










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.28 no.1 Bogotá Jan./Apr. 2010
FISIOLOGIA DE CULTIVOS
Javier Orlando Orduz-Rodríguez1, 3, Hernán Javier Monroy2 y Gerhard Fischer2
1 Centro de Investigación La Libertad, Corporación Colombiana de Investigación Agropecuaria (Corpoica), Villavicencio (Colombia).2 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá (Colombia).
3 Autor de correspondencia. jorduz@corpoica.org.co
Fecha de recepción: 21 de septiembre de 2009. Aceptado para publicación: 5 de marzo de 2010
RESUMEN
En el trópico, la precipitación pluvial es el principal factor climático que determina el crecimiento y desarrollo de los cítricos, mientras que en el subtrópico lo son las modificaciones estacionales de la temperatura media. El registro detallado de los sucesivos estadios que determinan las fases fenológicas de los cítricos, tales como la brotación, la floración y el desarrollo de los frutos, permite estudiar la relación entre las variables climáticas de una región determinada y las respuestas fisiológicas de las plantas. Con el objetivo de documentar las respuestas del crecimiento y desarrollo de la mandarina ‘Arrayana’ en las condiciones climáticas del piedemonte del Meta, se realizó un estudio sobre la fenología de esta variedad en un cultivo ubicado en el C.I. La Libertad, Villavicencio (4°03’ N, 73°29 W; 336 msnm). En árboles de mandarina ‘Arrayana’ de 7 años de edad, se evaluó el estadio fenológico de las ramas terminales, y se correlacionó con la precipitación mensual durante un año. Las plantas presentaron tres picos de brotación (marzo-abril, julio-agosto y septiembre-octubre). La principal floración anual acompañó la primera brotación, la cual inició 2-3 semanas después del comienzo de la época lluviosa, pero también se presentaron flores en los meses lluviosos en ausencia de estrés hídrico.
Palabras clave: Citrus reticulata, ecofisiología, estrés hídrico, inducción floral, trópico bajo.
ABSTRACT
Under tropical conditions the main climatic driver of citrus plant growth and development is rain precipitation, whereas in the subtropics it corresponds to seasonal average temperature variation. Thus, the detailed observation and recording of the main phenological stages of citrus trees, namely budding, flowering and fruit development, allows studying the existing relation between climatic variables and plant eco-physiological responses. The present study was conducted at research center ‘La Libertad’, Villavicencio (4°03’ N, 73°29’ W; 336 m a.s.l.), located in the eastern piedmont of the department of Meta, Colombia. In a 7 year old ‘Arrayana’ mandarin orchard, the relation between phenological stage of trees and monthly precipitation pattern was evaluated. The citrus trees displayed three major budding peaks (March-April, July-August and September-October). Starting 2-3 weeks after the beginning of the rainy season, the main annual flowering accompanied the sprouting of the first buds. Notwithstanding, the trees also flowered during the rainy season, when they were not under water stress.
Key words: Citrus reticulata, ecophysiology, water stress, flower induction, lowland tropics.
Introducción
La ‘Arrayana’ es la principal variedad de mandarina (Citrus reticulata Blanco) cultivada en Colombia y en la región de los Llanos Orientales, donde existen alrededor de 1.000 ha ubicadas casi en su totalidad en el piedemonte del Meta. La ‘Arrayana’ desarrolla un fruto grande, cuyo peso oscila entre 160 y 190 g, con semillas y de fácil pelado (Orduz, 2007). Es una variedad de entrada tardía con alternancia de cosechas inducida por el comportamiento de precipitaciones extemporáneas durante la época seca. Los estudios publicados acerca de esta variedad han analizado su comportamiento como injerto sobre diferentes patrones (Orduz et al., 2006), su rendimiento durante un periodo de doce años en la región del Ariari (Orduz et al., 2007) y la influencia del estrés hídrico en la inducción floral (Orduz y Fischer, 2007). En el piedemonte del Meta esta variedad presenta una sola floración al inicio del ciclo de lluvias, y una alta concentración de cosecha al finalizar el año (noviembre a enero), lo que ocasiona una fuerte disminución de precios para los productores y desabastecimiento de fruta para las ciudades de la región durante los restantes meses del año.
