










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.28 no.2 Bogotá May/Aug. 2010
Physicochemical characterization of 'Pinot Noir' grapevine (Vitis vinifera L.) fruit during its growth and development under high altitude tropical conditions
Caracterización físico-química del fruto de la uva (Vitis vinifera L.) 'Pinot Noir' durante su crecimiento y desarrollo en condiciones de trópico alto
Pedro José Almanza M.1, 5, Marco Antonio Quijano-Rico2, Gerhard Fischer3, Bernardo Chaves C.3, and Helber Enrique Balaguera-López4
1 Grupo de Ecofisiología Vegetal, Faculty of Agricultural Science, Universidad Pedagógica y Tecnológica de Colombia, Tunja (Colombia).
2 'Loma de Puntalarga' Vineyard and Winery, Nobsa (Colombia).
3 Department of Agronomy, Faculty of Agronomy, Universidad Nacional de Colombia, Bogota (Colombia).
4 Grupo de Investigaciones Agrícolas (GIA), Faculty of Agricultural Science, Universidad Pedagógica y Tecnológica de Colombia, Tunja (Colombia).
5 Corresponding author. ppcalma@gmail.com
Received for publication: 29 January, 2009. Accepted for publication: 28 July, 2010
ABSTRACT
Although native of temperate climates, grapevines are grown in Colombia in the department of Boyaca, between 2,200 and 2,560 m a.s.l. Under these specific conditions, both the physicochemical behavior of the fruit and its optimum harvest time as measured in growing degree days (GDD) had remained unknown so far. On these grounds, in the municipality of Nobsa (Boyaca, Colombia), grapevines of the variety Pinot Noir were physico-chemically characterized during their growth and development. Since day 21 after anthesis (DAA) until overripening, the physicochemical characteristics of 20 berries from each of three clusters we collected every week were carefully assessed. A total of 826.2 GDD accumulated during fruit growth and development, thus completing 126 DAA. The accumulation of both fresh and dry mass followed double sigmoid curves. During early growth there was a decrease in pH, total soluble solids (TSS) and technological maturity index (TMI=TSS/TTA), coupled to an increase in total titratable acidity (TTA). After this period and until harvest, pH, TSS and TMI increased while TTA declined. Based on fresh mass, SST and TMI it was possible to set optimum harvest time at 800.6 GDD.
Key words: tropical viniculture, temperature accumulation, phenological stages.
RESUMEN
A pesar de ser originaria de climas templados, la uva para vino se cultiva en Colombia en el departamento de Boyacá, entre los 2.200 y 2.560 msnm, condiciones en las cuales el comportamiento fisicoquímico del fruto, así como el momento óptimo de su cosecha en función de la acumulación de grados día de crecimiento (GDC), eran hasta ahora desconocidos. Por ello, a través de la presente investigación –realizada en el municipio de Nobsa (Boyacá, Colombia)– se llevó a cabo la caracterización fisicoquímica del fruto de la variedad Pinot Noir durante su crecimiento y desarrollo. Desde el día 21 después de la antesis (DDA) hasta la sobremaduración, se recolectaron semanalmente tres racimos, de cada uno de los cuales se tomaron 20 bayas para la evaluación de sus parámetros físico-químicos. Durante el crecimiento y desarrollo del fruto se acumularon 826,2 GDC (126 DDA). Tanto la acumulación de masa seca como la de masa fresca siguieron una curva doble sigmoide. Durante el inicio del crecimiento se observó una disminución de pH, sólidos solubles totales (SST) e índice de madurez tecnológica (IMT=SST/ATT), acompañada de un aumento en la acidez total titulable (ATT). Pasado este momento, y hasta la cosecha, se observó un aumento de pH, SST e IMT, mientras que la ATT disminuyó. Con base en los registros de masa fresca, SST e IMT, fue posible establecer el punto óptimo de cosecha en 800,6 GDC.
Palabras clave: viticultura tropical, acumulación de temperatura, estados fenológicos.
