones requieran más tiempo que las generaciones del hospedero (Gilbert et al., 1976). Sin embargo, según Bernal y González (1993), es necesario tener presente que los parasitoides no deben tener umbrales mucho más altos que sus hospederos porque aparecerán cuando los hospederos sean muy abundantes y hayan alcanzado probablemente niveles de daño.
Conclusiones
El análisis de las tasas de desarrollo indica que entre 17 y 30°C se alcanza un desarrollo normal de las avispas del género Praon.
Si se quiere tener una cría de producción continua de P. pos. occidentale , es necesario considerar temperaturas entre 25 y 30°C, que permiten una mayor tasa de desarrollo y, por tanto, mayor número de ciclos de la avispa en un tiempo determinado.
El amplio rango de variación de temperatura en los invernaderos podría afectar negativamente a las poblaciones de Praon sp. y, por consiguiente, limitar su potencial de control.
Con el fin de ofrecer una alternativa efectiva y segura para el control biológico de áfidos con el uso de parasitoides, se sugiere considerar en futuras investigaciones:
a) el desarrollo de un método masivo que suministre constantemente avispas del género Praon en la región de la Sabana de Bogotá;
b) trabajos que permitan conocer la capacidad reproductiva de la avispa;
c) hacer liberaciones en campo para conocer la dispersión y migración del parasitoide;
d) la posibilidad de complementar la acción de Praon sp. con otros parasitoides y
e) identificación de la especie.
Agradecimientos
A la Universidad Militar Nueva Granada, a Asocolflores, por la financiación y el apoyo en este proyecto. Al Centro de Investigaciones y Asesorías Agroindustriales (CIAA) por el préstamo de sus instalaciones y sus servicios durante el tiempo de investigación.
Literatura citada
Berlandier, F. 2005. Identifiyng and managing aphids in potatoes. En: Department of Agriculture and food, Government of Australia, http://www.agric.wa.gov.au/PC_92871.html?s=1001; consulta: noviembre de 2009. [ Links ]
Bernal, J. y D. González. 1993. Temperature requirements of four parasites of the Russian wheat aphid Diuraphis noxia. Entomol. Exp. Appl. 69, 173-182. [ Links ]
Campbell, A., B.D. Frazer, N. Gilbert, A.P. Gutiérrez y M. Mackauer. 1974. Temperature requirements of some aphids and their parasites. J. Appl. Ecol. 11, 431- 438. [ Links ]
Campbell, A. y M. Mackauer. 1975. Thermal constants for development of the pea aphid (Homoptera:Aphididae) and some of its parasites. Can. Entomol. 107, 419-423. [ Links ]
Carey, J.R. 1993. Applied demography for biologists, with special emphasis on insects. Oxford University Press, New York, NY. [ Links ]
Eastop, V.F. 1983. The biology of the principal aphid virus vectors. pp. 115-132. En: Plumb, R.T. y J.M. Thresh (eds.). Plant virus epidemiology. Blackwell Scientific Publications, Oxford, UK. [ Links ]
Gilbert, N., A.P. Gutiérrez, B.D. Frazer y R.E. Jones. 1976. Ecological relationships. W.H. Freeman & Co., San Francisco, CA. [ Links ]
Gutiérrez, A.P. 1996. Applied population ecology: a supply-demand approach. John Wiley and Sons, New York, NY. [ Links ]
Hafez, M. 1961. Seasonal fluctuations of population density of the cabbage aphid, Brevicoryne brassicae (L.), in the Netherlands, and the role of its parasite, Aphidius (Diaretiella) rapae (Curtis). Tijdschr. Plziekt. 67, 445-548. [ Links ]
Hofsvang, T. y E. Hagvar. 1975. Duration of development and longevity in Aphidius ervi and Aphidius platensis (Hymenoptera: Aphidiidae) two parasites of Myzus persicae (Homoptera: Aphididae). Entomophaga 20(1), 11-22. [ Links ]
Jervis, M.A. y M.J.W. Copland. 1996. The life cycle. pp. 63-160. En: Jervis, M.A. y N.A.C. Kidd (eds.). 1996. Insect natural enemies practical approaches to their study and evaluation. Chapman and Hall, Oxford, UK. [ Links ]
Lactin, D.J., N.J. Holliday, D.L. Johnson y R. Craigen. 1995. Improved rate model of temperature-dependent development by arthropods. Environ. Entomol. 24, 68-75. [ Links ]
Lee, R., J.R. Cure y M. Pérez. 1999. Tan cercas y tan vivas. Asocolflores 56, 4-12. [ Links ]
Logan, J.A., D.J. Wollkind, S.C. Hoyt y L.K. Tanigoshi. 1976. An analytical model for description of temperature dependent rate phenomena in arthropods. Environ. Entomol. 5, 1133-1140. [ Links ]
Mackauer, M. 1983. Quantitative assessment of Aphidius smithi (Hymenoptera: Aphidiidae): fecundity, intrinsic rate of increase, and functional response. Can. Entomol. 115, 399-415. [ Links ]
Malais, M. y W.J. Ravensberg. 1992. Knowing and recognizing. The biology of glasshouse pests and their natural enemies. Koppert Biological Systems, Berkel en Rodenrijs, The Netherlands. [ Links ]
Marsh, P.M., S.R. Shaw y R. Wharton. 1987. An identification manual for the North-American genera of the genera of the family Braconidae (Hymenoptera). pp. 103-109. Memoirs of the Entomological Society of Washington. No. 13. The Entomological Society of Washington, Washington DC. [ Links ]
Muesebeck, C. 1956. Two new parasites of the yellow clover aphid and the spotted alfalfa aphid. Bull. 51. Brooklyn Entomol. Soc. pp. 25-28. [ Links ]
Nealis, V.G. y S. Fraser. 1988. Rate of development, reproduction, and mass- rearing of Apanteles fumiferanae (Hymenoptera. Braconidae) under controlled conditions. Can. Entomol. 120(3), 197-204. [ Links ]
Schlinger, E.I. y J.C. Hall. 1960. Biological note on Pacific coast aphid parasites and lists of California parasites (Aphidiinae) and their aphid hosts (Hymenoptera: Braconidae). Ann. Entomol. Soc. Amer. 53, 404-415. [ Links ]
Sigsgaard, L. 2000. The temperature-dependent duration of development and parasitism of three cereal aphid parasitoids, Aphidius ervi, A. rhopalosiphi and Praon volucre. Entomol. Exp. Appl. 95(2), 173-184. [ Links ]
Torrado, E., M. Pérez, J.R. Cure, M. García y C. Echeverri. 2001. Evaluación de sistemas de control biológico utilizados comercialmente en Europa, para el control de plagas de rosa bajo invernadero en la Sabana de Bogotá. Asocolflores 61, 37-47. [ Links ]
Walpole, R.E. y R.H. Myers. 1992. Probabilidad y estadística. 4a ed. McGraw-Hill, México DF. [ Links ]