En cítricos, el crecimiento y desarrollo del mandarino están determinados por la dotación genética de la planta injertada, por su interacción con el patrón utilizado y por las condiciones climáticas de la zona de cultivo (Agustí, 2003). Siendo los cítricos plantas de día neutro (Davies y Albrigo, 1994), la brotación y el crecimiento de los tallos son procesos que dependen de las condiciones ambientales (Jackson, 1999). Específicamente, las bajas temperaturas y el estrés hídrico constituyen los factores inductivos primarios de los cítricos (Agustí, 2003).
Es así que en el subtrópico el inicio de estos procesos está regulado por el incremento de la temperatura (>12,5°C), que se presenta al comienzo de la primavera, mientras que en el trópico depende de la disponibilidad de agua (Cassin et al., 1969). En efecto, allí el principal factor de inducción de la floración es el estrés hídrico, el cual determina el momento y la intensidad, así como la duración y distribución de la floración (Cassin et al., 1969; Reuther, 1973; Davenport, 1990). En un cultivo de naranja ‘Valencia’ en São Paulo, Brasil, Prado (2006) reporta una floración intensa cuando, durante la fase de inducción de la misma, la disminución de la temperatura se presentó acompañada por un momento de estrés hídrico (Prado, 2006).
Tanto la elongación estacional de los tallos, como la acumulación de materia seca, suelen ser mayores en el trópico bajo húmedo, donde hay días de brillo solar constante durante todo el año, acompañados de temperaturas altas, tanto diurnas como nocturnas (Davies y Albrigo, 1994). Los mismos autores consideran que en estas regiones se cultivan las plantas de mayor tamaño de todas las regiones citrícolas del mundo.
El meristemo vegetativo de la yema apical del brote de los cítricos forma hojas, yemas axilares y espinas a intervalos regulares. Los puntos donde se forman estas estructuras son los nudos. Los tallos, al elongarse, hacen que los nudos se alejen entre sí, contando para esto con un meristemo intercalar en la base de cada nudo. Las yemas axilares dan origen a las brotaciones regulares (Tadeo et al., 2003).
En condiciones subtropicales, la brotación y elongación de los tallos de los cítricos presenta de dos a cinco picos de crecimiento anual, mientras que en condiciones tropicales estos ocurren con diferente intensidad y a intervalos variables, dependiendo del régimen de lluvias y del manejo del cultivo, en especial de la fertilización con nitrógeno.
En el Mediterráneo español (hemisferio N), la mandarina presenta tres brotaciones importantes en el año: una en primavera, entre marzo y abril, otra en el verano, entre junio y julio, y la final en el otoño, entre septiembre y octubre (Tadeo et al., 2003). En estas condiciones, la brotación principal (dada su importancia comercial) tiene lugar en primavera, produciéndose muchos brotes con entrenudos cortos, mientras que en verano y otoño los brotes son largos y tienen pocos puntos de crecimiento (Tadeo et al., 2003).
El estudio de los eventos periódicos naturales involucrados en la vida de las plantas se conoce como fenología (Volpe, 1992). Las observaciones fenológicas proporcionan información básica sobre el comportamiento de las distintas especies de plantas en las regiones específicas en que se desarrollan (Azkue, 2009). El registro pormenorizado de los estadios sucesivos que detallan las fases fenológicas de los cítricos (brotación, floración y desarrollo de los frutos), permite analizar la relación entre el clima y la respuesta de la planta frente a las condiciones locales.
Con el fin de contribuir al conocimiento de los principales eventos fenológicos de la mandarina ‘Arrayana’ en las condiciones climáticas del piedemonte del Meta, estos fueron cuantificados durante un ciclo anual, describiéndose además su relación con la precipitación. Paralelamente se adaptó una metodología para la descripción de procesos fenológicos en condiciones tropicales. Esta información es de interés para estudios de epidemiología, nutrición mineral, riego, fisiología del crecimiento y desarrollo de la variedad ‘Arrayana’ en la región, al tiempo que proporciona elementos para comprender el crecimiento y desarrollo de los cítricos en condiciones tropicales.
