










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.27 no.3 Bogotá Sep./Dec. 2009
Guido Armando Plaza T.1, 3, Diego Nicolás Quintana V.1, Ligia Stella Aponte2 y Bernardo Chaves C.1
1 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá.
2 Biológicos y Fertilizantes, BAM S.A., Hacemos florecer, Bogotá (Colombia).
3 Autor de correspondencia. gaplazat@unal.edu.co
Fecha de recepción: 13 de febrero de 2009. Aceptado para publicación: 6 de noviembre de 2009
RESUMEN
Se llevó a cabo el reconocimiento y la caracterización de la comunidad de arvenses asociadas a un cultivo de rosa bajo invernadero, mediante un muestreo dirigido en el cual se consideraron algunos atributos poblacionales, tales como densidad, cobertura y frecuencia. Dichos atributos fueron evaluados en dos estilos de siembra (camas levantadas y confinadas), y adicionalmente en los bordes internos y externos de los invernaderos. Se registraron 46 especies pertenecientes a 41 géneros, 23 familias, 18 órdenes y tres subclases. El índice de diversidad (H) de la comunidad alcanzó un valor de 1,9. Por su parte, el índice de equidad ascendió a 0,497. Esto indica la alta dominancia de unas pocas especies, que fueron Cardamine hirsuta (dentro del invernadero, principalmente en las camas confinadas) y Pennisetum clandestinum (en los bordes del invernadero, sobre todo externos) las cuales contribuyeron con el 67% de la cobertura total. Estas fueron también las especies más abundantes. Se evidenció una clara diferencia entre los dos tipos de cama, y entre estos y los bordes.
Palabras clave: riqueza, estructura, diversidad, Cardamine hirsuta.
ABSTRACT
The community of weeds associated to a rose crop grown under greenhouse conditions was recognized and characterized by means of supervised sampling, analyzing a series of population attributes such as density, coverage and frequency. These were assessed in two different planting modes (confined and elevated beds), and additionally on the external and internal edges of the greenhouses. Forty six species belonging to 41 genera, 23 families, 18 orders and three subclasses were identified. Diversity index (H) of the community was 1.9, whereas equity index was 0.497, indicating the dominance of a few species, namely Cardamine hirsuta (inside the greenhouses, mainly in the confined beds), and Pennisetum clandestinum (outside the greenhouses, mainly on the edges), which responded for 67% of the whole plant coverage. These were also the most abundant species. A clear difference was detected between the two planting modes, and between them and the edges.
Key words: richness, structure, diversity, Cardamine hirsuta.
Introducción
La producción de flores para exportación ocupa cerca de 7.270 ha en todo el país, centrándose en la Sabana de Bogotá con 79%, Antioquia con 17%, y ciertas zonas del centro y occidente del país con 4%. La producción se destina principalmente a los mercados de Estados Unidos (80,44%), el Reino Unido (3,63%), Rusia (3,62%), Japón (2,51%), y el porcentaje restante (10%) se reparte entre otros países de América del Norte y Europa, principalmente. La importancia de este sector se puede medir por la cantidad de empleos generados, 98.641 directos y 85.533 indirectos para un total de 182.184 empleos. En este contexto la producción de rosas para exportación alcanzó 59.675 t, 29,7% de la producción total de flores para exportación, seguidas de clavel estándar con 31.846 t que representan 12,7% (Asocolflores, 2007).
Los valores de las exportaciones han aumentado de US$703 millones en 2004 a US$1.114 millones en 2007, con un incremento del 58% en los anteriores cuatro años; durante el periodo comprendido entre los años 2006 y 2007 el incremento porcentual fue de 15,3%. En la última década, las flores han aportado el 54,4% de las exportaciones de productos no tradicionales en la parte agrícola, US$7.058 millones, seguidas de banano con 40,3%. En el global de los productos no tradicionales, en el mismo periodo, las flores tienen un peso de 8,12% del total de exportaciones (Banco de la República, 2008).
Se ha definido como malezas a aquellas plantas que interfieren con los intereses humanos, sea por interferir en actividades económicas, establecerse en lugares o momentos indeseados. Este título es subjetivo ya que es propio del momento y del investigador (Zimdahl, 1999).