Introduction
Originated in the temperate zone of western Asia, where it produces one yearly crop, the grapevine (Vitis vinifera L.) is grown in the five continents, among which the largest cultivated area (4,900,000 ha) corresponds to Europe. In America, countries such as Chile, Argentina, Brazil, Peru and Uruguay stand out both for their production records and planted areas (OIV, 2005). In Colombia, under cold tropical climate conditions, the main wine grape producers are located in the department of Boyaca, between 2,200 and 2,560 m a.s.l., where they have been obtaining two yearly crops during the last 24 years due to local agroclimatic conditions and adequate crop management (Quijano, 2004).
Good wines depend on good grape quality, which in turn results from a series of factors such as cultivar, environment, climate and crop management practices, all of which interact for the necessary sugars and aromatic compounds to reach optimum levels during fruit ripening. A wine's main chemical features, which determine its sensorial attributes, result from the complex interaction between territory, variety and man, as summarized by the French term 'terroir' (Quijano, 2006).
The choice of grapevine varieties for a specific region, insofar as it implies assessing the genotype-environment dynamics, requires investigating the crop's phenological stage timing and behavior, which are in turn related to regional climate (Mullins et al., 1992; Chavarría et al., 2009).
In this respect, Jones and Davis (2000) and Tesic et al. (2002) assert that phenological stage span varies with climate, cultivar, and geographical location. Such analysis is also important for interpreting the interaction of the crop with the local microclimate (Terra et al., 1998). In addition, the physicochemical and thermal characterization of the grapevine during fruit growth and development is necessary to program crop management practices such as fertilization, irrigation, phytosanitary control and harvest (Mullins et al., 1992).
Three stages can be observed during grape growth and ripening:
During the herbaceous period the grape starts its evolution featured by its hard consistency, green color and high photosynthetic capacity. This stage goes from fruit set to the beginning of veraison, and is characterized by rapid fruit growth resulting from active cell division (Reynier, 1995; Agustí, 2000; Hidalgo, 2002). Taking place at the end of this phase, veraison consists in a brief growth delay accompanied by a color change in the fruit's epidermis. In the case of variety 'Pinot Noir', said change goes from green to light red due to chlorophyll loss and anthocyanin synthesis. During this stage, the beginning of pectin synthesis resulting from the development of the seed competes with that of the fruit, which, nevertheless, finally attains physiological maturity (Ribéreau-Gayon et al., 1998; Winkler et al., 1974).
During the ripening period the grape changes its color, resumes growth and behaves as a transformation and mostly a storage organ. A swift physicochemical evolution characterizes the fruit during this stage (Salazar and Melgarejo, 2005).
Along the overripening period the grape loses water due to transpiration. No sugar accumulation is registered, but an acidity decline due to low supply from the source, and possibly to a respiration increase. According to Reynier (1995), low acidity determines low wine quality; and De Rosa (1998) mentions that great wines depend on overripening.
Grape development stage assessment can be done through evaluating physicochemical parameters such as fresh and dry mass, pH, total acidity and soluble solid content (Salisbury and Ross, 1994). In the case of grapes all these parameters follow double sigmoid curves (Agustí, 2000; Winkler et al., 1974).
The presence of grape quality indicating compounds is assessed through bioclimatic indexes, among which physiological time, expressed in units known as Growing Degree Days (GDD), indicates the accumulated heat that, affecting the development rate of many organisms, is required by them to shift to a further stage in their life cycle (Rodríguez and Flórez, 2006). Stenzel et al. (2006) assert that thermal sum, also defined as accumulated GDD, is one of the most frequently used methods to ponder the relation between plant development and temperature. It is defined as the sum of the daily temperature records scoring above a specific minimum base level and below a maximum threshold one. This specific heat accumulation is needed by the plant to complete either its whole life cycle or a specific phenological stage (Souza, 1990). According to Champagnol (1984) and Hidalgo (2002), grapevine progressive budding is promoted by temperatures above 10°C, which is thus considered its base temperature.
In warm tropical zones the temperature remains steady within a 20 to 30°C range, which stimulates faster phenological phenomena than temperate and subtropical climate temperatures (Valor and Sánchez, 2003). High Andean valleys allow the successful growing of an ample variety of fruit crops including V. vinifera, to which the genotype –climate interaction facilitates bearing products with favorable physicochemical features. Thus, the present study focused on the grapes of the variety 'Pinot Noir', which is abundantly grown in the 'Loma de Puntalarga' region (Boyaca, Colombia) to prepare high quality wines (Quijano, 2001). The physicochemical behavior of said fruits along their growth and development as a function of GDD accumulation under the high altitude tropical conditions of the studied site was unknown until the present study, and thus was defined as its central objective.