Materiales y métodos
El estudio se inició en el mes de enero de 2004, y continuó hasta enero del año siguiente. Durante este periodo se cumplió un ciclo de brotaciones que comprendió desde el reposo de la época seca durante la transición 2003-2004, hasta la siguiente época seca entre 2004-2005. Las condiciones climáticas registradas durante el ciclo de cultivo se presentan en la Tab. 1. El estudio se realizó en el Centro de Investigación La Libertad de Corpoica, en Villavicencio (Meta), ubicado a 4°03’ N, 73°29 W, a una altitud de 336 msnm, en un cultivo de mandarina ‘Arrayana’ de 7 años de edad, injertado sobre mandarino ‘Cleopatra’, con una distancia de plantación de 8 x 5 m.
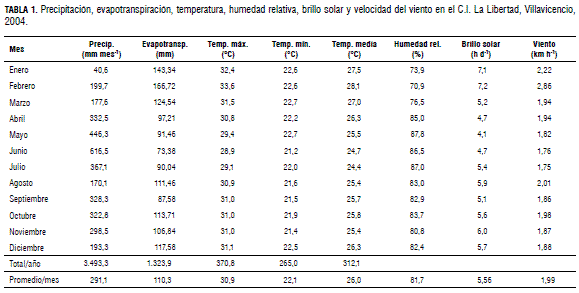
El tipo de suelo del área experimental se clasifica como Typic Haplustox, de textura franco arenosa (FA). En la clasificación regional estos suelos se incluyen en la clase IV (terrazas altas), la cual es recomendada para cítricos en la zona (Orduz y Baquero, 2003). En la cosecha inmediatamente anterior a la realización del estudio (2003-2004) el cultivo presentó un rendimiento de 8 t ha-1, que, dado el patrón en que está injertada, se considera moderado pero apropiado para la variedad.
En cada lectura se evaluaron cuatro árboles seleccionados al azar, siguiendo la escala y la metodología propuestas por Garrán et al. (1993). En cada árbol se escogieron aleatoriamente diez ramas de cada uno de los cuatro puntos cardinales. En cada rama las observaciones se realizaron en un segmento de 30 cm medidos a partir del ápice. Allí, de acuerdo con las categorías definidas en la Tab. 2. Se registró el estadio fenológico predominante entre los brotes vegetativos y entre los brotes reproductivos. Este proceso se repitió cada dos semanas, desde el 16-01-2004 hasta el 12-01-2005, con excepción de la segunda semana de marzo, en que no se llevó a cabo.
Para el cálculo del requerimiento hídrico del cultivo (ETc) se utilizó la metodología propuesta por la FAO (2006), a través de la fórmula1:
Donde Kc es 0,75 para los meses lluviosos y 0,80 para los meses secos (FAO, 2006). Los promedios de evaporación mensual correspondientes al periodo comprendido entre octubre de 2003 y abril de 2005 se tomaron de la estación climatológica del Ideam, ubicada en el C.I. La Libertad.
El tratamiento estadístico de los datos se llevó a cabo por medio de procedimientos descriptivos. Se realizaron 17 evaluaciones a lo largo del año, en cada una de las cuales, tras la toma de información en campo a partir de las ramas muestreadas, se totalizó el número de ramas en las que predominaba cada uno de los distintos estadios, y con base en ello se calculó su porcentaje correspondiente. Así, en cada lectura se obtuvieron dos series de porcentajes, una para los estadios de los brotes vegetativos y otra para los estadios de los brotes reproductivos. Posteriormente se agruparon todos los estadios en una sola escala de categorías fenológicas que condensan la información acerca de los brotes vegetativos y reproductivos (Tab. 2), con el fin de representarla gráficamente de forma cronológica. Finalmente, se estableció una relación descriptiva entre el comportamiento de la precipitación a lo largo del año y el ciclo fenológico del cultivo.

Resultados y discusión
Balance hídrico
El requerimiento hídrico anual de los cítricos en el piedemonte del Meta oscila alrededor de 1.046 mm al año (Orduz y Fischer, 2007). En la Fig. 1 se presenta el requerimiento hídrico del cultivo estudiado en el presente trabajo durante el año de su realización. A finales de 2003 se presentó un déficit hídrico de más de dos meses, mientras que en 2004 hubo un exceso de precipitación (desde el punto de vista del abastecimiento del cultivo) que se prolongó por varios meses, desde febrero hasta diciembre. El estrés ocasionado por el periodo de déficit hídrico es el principal factor de inducción de la floración, la cual, para esta variedad, y dadas las condiciones del estudio, se presenta 15 días después del inicio de las lluvias (Orduz y Fischer, 2007). La floración como respuesta de la planta ante el déficit hídrico estaría internamente regulada por la disminución de los niveles de giberelinas y su transporte hacia la copa, lo cual es resultado de la detención del crecimiento de la raíz, que, junto con el fruto, son los principales órganos de síntesis de esta hormona (Monselise, 1985; Agustí, 2003).