Los aspectos indeseables de las malezas son, entre otros, su capacidad de competir con las plantas de cultivo por factores como la luz solar, el agua y los nutrientes, lo cual lleva a aumentar los costos de protección, además de ser posibles hospedantes de plagas y patógenos. Con todo esto es muy probable que disminuyan la calidad de las cosechas, interfieran con la producción animal, reduzcan el valor de las tierras, afecten la salud humana y la calidad de las aguas (Zimdahl, 1999).
Una comunidad se describe como la articulación de las poblaciones de diversas especies presentes en el mismo espacio e intervalo de tiempo (Booth et al., 2003). Entre los principales atributos de una comunidad se encuentra la diversidad, abundancia y riqueza de especies. La diversidad se puede cuantificar mediante el índice de Shannon-Weiner (ecuación 1).
(H'= -Σ(Ni/N) * log (Ni/N)) (1)
en el cual se considera la cantidad de individuos de una misma especie, Ni, y el total de individuos presentes, N (Booth et al., 2003).
El estudio de las comunidades de malezas en sistemas de producción de flores en la Sabana de Bogotá ha estado más enfocado al estudio de la interacción entre organismos plaga, como ácaros y las malezas, que a la descripción y cuantificación de las comunidades de malezas. En un estudio hecho en 2004 en la Sabana de Bogotá se encontraron 40 especies de malezas relacionadas a tres cultivos de flores (rosa, clavel y alstroemeria). En rosa, particularmente, la riqueza de arvenses fue mayor en comparación con clavel y alstroemeria, 40 especies, de las cuales 36 hospedaban ácaros. Dos especies, Cardamine hirsuta y Poa annua, fueron las de mayor densidad dentro de los invernaderos bajo estudio (Triana et al., 2004).
Se ha definido como estrategia en la ecología de malezas, la explicación y la predicción de los niveles de abundancia de los componentes de cada población de malezas en las condiciones particulares de manejo en que se encuentren. Ello con el interés de poder proponer modelos de predicción de los niveles de poblaciones para las generaciones siguientes de plantas o ciclos de cultivo (González-Andújar y Hughes, 2000).
La caracterización de la comunidad de arvenses en el sistema productivo de rosa mediante la valoración de la riqueza y la estructura fue el objetivo del presente trabajo.
Materiales y métodos
Este trabajo se realizó en el municipio El Rosal (Cundinamarca) en la finca C.I. Splendor Flowers El Rosal, con un área total de 60 ha y cerca de 56 ha en producción de rosa, dividida en 116 invernaderos. En promedio, cada invernadero cuenta con 10 naves dentro de los cuales se emplean dos sistemas de siembra denominados confinado y levantado. El primer sistema "confinado" consta de la formación de camas que sobresalen del suelo en forma rectangular delimitadas por diversos materiales como angeo. El sistema "levantado" es la acumulación de suelo en forma de camas simples sin ninguna clase de material que lo delimite.
El muestreo realizado en los invernaderos fue sistemático tomando una cama y su camino adyacente. La ubicación de las unidades de muestreo fue la parte media de la cama intermedia en cada nave. Adicionalmente se tomaron seis unidades de muestreo en bordes exteriores y seis en los bordes interiores de los invernaderos.
Las unidades de muestreo en camas y caminos correspondieron a 1 m largo por el ancho de cada uno, y en los bordes correspondieron a cuadrados de dimensiones de 1,0 m de largo por 0,5 m de ancho. El porcentaje de área muestreado fue de 0,994% del total de la finca.
Entre las unidades de muestreo se registraron las especies de arvenses "riqueza", la densidad y la cobertura. A partir de los valores obtenidos se calcularon valores de frecuencia general (sin tener en cuenta el sitio de muestreo en el área de cultivo) y particulares (en camas levantadas y confinadas y los bordes internos y externos). Con base en las frecuencias en el área de cultivo (camas, caminos y bodes internos), las especies se agruparon en cuatro categorías: I: 0 a 25%; II: 25 a 50%; III: 50 a 75% y IV: 75 a 100%.
La comunidad de arvenses se caracterizó mediante los índices de diversidad y equidad de Shannon-Weiner para los tipos de siembra (confinada y levantada en camas y caminos) y bordes de invernaderos (externos e internos) (Ramírez, 2005).
Se efectuaron análisis de varianza y se aplicaron pruebas de comparación de medias de Tukey. Así mismo, se calcularon las posibles correspondencias entre ambientes y especies mediante análisis de correspondencias simples y elaboración de dendogramas en función de la similitud entre ambientes mediante el índice de Jaccard (Ramírez, 2005).