Materials and methods
The work was conducted in 'Loma de Puntalarga' vineyard, in the municipality of Nobsa, Boyaca (Colombia), located at 5°46'47.1" N; 72°58'36.5" W; 2,560 m a.s.l. During the research, an average temperature of 16.5°C was recorded for the region, as well as a bimodal precipitation regime of 830 mm year-1, with peaks in April-May and October-November. It is covered with sandy loam, light-textured soils which are low in fertility and cemented by iron oxides (Quijano, 2006). Cloudiness is scarce, and irradiation reaches 476 cal cm-2 along 6.5 h d-1. Sprinkling irrigation is applied according to phenology and crop needs. Trained to a simple Guyot system with a three wire trellis, the 23 year old plants were cultivated at a 1.2 x 2.0 m distance. Fifty one inflorescences were chosen to be treated according to protocol established by Bautista and Vargas (1981), making sure they had all reached anthesis. Three bunches were randomly chosen every week since day 21 after anthesis (DAA) until fruit development was completed. Twenty grapes were obtained from each bunch (10 for chemical analysis, and 10 for physical testing), for a total of 1,020 evaluated fruits, according to methodology specified by Bordeu and Scarpa (1998).
A Tecpel data logger 322 was placed inside the vineyard to register the temperature records involved in GDD accumulation; which was calculated through equation 1, according to methodology by Rodríguez and Flórez (2006).
GDC = ((Tmax + Tmin)/2) - Tbase, (1)
where Tmax and Tmin are, respectively, maximum and minimum daily air temperatures, and Tbase is the temperature at which the metabolic process of the grape is at its lowest point, which corresponds to 10°C (Reynier, 1995; Champagnol, 1984; Hidalgo, 2002; Winkler et al., 1974).
The evaluated fruit response variables and their respective determination methods were: fresh mass as measured directly with a 0.01 g precision scale; dry mass, as obtained after dehydration in a muffle furnace at 75°C for 48 h; pH, determined with a potentiometer; total soluble solids (TSS), with an Atago manual refractometer, for them to be expressed in Brix degrees; total titratable acidity (TTA), through titration with 0.1 N NaOH until reaching pH 8.2, and expressed as tartaric acid (AOAC, 1990); technical maturity index, calculated as (TMI = TSS/TTA); and finally absolute and relative growth rates (AGR and RGR, respectively), according to the methodology of Carranza et al. (2009).
The obtained data were analyzed by descriptive statistics in a SAS® v. 8e. solver (Cary, NC), which determined mean and standard error values. Statistical models were used to adjust the different variables for GDD.
Results and discussion
Physical characteristics
Fresh and dry mass were observed to increase following a double sigmoid curve. Such behavior is explained here by means of two logistic growth models in each case. The first model explains growth from 195.8 to 565.8 GDD, which is when veraison takes place. The second model reproduces growth from 565.8 to 826.2 GDD regarding dry mass, and to 800.6 GDD with regards to fresh mass. After this moment, the fruits dehydrated and lost fresh mass considerably, which is the reason why such data were not taken into account to adjust the model (Fig. 1A and 4). The double sigmoid growth curve found here for variety 'Pinot Noir' was also reported for grapes by Almanza-Merchán and Balaguera-López (2009), as well as by Opara (2000), and is featured by two periods of fast growth separated by a slow or nil growth one (Opara, 2000).
During the herbaceous period, growth has been found to be more the result of cell division than of cell elongation (Hidalgo, 2002; Salazar and Melgarejo, 2005). According to our results, a total of 486.2 GDD accumulated during this stage, which went until 63 DAA. Fresh and dry mass increments were slow. Thus, at 195.8 GDD (21 DAA), these parameters exhibited respective values of 0.048±0.00057 g and 0.0056±0.00088 g; which, by the end of the period, had shifted to 0.85±0.041 g and 0.064±0.0034 g (Fig. 1A and 4).