Brotación y crecimiento vegetativo
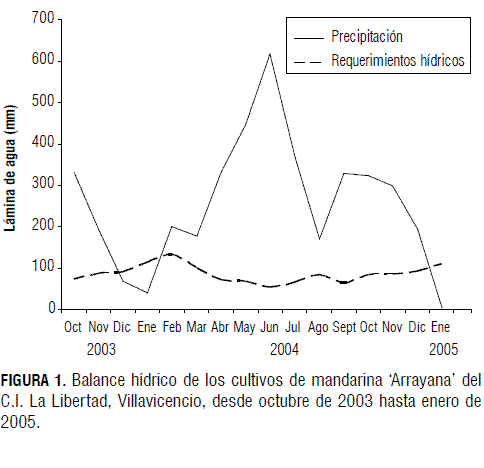
El principal momento de brotación del cultivo estudiado se dio al inicio de la temporada de lluvias, momento en el cual todas las ramas evaluadas (100%) presentaron al menos un brote. En relación con el origen de los fotoasimilados utilizados en la brotación y elongación, Erner y Bravdo (1983) reportan que las hojas nuevas son importadoras de carbono durante un periodo de 4 a 6 semanas después de la floración (madurez de las hojas). Durante esta etapa se presenta una competencia por fotoasimilados entre las hojas en desarrollo (Fig. 2) y los frutos que se encuentran en la fase I (Fig. 3), durante la cual se lleva a cabo la proliferación celular en los diferentes tejidos del fruto. Después de la maduración de las hojas se evidencia un incremento en la asimilación de CO2 por parte de estas durante los seis meses siguientes, la cual permanece estable en la madurez y declina en la vejez. La duración de las hojas oscila entre dos y tres años, dependiendo del vigor de la planta y de sus problemas fitosanitarios (Davies y Albrigo, 1994). La información de la Fig. 2 indica que durante la época lluviosa las plantas presentan una abundante producción de brotes, que disminuye con la llegada de la época seca.
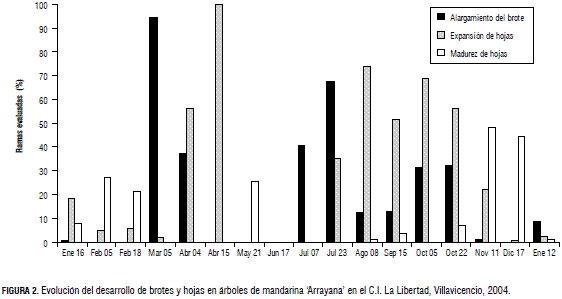
En la Fig. 3 se observa que en el año de estudio el cultivo desarrolló brotes vegetativos durante un periodo de nueve meses. La menor brotación ocurrió en enero, mientras que en marzo, después del inicio de la temporada lluviosa, más del 90% de las ramas evaluadas presentaron brotes. Durante los meses de mayo y junio quedaron suspendidas las brotaciones, para reactivarse en julio, agosto, septiembre y octubre, con porcentajes mensuales de ramas en brotación del 54, 13, 13 y 32%, respectivamente. En la Fig. 3 se advierte que las plantas presentaron tres picos de brotación en el transcurso del año, lo cual está relacionado con el comportamiento monomodal de la precipitación en el piedemonte del Meta.
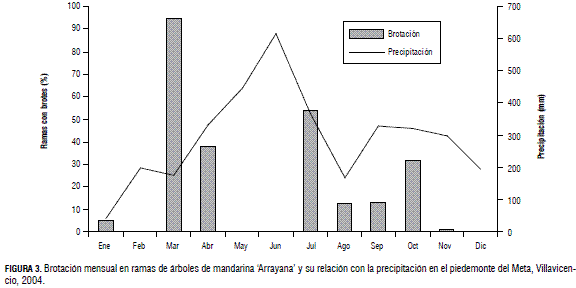
El comportamiento de la brotación vegetativa es parecido al que presentan los cítricos en condiciones subtropicales, en las cuales se obtienen de tres a cuatro picos de crecimiento (Davenport, 1990; Spiegel-Roy y Goldschmidt, 1996; Agustí, 2003; Tadeo et al., 2003). Este comportamiento contradice el reporte de Mendel (1969), quien menciona que el crecimiento de los cítricos en el trópico presenta una producción de brotes permanente durante todo el año. Es probable que tanto la brotación como su intensidad cambien con los diferentes regímenes de precipitación en las diversas regiones citrícolas del país, en especial las de tipo bimodal.