Las especies muestreadas fueron determinadas taxonómicamente, inicialmente en el Herbario de la Facultad de Agronomía y posteriormente confirmadas por botánicos especialistas en el Herbario Nacional de Colombia, Bogotá.
Resultados y discusión
Se registraron en total 46 especies, 41 géneros, 23 familias, 18 órdenes y tres subclases (Tab. 1).
La subclase más abundante fue Dicotiledónea, con 13 órdenes, 17 familias, 29 géneros y 33 especies, valores que porcentualmente corresponden a 75, 78, 76 y 76%, respectivamente, del total de las especies. El 22% de los géneros presentes pertenece a la familia Asteraceae, lo cual corresponde al 31% en lo relativo a las dicotiledóneas. Igualmente el 22% del total de las especies y 30% de las dicotiledóneas fueron asteráceas; otras familias importantes fueron Brassicaceae y Polygonaceae.
Por su parte, se registraron dentro de las monocotiledóneas 3 órdenes, 5 familias, 11 géneros y 12 especies, lo que significa el 16% de los órdenes, 21% del total de familias, 27% de los géneros y 26% del total de especies. La familia con mayor número de especies en esta subclase fue Poaceae, con seis especies o 51% de las especies monocotiledóneas y 13% del total de especies de la finca. Otras familias importantes fueron Cyperaceae e Iridiaceae.
La subclase con menor peso en la composición de la comunidad fue Polypodiopsida, la cual incluye solo un orden Polypodiales, una familia, Dryopteridaceae, un género y una especie.
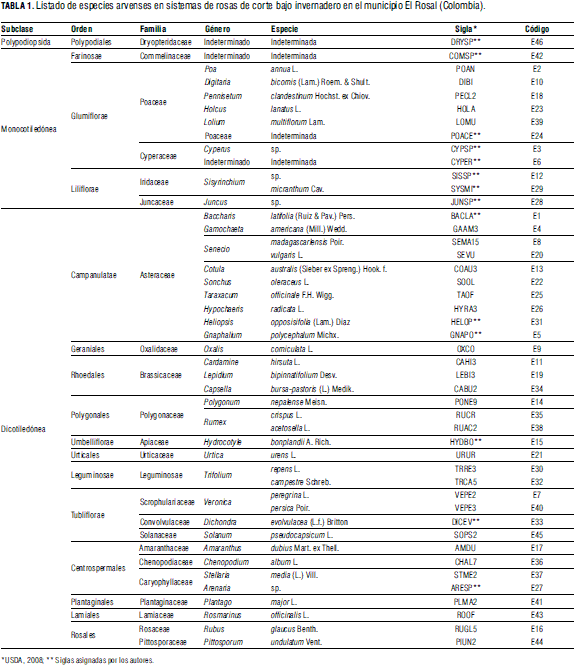
Los resultados del análisis de la variable frecuencia en el total de sitios muestran a Cardamine hirsuta y Pennisetum clandestinum como las especies más frecuentes (43 y 30%, respectivamente) seguidas en segundo lugar por Poa annua y Senecio vulgaris, 13% cada una; en tercera posición están Oxalis corniculata e Hydrocotyle bonplandii con 8% de frecuencia, y en cuarto lugar Trifolium repens con 6%. Rumex acetosella, Cotula australis y Cyperus sp. registraron valores de frecuencia entre 6 y 3%; las restantes especies fueron poco frecuentes (≤3%).
Los resultados destacan que Cardamine hirsuta y Pennisetum clandestinum se encuentran en todos los invernaderos. Del total de especies, 13 presentan frecuencias superiores al 50%, 24 superiores a 10% y las restantes 22 especies con frecuencias inferiores a 10% (Fig. 1).
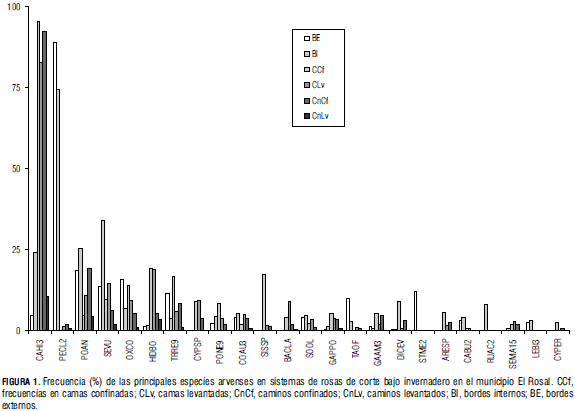
Los resultados del análisis en camas, caminos y bordes internos muestran que la categoría I agrupa 26 especies (57%), las categorías intermedias (II y III) agrupan 12 especies (15%) cada una, y en la categoría IV se ubicaron seis especies con el 13% (Fig. 2).