During veraison, which went from 486.2 to 565.8 GDD (Fig. 1 and Tab.1), the fruit accumulated 79.6 GDD (14 d). Both fresh and dry mass were observed to increase slowly, even reaching a slight interruption, as reported by Salazar and Melgarejo (2005) and by Ribéreau-Gayon et al. (1998). This period was featured by a fruit color change from green through light red to wine red.
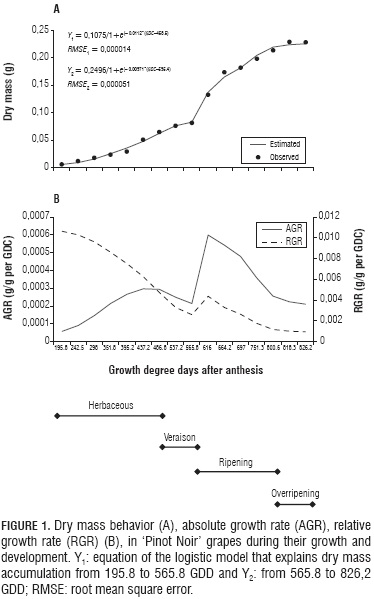
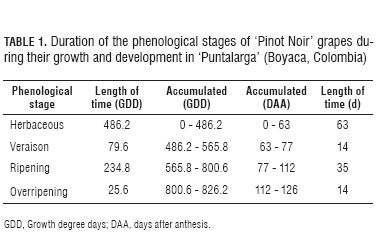
The ripening stage started at 565.8 GDD (77 DAA), and finished at 800.6 GDD (112 DAA) (Fig. 1), after accumulating 234.8 GDD along a period of 35 d (Tab. 1). According to Hidalgo (2002), it is featured by rapid mass increase, concomitant cell growth and biochemical component evolution.
Accompanying the practice observed among vine growers to obtain higher prices, the present assessment kept their 14 day overripening period, which accumulated a total of 25.6 GDD. A slight fruit dry mass increase (Fig. 1A) was accompanied by a significant fresh mass reduction (Fig. 4) due to water loss resulting from transpiration. This is in agreement with a report by During et al. (1987), who pointed out that the fruits undergo a fresh mass loss since day 95 after flowering.
Absolute growth rate (AGR), which expresses fruit size increment over time (Opara, 2000), was found to increase continuously since fruit set until 458.5 GDD (0.0003 g/GDD).
Then, by 565.8 GDD, it had dropped rapidly to 0.00021398 g/GDD, just to increase again and reach its peak (0.00060528 g/GDD) at 595.4 GDD, which was followed by a new drastic reduction that went on until harvest (Fig. 1B). These peaks define the double sigmoid curve that featured dry mass behavior, which contrasts with that of fruit crops that follow a simple sigmoid curve, whose AGR is characterized by showing only one peak found approximately at the middle of the growth process (Opara, 2000).
The first AGR peak (first sigmoid phase) corresponded to a considerable dry mass accumulation during the herbaceous period; and the second one (second sigmoid phase) is explained by the highest dry mass increase of the whole fruit growth. The first AGR valley corresponds to veraison, and the final one to the ripening - overripening transition. Both moments were featured by low dry mass increments (Fig. 1). Regarding this process, DeJong and Goudriaan (1989) assert that the first peak is determined by cell division and differentiation, and the second one by cell elongation and maturation. Similar AGR results have been found for Vaccinium corymbosum L., a fruit that also exhibits double sigmoid curve behavior (Godoy et al., 2008).
According to Carranza et al. (2009), relative growth rate (RGR) expresses the plant's dry mass increase (in the present case applied restrictively to the fruit) along a given time interval, taking as a reference the initial value of accumulated dry mass. For this reason, RGR tends to decrease as growth proceeds. In the present case, it reached a 0.0106 g/g per GDD peak at 195.8 GDD, and decreased drastically until the end of veraison (0.0026 g/g per GDD). At the beginning of ripening it went through a slight increase, which lasted until 595.4 GDD, and coincided with the highest AGR score. Then it dropped again until the end of overripening, when it showed its lowest value (0.0009 g/g per GDD) (Fig. 1B). The behavior of AGR found in the current research coincides with those reported by Staud et al. (1986) for grapes of cv. Bacchus, and by Godoy et al. (2008) for fruits of V. corymbosum.