Floración y cuajado
El análisis de la floración a lo largo del ciclo anual estudiado permite observar cómo el estrés hídrico es el principal inductor de este proceso en cítricos en condiciones tropicales, coincidiendo con reportes anteriores (Reuther, 1973; Davenport, 1990; Davies, 1997), y como también lo documentan Orduz y Fischer (2007) para la mandarina ‘Arrayana’ en el piedemonte llanero.
Para las plantas en estudio, el estrés hídrico se inició a finales del mes de noviembre, y se prolongó hasta la primera semana de febrero. Así, la brotación de la primera semana de marzo constituyó la principal floración del año, mientras que las últimas flores abiertas y el final de la antesis se registraron en la primera semana de abril (Fig. 4). Este comportamiento floral coincide con lo reportado por Stover et al. (2002), quienes afirman que los cítricos en las zonas tropicales florecen después de un periodo seco seguido por la presencia de lluvia o aplicación de riego.
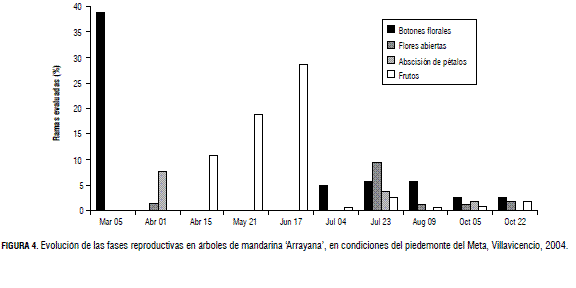
Adicionalmente, se presentaron floraciones de menor intensidad durante el invierno, acompañando las brotaciones de julio y agosto (con un 6% de las ramas evaluadas y en octubre con un 3%). Como se puede observar, ello ocurrió en ausencia de estrés hídrico (el principal promotor de la floración) y de frutos en maduración (que han sido reportados por Guardiola [1997] como inhibidores de la floración). Si bien se desconocen los factores relacionados con la inducción de estas yemas extemporáneas, su producción inicial pudo haber sido determinada durante el periodo de estrés hídrico de la época seca, pero su brotación habría sido inhibida por la presencia de otras yemas o de frutos en desarrollo. Alternativamente, cabe pensar que la inducción de estas yemas florales ocurrió en ausencia de estrés hídrico. En tal caso, el proceso podría estar regulado por otros factores endógenos relacionados con el balance de hormonas en las ramas con flores extemporáneas.
Existe evidencia de que los brotes jóvenes, los frutos en desarrollo y los ápices radicales son eficientes productores de giberelinas (Gil, 2000). De otra parte, aplicaciones exógenas de este fitorregulador han inhibido el proceso de inducción floral (Davenport, 1990; Guardiola, 1997). En zonas subtropicales, plantas de mandarina sometidas a estrés hídrico severo (-1,5 a -2,0 MPa) durante el otoño, redujeron el número de nudos florales simultáneamente con un aumento de la concentración de AG3 (Koshita y Takahara, 2004). Tal y como se ha estudiado en condiciones subtropicales, es probable que en el caso del trópico las giberelinas también ocupen un papel central en el proceso de inducción de este tipo de floración, teniendo incidencia únicamente sobre las ramas o partes de estas que presentan floraciones extemporáneas.
Los cultivares de cítricos de mayor importancia comercial presentan una floración abundante. Se han contabilizado de 100 a 200 mil flores en un árbol adulto. Sin embargo, menos del 1 a 2% de estas flores llegará a fruta madura (Erickson y Brannaman, 1960). En la Fig. 4 se observa cómo cerca del 40% de las ramas evaluadas presentaron flores en la primera semana de marzo, mientras que el porcentaje de ramas con estructuras reproductivas decrece en las lecturas siguientes, evidenciando la disminución de estructuras reproductivas en el transcurso del tiempo.