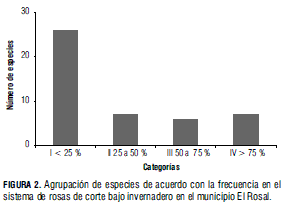
El análisis de varianza para la variable densidad indica diferencias significativas (P≤0,0001) para la mayoría de los ambientes; únicamente los bordes exteriores y los caminos levantados no presentan diferencias entre sí.
La densidad general en las camas fue 117 individuos/m2, y la especie de mayor densidad fue Cardamine hirsuta con 114,68 individuos/m2 (98% de los individuos). En cuanto a camas confinadas se encontró 232 individuos/ m2. Igualmente, la especie con mayor densidad fue Cardamine hirsuta con 228 individuos/m2. La densidad de especies acompañantes, como Arenaria sp., Sisyrinchium sp., Polygonum nepalense y Cyperus sp., varió entre 1,5 y 0,5 individuos/m2. En las camas levantadas, la densidad fue 80 individuos/m2; nuevamente Cardamine hirsuta es la especie con mayor densidad, 77 individuos/m2. Otras especies presentes fueron Senecio vulgaris 1,7 individuos/m2, Baccharis latifolia, 0,38 individuos/m2, Cyperus sp., 0,35 individuos/m2 y Cotula australis, 0,24 individuos/m2 (Fig. 3).
El número de individuos general, en los caminos, fue de 39 individuos/m2, y Cardamine hirsuta fue la especie más abundante con 38 individuos/m2, equivalente a 97% del total. Otras especies secundarias fueron Senecio vulgaris, Baccharis latifolia, Cyperus sp. y Cotula australis (Fig. 3).
Los caminos confinados presentaron 140 individuos/m2, y la especie de mayor aporte fue Cardamine hirsuta con 97,95% (136 individuos/m2); las especies acompañantes fueron Gamochaeta americana, Arenaria sp., Senecio vulgaris y Cotula australis (Fig. 3). En los caminos levantados la densidad fue de 7 individuos/m2 colocándose de nuevo Cardamine hirsuta como la especie de mayor peso, con 6,35 individuos/m2 (96%). En el grupo de especies acompañantes están presentes Senecio vulgaris, Cotula australis, Gnaphalium polycephalum y Baccharis latifolia.
El número de individuos en los bordes internos fue de 35 individuos/m2. La especie con mayor aporte fue Cardamine hirsuta con 19 individuos/m2 (54,62%) acompañada con Senecio vulgaris 10,13 individuos/m2; las otras especies con mayor importancia fueron Rumex acetosella, Cotula australis y Capsella bursa-pastoris. En el caso de los bordes externos, la densidad fue de 15 individuos/m2. Cardamine hirsuta, con 3,95 individuos/m2, fue la especie más relevante, seguida de Senecio vulgaris, con 3,53 individuos/m2 (Fig. 3).
Los resultados del análisis estadístico para la variable cobertura permiten apreciar diferencias altamente significativas (P≤0,001) en ambientes, donde el borde externo fue el de mayor cobertura y el camino levantado el de menor cobertura (Fig. 4).
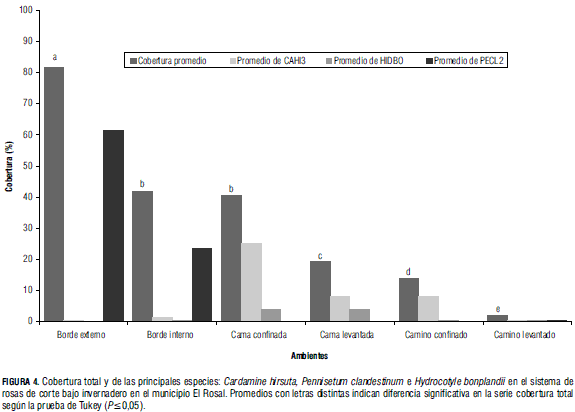
En las camas se registró una cobertura promedio de 24%. Las especies de mayores valores de cobertura fueron Cardamine hirsuta (54% de cobertura) e Hydrocotyle bonplandii con 18,54%. Otras especies de importancia fueron Hydrocotyle bonplandii, Oxalis corniculata, Senecio vulgaris y Trifolium repens, con promedios de 13,22, 4,47 y 1,33%, respectivamente. En las camas confinadas la cobertura de malezas fue 41% de cobertura; Cardamine hirsuta presenta el mayor valor con 25,67%, y le siguieron en importancia Hydrocotyle bonplandii, 4,01% de cobertura, Oxalis corniculata (1,93%), Trifolium repens (1,64%) y Polygonum nepalense (1,16%).