Chemical features
The observed pH behavior fits a quadratic model, featured in this case by a reduction that went from 2.69±0.0088 at 195.8 GDD to 2.13±0.028 at 437.2 GDD. During this stage, the fruit exhibits active respiration and great capacity to synthesize malic and tartaric acids (Salazar and Melgarejo, 2005). At the end of the fruit growth, pH went up to 3.58±0.066 as a result of the degradation of organic acids and larger sugar accumulation (Fig. 2B and 3A). Plane et al. (1980) mention that grape pH for wine preparation should be fixed between 3.1 and 3.7 because higher or lower values affect wine color. In the current research, such range was reached after having accumulated 751.3 GDD (105 DAA) (Fig. 2A). In studying this same variety, Almanza-Merchán and Balaguera-López (2009) reported that pH started to increase after veraison, which constitutes a similar behavior to the one found in the present work.
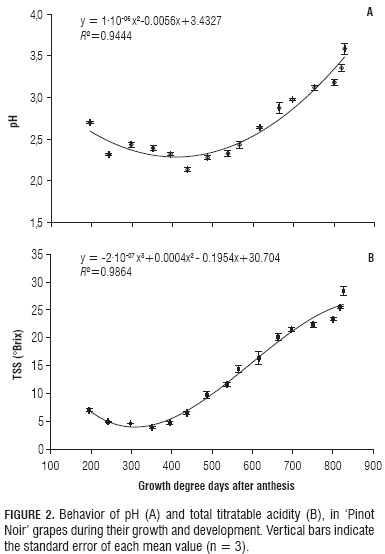
Cell pH is very important in regulating metabolism. In fruits, more than 90% of a cell's volume is occupied by the vacuole, which is usually very acid, thus exhibiting lower than 5 pH values (Nanos and Kader, 1993). In the case of grapes, values below 2 are prone to affect wine microbiological stability (Plane et al., 1980).
Also in extracting wine aromas, the most important factor is pH. For this reason, this parameter must be closely assessed during grape ripening. According to Vine (1997), the taste of unripe grapes between pH 3.2 and 3.3 accuses elevated total acidity. But, as higher levels promote the growth of undesirable bacteria, the limiting factor in preserving wine aromas is the ability of the winemaker.
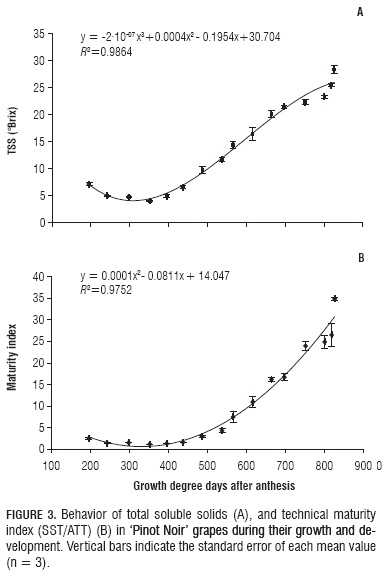
The observed TSS variation during grape growth and development fits a third grade polynomial featured by a slight decrease up to 351.8 GDD, followed by a considerable increase until harvest. However, with a record of 9.76±0.65 °Brix at 486.8 GDD, TSS increase was slow during the herbaceous period. A clear TSS gain started with veraison, at the end of which (664.2 GDD) this parameter reached 20.1±0.59°Brix. After that moment, accumulation was slower, until reaching harvest maturity with 23.3±0.24°Brix at 800.6 GDD. Anyway, during overripening there was another considerable TSS increment because the fruits, as it was already mentioned, experimented a fresh mass loss that increased TSS concentration up to 28.4±0.8°Brix (Fig. 3A). The TSS values found here are consistent with those reported by Almanza-Merchán and Balaguera-López (2009) and by Williams et al. (1985).
Wine quality can be improved by delaying grape harvest until the fruit reaches higher TSS levels and lower weight. Diminishing acidity can even lead to better acceptability, insofar as certain consumers prefer such lower levels. It should be highlighted that during the present work, the region where it was carried out presented elevated temperatures, which contributed to speeding up overripening.