El fruto de los cítricos presenta tres periodos bien diferenciados, conocidos como fases. La fase I, o periodo de crecimiento exponencial, va desde el final de la antesis hasta la caída fisiológica de los frutos. A continuación se inicia la fase II, que se caracteriza por la marcada expansión de los tejidos, y por la elongación celular. Finalmente, en la fase III, que comprende todos los cambios relacionados con la maduración, el fruto presenta una reducida tasa de crecimiento (Agustí, 2003). En el caso que nos ocupa, la duración de la fase I fue de 5 semanas a partir del final de la antesis (Orduz et al., 2009). Esto indica que la caída fisiológica se presentó alrededor de la tercera semana de mayo, situación que no se evidencia en las lecturas realizadas, ya que estas muestran un aumento en la proporción de ramas con frutos en los meses de mayo y junio. Esto implica que la metodología utilizada en el presente trabajo tiene limitaciones para el estudio de procesos puntuales, y debe ser complementada con otros procedimientos.
Conclusiones
Las plantas de mandarina ‘Arrayana’ analizadas en el presente estudio presentaron tres picos de brotación anual, correspondientes a marzo-abril, julio-agosto y septiembre-octubre. Este comportamiento se asemeja al obtenido en regiones subtropicales, y está relacionado con el comportamiento monomodal de la precipitación en el piedemonte del Meta.
La principal floración del cultivo acompañó la brotación de marzo (dos a tres semanas después del inicio de las lluvias). Este comportamiento confirma al estrés hídrico como el principal factor de inducción floral en condiciones tropicales.
Además de la floración inicial, se observaron flores extemporáneas durante los meses de lluvia (en ausencia de estrés hídrico). Se desconoce qué factores endógenos inducen la floración en estas condiciones climáticas.
La metodología de Garran et al. (1993) presentó una buena adaptación para estudios fenológicos tropicales, ya que cuantifica las modificaciones que experimentan las plantas por influencia de las variables climáticas, en especial de la precipitación.
La metodología utilizada no es sensible a fenómenos puntuales, tales como la caída fisiológica. Por tal razón debe ser complementada con otras metodologías desarrolladas para estudios que tengan esas características.
Agradecimientos
Por la colaboración prestada en la realización de este trabajo, los autores agradecen a los señores Heberth Velásquez, Capitolino Ciprian, Alfredo Pardo y Ricardo Quevedo; de la misma forma, al personal directivo y administrativo y a los compañeros investigadores del C.I. La Libertad de Corpoica.
Literatura citada
Agustí, M. 2003. Citricultura. Ediciones Mundi-Prensa, Madrid. [ Links ]
Azkue, M. 2009. La fenología como herramienta en la agroclimatología. En: www.infoagro.com/frutas/fenologia.htm consulta: junio de 2009. [ Links ]
Cassin, J., J. Bourdeaut, J. Fougue, V. Furan, J.P. Galliard, J. Le- Bourdelles, G. Montagut y C. Moreuil. 1969. The influence of climate upon the blooming of citrus in tropical areas. pp. 315-323. En: Proc. First Intl. Citrus Symp., Riverside, CA. [ Links ]
Davenport, T.L. 1990. Citrus flowering. Hort. Rev. 12, 349-408. [ Links ]
Davies, F.S. 1997. An overview of climatic effects on citrus flowering and fruit quality in various parts of the world. pp. 1-4. En: Proc. Citrus Flowering and Fruit Short Course. IFAS, Citrus Research and Education Center, University of Florida, Lake Alfred, FL. [ Links ]
Davies, F.S. y L.G. Albrigo. 1994. Citrus. CAB International, Wallingford, UK. [ Links ]
Erickson, L.C. y B.L. Brannaman. 1960. Abscission of reproductive structures and leaves of orange trees. Proc. Amer. Soc. Hort. Sci. 75, 222-229. [ Links ]
Erner, Y. y B. Bravdo. 1983. The importance of inflorescence leaves in fruit setting of Shamouti orange. Acta Hort. 139, 109-113. [ Links ]