Las camas levantadas estuvieron cubiertas en 19%, inferior a la cobertura de las camas, 24%, y dos veces menor al promedio de las camas confinadas, 41%. Las especies con cobertura más abundante fueron Cardamine hirsuta (8,43%), Hydrocotyle bonplandii (4,5%), Senecio vulgaris (1,26%), Oxalis corniculata (0,98%) y Baccharis latifolia (0,6%). Los caminos estuvieron cubiertos solo en 7% y Cardamine hirsuta (2,32%) fue la especie dominante. Las especies acompañantes fueron Poa annua, Hydrocotyle bonplandii, Trifolium repens y Pennisetum clandestinum. La cobertura de malezas en los caminos confinados fue de 14%; nuevamente Cardamine hirsuta (8,14%) fue la especie con mayor cobertura, seguida de Poa annua (1,18%), Trifolium repens (0,73%), Hydrocotyle bonplandii (0,41%) y Gamochaeta americana (0,41%). En los caminos levantados la cobertura promedio fue 2,00%, siendo Hydrocotyle bonplandii (0,43%) la especie más relevante. Le acompañaron Cardamine hirsuta (0,37%), Pennisetum clandestinum (0,24%), Poa annua (0,19%) y Trifolium repens (0,12%). La cobertura en los bordes internos fue de 41,00%, donde Pennisetum clandestinum, con 23,26% de cobertura, fue la especie con mayor cobertura, seguida por Senecio vulgaris (4,74%), Poa annua (4,04%), Cardamine hirsuta (1,4%) y Rumex acetosella (1,14%).
Los bordes externos constituyeron el ambiente más densamente cubierto por malezas (81%). En este ambiente proliferó Pennisetum clandestinum (61,58% de cobertura); otras especies presentes fueron Poa annua (3,82%), Rumex acetosella (2,38%), Trifolium repens (2,34%) y Oxalis corniculata (2,25%) (Fig. 4).
El análisis de la comunidad incluyó la diversidad para toda el área de trabajo, esto es, sin diferenciar los ambientes. Este valor fue de H´ = 1,90, que indica baja diversidad, y se refleja en la dominancia de dos especies, Pennisetum clandestinum y Cardamine hirsuta, las cuales aportan 50 y 17% del valor de cobertura, respectivamente.
En las camas, Cardamine hirsuta fue la especie dominante, tanto en camas levantadas como en confinadas (45 y 66% de cobertura, respectivamente), siendo evidente su dominancia en las dos situaciones; la influencia de otras especies en la diversidad no es sobresaliente, dado el caso de las camas confinadas donde las siguientes cuatro especies con mayor cobertura únicamente representan el 21% de la cobertura. En los caminos confinados, Cardamine hirsuta es la especie de mayor relevancia, 60% de la cobertura, y en los caminos levantados la especie que se destaca es Hydrocotyle bonplandii, con 23,6%.
En las áreas de bordes, Pennisetum clandestinum es la especie de mayor relevancia en general, tendencia marcada a partir de los bordes externos. En los bordes internos la dominancia de Pennisetum clandestinum se mantiene, complementada por especies como Senecio vulgaris y Poa annua (Tab. 2).

La similitud entre los ambientes se calculó a partir de la composición florística o riqueza de especies, y se representa a manera de dendograma (Fig. 5). Los ambientes más parecidos entre sí fueron los caminos en general y los caminos confinados; por ello los componentes de los caminos confinados fueron más cercanos al ambiente camino, porque estaban inmersos en su totalidad en el ambiente general de camino; los caminos levantados se distanciaron de estos, pues presentan un número de especies bajo (19), en comparación a los caminos confinados, y existieron especies particulares en este ambiente que no se presentaron con regularidad en los demás (Fig. 5).