TTA followed a cubic polynomial model. From 195.8 to 437.2 GDD, it was found to increase steeply until reaching a value of 4.068±0.0043%. This is explained by the fact that during this stage, which corresponds to the herbaceous period, organic acids such as malic and tartaric ones are actively synthesized (Reynier, 1995). After this point, TTA declined drastically up to 751.3 GDD, and then slowly until harvest (826.2 GDD), with respective values of 0.93±0.041% and 0.81±0.03% (Fig. 2B). Grapes are reported to present an acidity decrease not only due to the transformation of malic acid into other compounds, but also to respiration, to dilution resulting from water accumulation, and to migration of bases from the roots, which increases ash alkalinity (Reynier, 1995).
Bluske (2008) reports that in high altitudes, malic acid levels are relatively high as compared to those of tartaric acid, because the latter is consumed by respiration in larger amounts, thus misbalancing these acids in the resulting wine. This implies that malolactic fermentation must be handled carefully; otherwise, excessively light wines could be obtained. This same author states that red grape titratable acidity ranges between 7 and 10 g L-1 (0.7 to 1.0%) as expressed in tartaric acid, which is in agreement with the results of the present work, in which this parameter reached 0.94% at 803.8 GDD, 112 DAA (Fig. 2B).
A simple way to determine the optimum harvest moment is to complement the traditional methods applied by vine growers through monitoring grape fresh mass evolution. During the herbaceous stage, dry mass increment is relatively slow, and very slow during veraison, when growth takes place mainly in the seed. After this stage, fresh mass increases considerably until it starts to decline due to dehydration, which appears as a rise in Brix degrees (Fig. 4).
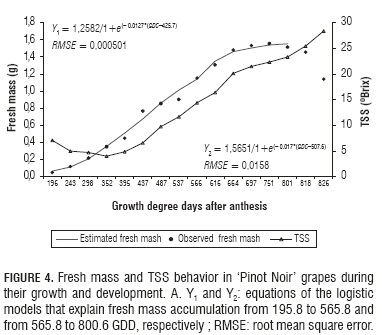
The largest fresh mass record (1.55 g) was attained 105 DAA, at 751.3 GDD. After this point, it started to decline due to transpirational fruit dehydration (Coombe and Mc Carthy, 2000). The fresh mass curve crossed the TSS one between 800.6 and 818.3 GDD (Fig. 4). This could be a good harvest moment as far as a considerable fresh mass accumulation coincides with the recommended TSS level (23 to 24°Brix) for quality wine grape (Reynier, 1995).
Several criteria are applied in determining grapevine harvest. Among them, technological maturity index (TMI) corresponds to the most commonly used ones.
In our results, this parameter's behavior shows a quadratic curve that goes down from 195.8 to 351.8 GDD, coinciding with an acidity increase during this period. Since then (351.8 GDD), a considerable, steady increment was observed until reaching 826.2 GDD, when a value of 34.88±0.34 (Fig. 3B) was observed as a result of the degradation of organic acids and TSS increase.
High TSS and low TTA levels are likely to determine microbiological stability problems, which in turn might affect wine color. According to Paoletti (2003), sensorial assessment is critical to confirm analytical evaluation data.
The observed TMI behavior is in agreement with that reported by Almanza-Merchán and Balaguera-López (2009), who hold that in the agroecological conditions of 'Loma de Puntalarga' vineyard, the Pinot Noir variety can be harvested at 126 DAA, when TMI is 21.43, TSS 24.4°Brix, TTA 1.14% and pH 3.33, corresponding to the beginning of overripening. Notwithstanding, in the present study the moment marked by these parameter values corresponded to the end of such period. This can be explained by the fact that the environmental conditions, especially temperature, were different in the two experiments. This manifests the importance of a more objective measure such as GDD. In this regard, and corroborating what we mentioned above about the relationship between fresh mass and TSS, the present research study indicates that this cultivar can be harvested after it has reached 800.6 GDD, when TMI is 24.87±1.47, TSS = 23.33 ± 0.24°Brix, TTA = 0.94±0.06%, pH = 3.18±0.03, dry mass = 0.21±0.0057 g and fresh mass = 1.51±0.01 g.
Conclusions
Fresh and dry mass behavior of the studied 'Pinot Noir' grapes followed a double sigmoid curve with two fast growing periods separated by a slow one that corresponded to veraison. Such behavior was explained by means of two subsequent logistic models.