FAO. 2006. Evapotranspiración del cultivo. Guías para la determinación de los requerimientos de agua de los cultivos. Bol. No. 56. Roma. [ Links ]
Garrán, S.M., M. Ragone y D. Vázquez. 1993. Observaciones fenológicas en plantas cítricas. p. 171. En: Resúmenes XVI Congreso Sociedad Argentina de Horticultura, Corrientes, Argentina. [ Links ]
Gil, G.F. 2000. Fruticultura: la producción de fruta. Ediciones Universidad Católica de Chile, Santiago. [ Links ]
Guardiola, J.L. 1997. Overview of flower bud induction, flowering and fruit set. pp. 5-21. En: Proc. Citrus Flowering and Fruit Short Course. IFAS, Citrus Research and Education Center, University of Florida, Lake Alfred, FL. [ Links ]
Jackson, L.K. 1999. Citrus cultivation. pp. 17-27. En: Timmer, L.W. y L.W. Duncan (eds.). Citrus health management. University of Florida, Gainesville, FL. [ Links ]
Koshita, Y. y T. Takahara. 2004. Effect of water stress on flower-bud formation and plant hormone content of Satsuma mandarin (Citrus unshiu Marc.). Scientia Hort. 99, 301-307. [ Links ]
Monselise, S.P. 1985. Citrus and related species. pp. 275-294. En: Halevy, A.H. (ed.). Handbook of fruit set and flowering. Vol. II. CRC Press, Boca Raton, FL. [ Links ]
Mendel, K. 1969. The influence of temperature and light on the vegetative development of citrus trees. pp. 259-265. En: Proc. First Intl. Citrus Symp. Riverside, CA. [ Links ]
Orduz, J. 2007. Estudios ecofisiológicos y caracterización morfológica y molecular de la mandarina Arrayana (Citrus reticulata Blanco) en el piedemonte llanero de Colombia. Tesis de doctorado. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. [ Links ]
Orduz, J. y J. Baquero. 2003. Aspectos básicos para el cultivo de los cítricos en el piedemonte llanero. Rev. Achagua 7(9), 7-19. [ Links ]
Orduz, J.O., A. Chacón y V.M. Linares. 2007. Evaluación del potencial de rendimiento de tres especies y un híbrido de cítricos en la región del Ariari del departamento del Meta (Colombia) durante 12 años, 1991-2003. Orinoquia 11(2), 41- 48. [ Links ]
Orduz, J. y G. Fischer. 2007. Balance hídrico e influencia del estrés hídrico en la inducción y desarrollo floral de la mandarina Arrayana en el piedemonte llanero de Colombia. Agron. Colomb. 25(2) 255-263. [ Links ]
Orduz, J.O., H. Monroy, L. Arango y G. Fischer. 2006. Comportamiento de la mandarina Arrayana en seis patrones en suelos ácidos del piedemonte llanero de Colombia. Agron. Colomb. 24(2), 266-273. [ Links ]
Orduz-Rodríguez, J.O., H. Monroy, G. Fischer y A. Herrera. 2009. Crecimiento y desarrollo del fruto de mandarina (Citrus reticulata) Arrayana en condiciones del piedemonte del Meta, Colombia. Rev. Colomb. Cienc. Hort. 3(2), 149-160. [ Links ]
Prado, A.K.S. 2006. Flowering, fruit set and carbohydrate levels in Valencia orange trees under different crop load status and with and without irrigation. Tesis de maestría, Mestrado em Agricultura Tropical e Subtropical, Instituto Agronômico, Campiñas, Brasil. [ Links ]
Reuther, W. 1973. Climate and citrus behavior. pp. 281-337. En: Reuther, W., L.D. Batchelor y H.J. Webber ( Eds.). Citrus industry. Vol. 3. University of California, Davis, CA. [ Links ]
Spiegel-Roy, P. y E.E. Goldschmidt. 1996. Biology of citrus. Cambridge University Press, Cambridge, UK. [ Links ]
Stover, E., B. Boman y L. Parsons. 2002. Physiological response to irrigation and water stress. pp. 112-116. En: Boman, B.J. (Ed.). Citrus and water stress. Institute of Agricultural Sciences, University of Florida, Gainesville, FL. [ Links ]
Tadeo, F., J.L. Moya, D. Iglesias, M. Talón y E. Primo-Millo. 2003. Histología y citología de cítricos. Generalitat Valenciana, Valencia, España. [ Links ]
Volpe, C.A. 1992. Citrus phenology. pp. 103-122. En: Proc. Second Intl. Sem. Citrus Physiol. Bebedouro, São Paulo, Brasil. [ Links ]