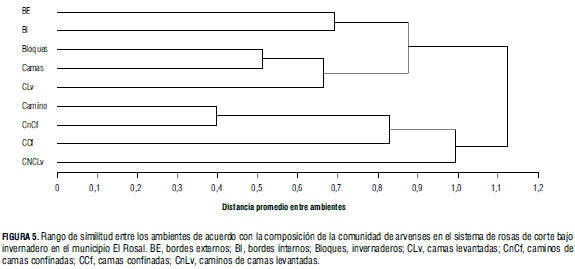
En la Fig. 5 se aprecia la clara semejanza entre los bordes, y se deduce de allí que las especies presentes en estos espacios, tanto interiores como exteriores, son las mismas; no se puede decir lo mismo de estas especies en relación con las áreas de producción como camas o sus aledañas.
El análisis de correspondencia muestra la proximidad entre especies, ambientes, o la ausencia de especies en un ambiente. El análisis de la Fig. 6 agrupa las respuestas en tres grupos: el primero incluye ambos tipos de cama y caminos confinados; el segundo, bordes interno y externo, y el tercero incluye los caminos levantados.
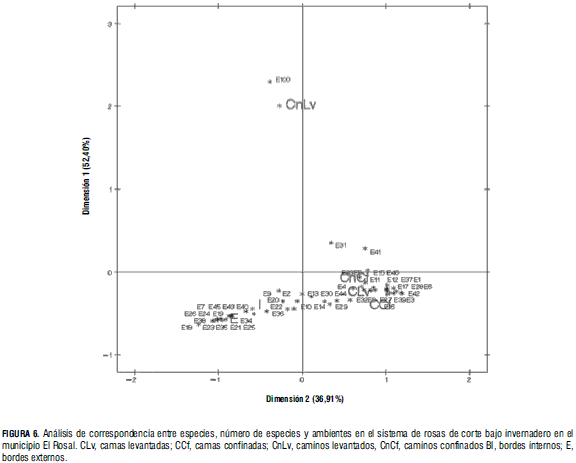
La correspondencia entre el primer grupo de ambientes y la frecuencia de las especies se presenta de forma aglomerada y con el mayor número de especies (25) que los restantes grupos. El nivel de correspondencia entre las especies y el segundo grupo de ambientes es más disperso y con presencia de menor número de especies (19). En el ambiente "camino levantado", la correspondencia con la ausencia de especies es marcada, relación que se da con sus valores de densidad y cobertura más bajos con relación a los demás.
De manera clara se aprecia la presencia casi permanente de algunas especies en los diferentes ambientes, Cardamine hirsuta, en las camas confinadas y levantadas además de ambos tipos de caminos y Pennisetum clandestinum en ambos tipos de bordes.
Discusión
La dominancia de Cardamine hirsuta puede ser debida a aspectos de su biología, entre ellos está la capacidad de tolerar ambientes de luz directa o de sombra indistintamente, su corto ciclo de vida (5 d para germinación y 5 semanas para completar su ciclo reproductivo), llegando a producir aproximadamente 5.000 semillas con porcentajes de germinación de 90%; estas plantas poseen un mecanismo de autodispersión mediante dehiscencia explosiva, que les permite a las semillas ser arrojadas a cerca de 1 m de distancia. Ésta especie ha demostrado su alta capacidad de adaptarse a ambientes diferentes al de origen y un ejemplo es la modificación en respuesta a condiciones de temperatura como lo reporta Kudoh et al. (2007) en Japón.
C. hirsuta es originaria de Europa y se reporta establecida como maleza en otros continentes convirtiéndose en parte de la flora acompañante de diversos sistemas productivos como lo son, sistemas de producción y propagación de plantas ornamentales en los Estados Unidos (Cross y Skroch, 1992), particularmente en el suroriente de este país, ambientes recién disturbados y cultivos de hortalizas en Japón (Yatsu et al., 2003). En ambas situaciones se relaciona la contaminación del suelo o sustrato, y su transporte como el factor de propagación más importante a larga distancia (Cross y Skroch, 1992; Yatsu et al., 2003). Otro aspecto relevante es ser reportada como especie hospedante de diversas plagas como ácaros (Triana et al., 2004).