During the beginning of the herbaceous period, the studied fruits revealed reductions in pH, TSS and TMI coupled to an increase in TTA. After this moment and until harvest, the opposite behavior was observed, that is, pH, TSS and TMI increased while TTA diminished.
The variety Pinot Noir can be harvested at the beginning of the overripening period (800.6 GDD), which guarantees sufficient fresh mass and TSS accumulation to favor both the farmer and the wine maker. At this point, fruit parameters were: TMI = 24.87±1.47; TSS = 23.33±0.24°Brix; TTA = 0.94±0.06%; pH = 3.18±0.03; dry mass = 0.21±0.0057 g and fresh mass = 1.51±0.01 g.
Literature cited
Agustí, M. 2000. Crecimiento y maduración del fruto. pp. 419-433. In: Azcon-Bieto, J. and M. Talón (eds.). Fundamentos de fisiología vegetal. McGraw-Hill Interamericana, Madrid. [ Links ]
Almanza-Merchán, P. and H.E. Balaguera-López. 2009. Determinación de los estadios fenológicos del fruto de Vitis vinifera L. bajo condiciones del altiplano tropical en Boyacá. Rev. UDCA Actual. Divulg. Cient. 12(1), 141-150. [ Links ]
AOAC. 1990. Official methods of analysis. 15th ed. Association of Official Analytical Chemists, Arlington, VA. [ Links ]
Bordeu, E. and J. Scarpa. 1998. Análisis químico del vino. Impresos Universitaria S.A., Santiago. [ Links ]
Bluske, I. 2008. Los vinos de altura. In: http://vinosdealtura.es/img/pdf/producción.pdf; consulted: January, 2010. [ Links ]
Bautista, D. and G. Vargas. 1981. Estudio del ciclo y determinación de los requerimientos heliotérmicos de algunas variedades de vid en condiciones tropicales. Agron. Trop. 31(1-6), 1-13. [ Links ]
Carranza, C., O. Lanchero, D. Miranda, and B. Chaves. 2009. Análisis del crecimiento de lechuga (Lactuca sativa L.) 'Batavia' cultivada en un suelo salino de la Sabana de Bogotá. Agron. Colomb. 27(1), 41-48. [ Links ]
Champagnol, F. 1984. Elements de physiologie de la vigne et de viticulture générale. Imprimerie Déhan, Montpellier, Francia. [ Links ]
Chavarría, G., H.P. Dos Santos, F. Mandelli, G.A. Bettio, H. Bergamaschi, and L.S. Cardoso. 2009. Caracterização fenológica e requerimento térmico da cultivar moscato giallo sob cobertura plástica. Rev. Bras. Frutic. 31(1), 119-126. [ Links ]
Coombe, B. and M. McCarthy. 2000. Dynamics of grape berry growth and physiology of ripening. Aust. J. Grape Wine Res. 6, 131-135. [ Links ]
De Rosa, T. 1998. Tecnología de los vinos blancos. Ediciones Mundi-Prensa, Madrid. [ Links ]
DeJong, T.M. and J. Goudriaan. 1989. Modeling peach fruit growth and carbohydrate requirements: reevaluation of the double-sigmoid growth pattern. J. Amer. Soc. Hort. Sci. 114(5), 800-804. [ Links ]
During, H., A. Lang, and F. Oggionni. 1987. Patterns of water flow in Riesling berries in relation to developmental changes in their xylem morphology. Vitis 26, 123-131. [ Links ]
Godoy, C., G. Monterubbianesi, and J. Tognetti. 2008. Analysis of highbush blueberry (Vaccinium corymbosum L.) fruit growth with exponential mixed models. Scientia Hort. 115(4), 368-376. [ Links ]
Hidalgo, L. 2002. Tratado de viticultura general. Ediciones Mundi-Prensa, Madrid. [ Links ]
Jones, G.R. and E. Davis. 2000. Climate influences on grapevine phenology, grape composition, and wine production and quality for Bordeaux, France. Amer. J. Enol. Vitic. 51(3), 249-261. [ Links ]
Mullins, M.G., A. Bouquet, and L.E. Williams. 1992. Biology of the grapevine. Cambridge University Press, New York, NY. [ Links ]