Para el caso de la dominancia de Pennisetum clandestinum, ésta se debe al manejo agronómico buscando establecerlo como especie de cobertura, usando su hábito rastrero con tendencia a formar cespedones gruesos, haciendo uso de características como ciclo de vida, de donde se destaca su rápido crecimiento, planta de ciclo C4, en condiciones de temperaturas entre 15 y 30°C y altas humedades (Håkansson, 2003; Mateo, 2008). La presencia de estructuras como estolones y rizomas al igual que semillas, permite su rápida y alta diseminación (Mateo, 2008). Con respecto a sus semillas, estas pueden ser transportadas por el agua y animales, además de poder sobrevivir en el campo por más de 10 años. P. clandestinum es reportada como especie maleza importante en la mayoría de los países donde ha sido introducida (Håkansson, 2003; Mateo, 2008).
Conclusiones
En las condiciones del sistema de producción de rosa bajo invernadero se encontraron 46 especies malezas y dominancia de dos especies en particular Cardamine hirsuta y Pennisetum clandestinum.
En las áreas de producción, sean confinadas o levantadas, se presentaron 39 especies siendo Cardamine hirsuta la especie dominante; en las áreas bordes el número de especies fue menor (36) siendo Pennisetum clandestinum la especie dominante.
El caso de P. clandestinum como especie dominante combina el manejo que se le ha dado como cobertura vegetal y su capacidad intrínseca por la cual ha sido considerada una especie maleza muy importante.
Dentro de los diferentes sistemas de siembra, el sistema confinado es el que presenta mayor población de malezas y en especial de Cardamine hirsuta.
El sistema confinado tiene dos plantas de rosa más por m2, factor que implica mayores recursos, condiciones que favorecen la germinación, desarrollo y producción de mayor número de propágulos de las malezas.
La periodicidad en la ejecución de las prácticas de control de malezas afecta diferencialmente a las especies, y en caso particular del sistema de rosa bajo invernadero, éstas permiten a Cardamine hirsuta completar su ciclo de vida y a Pennisetum clandestinum desarrollar su sistema estolonífero.
De llegar a ser necesario el control de una ó de las dos especies dominantes, con el cambio en el método ó la periodicidad del mismo, podría ser suficiente para inducir cambios significativos en la estructura de la comunidad arvense.
Literatura citada
Asocolflores. 2007. Floricultura colombiana. Información general. En: Colombian association of flower exporters, http://asocolflores.org; consulta: septiembre de 2009. [ Links ]
Banco de la República. 2008. Principales productos no tradicionales de exportación (FOB). Información económica bases de datos sector externo, Banco de la República, Bogotá. [ Links ]
Booth, B., S. Murphy y C. Swanton. 2003. Weed ecology in natural and agricultural systems. Cabi Publishing, Wallingford, UK. [ Links ]
Cross, G.B. y W.A. Skroch. 1992. Quantification of weed seed contamination and weed development in container nurseries. J. Environ. Hort. 10, 159-161. [ Links ]
González-Andújar, J.L. y G. Hughes. 2000. Complex dynamics in weed populations. Functional Ecol. 14(1), 524-526. [ Links ]
Håkansson, S. 2003. Weeds and weed management on arable land. An ecological approach. CABI Publishing, Cambridge, UK. [ Links ]
Kudon, H., M. Nakayama, J. Lihova y K. Marhold. 2007. Does invasion involve alternation of germination requirements? A comparative study between native and introduced strains of an annual Brassicaceae, Cardamine hirsuta. Ecol. Res. 22, 869-875. [ Links ]
Mateo B., J.M. 2008. Prontuario de agricultura: cultivos agrícolas. Ediciones Mundi-Prensa, Madrid. [ Links ]
Ramírez, A. 2005. Ecología aplicada. Diseño y análisis estadístico. Universidad de Bogotá Jorge Tadeo Lozano, Bogotá. [ Links ]
Triana, R., A. Amado y A. Acosta. 2004. Arvenses hospedantes de ácaros plaga e cultivos comerciales de rosa, clavel y alstroemeria en la Sabana de Bogotá. Trabajo de grado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. [ Links ]
USDA, United States Department of Agriculture. 2008. Natural resources conservation service. En: www.plants.usda.gov; consulta: noviembre de 2009. [ Links ]
Yatsu, Y., N. Kachi y H. Kudoh. 2003. Ecological distribution and phenology of an invasive species, Cardamine hirsuta L., and its native counterpart, Cardamine flexuosa With., in central Japan. Plant Species Biol. 18(1), 35-42. [ Links ]
Zimdahl, R. 1999. Fundamentals of weed science. Academic Press, San Diego, CA. [ Links ]