Nanos, G.D. and A.A. Kader. 1993. Low O2-induced changes in pH and energy charge in pear fruit tissue. Postharv. Biol. Technol. 3, 285-291. [ Links ]
Opara, L.U. 2000. Fruit growth measurement and analysis. Hort. Rev. 24, 373-431. [ Links ]
Organización Internacional de la Viña y el Vino (OIV). 2005. Situación del sector vitivinícola mundial en 2005. In: http://news.reseau-concept.net/images/oiv_es/client/Commentaire_Statistiques_2005_ES.pdf; consulted: March, 2010. [ Links ]
Paoletti, A. 2003. Indici di maturazione fenolica de glories e terroir. pp. 502-503. In: Fregone, M., D. Schuster, and A. Paoletti (eds.). Terroir, zonazio ne viticultura. Phytoline, Affi-Caprino, Italia. [ Links ]
Plane, A.R., R.L. Mallick, and L.D. Wiers. 1980. An acidity index for taste of wines. Amer. J. Enol. Vitc. 31(3), 265-268. [ Links ]
Quijano, M. 2001. Los vinos del valle del sol. Cultura Científica 1, 5-11. [ Links ]
Quijano, M. 2004. Ecología de una conexión solar. De la adoración del sol al desarrollo vitivinícola regional. Cultura Científica 2, 5-9. [ Links ]
Quijano, M. 2006. Investigación e innovación. Promoción y defensa del "terroir" regional. Cultura Científica 4, 35-41. [ Links ]
Reynier, A. 1995. Manual de viticultura. 5th ed. Ediciones Mundi-Prensa, Madrid. [ Links ]
Ribéreau-Gayon, P., B. Donèche, D. Dubordieu, and A. Lonvaud. 1998. Traité d'oelignologie. Vol. 1. Microbiologie du vin: Vinifications. Dunod, Paris. [ Links ]
Rodríguez, W. and V. Flórez. 2006. Comportamiento fenológico de tres variedades de rosas rojas en función de la acumulación de la temperatura. Agron. Colomb. 24(2), 247-257. [ Links ]
Salazar, D.M. and P. Melgarejo. 2005. Viticultura. Técnicas del cultivo de la vid, calidad de la uva y atributos de los vinos. Ediciones Mundi-Prensa, Madrid. [ Links ]
Salisbury, F.B. and C.W. Ross. 1994. Fisiología vegetal. Grupo Editorial Iberoamérica, México DF. [ Links ]
Souza, P.R. 1990. Algunos aspectos da influência do clima e temperatura sobre a cultura do arroz irrigado no sul de Brasil. Lavoura Arrozeira 43, 9-11. [ Links ]
Stenzel, N.M.C., C.S.V.J. Neves, C.J. Marur, M.B.d.S. Scholz, and J.C. Gomes. 2006. Maturation curves and degree-day accumulation for fruits of 'Folha Murcha' orange trees. Sci. Agric. 63(3), 219-225. [ Links ]
Terra, M.M., E.J.P. Pires, and C.V. Pommer. 1998. Tecnologia para a produção de uva Itália na região noroeste do Estado de São Paulo. 2nd ed. CATI, Campinas, Brazil. [ Links ]
Tesic, T., D.J. Woolley, E.W. Hewett, and D.J. Martin. 2002. Environmental effects on cv. Cabernet Sauvignon (Vitis vinifera L.) grown in Hawke´s Bay, New Zealand. I. Phenology and characterization of viticultural environments. Aust. J. Grape Wine Res. 8(1), 15-26. [ Links ]
Valor, O. and J. Sánchez. 2003. Brotación, fertilidad de brotes laterales y ubicación del racimo en el cultivar de vid tucupita en condiciones tropicales. Bioagro 15(3), 201-208. [ Links ]
Vine, R. 1997. Winemaking. White wine. Chapman and Hall, New York, NY. [ Links ]
Williams, D.W., H.L. Andris, R.H. Beede, D.A. Luvisi, M.V.K. Norton, and L.E. Williams. 1985. Validation of a model for the growth and development of the Thompson seedless grapevine. II. Phenology. Amer. J. Enol. Vitic. 36(4), 283-289. [ Links ]
Winkler, A., J. Cook, N. Kliewer, and A. Lider. 1974. General viticulture. University of California Press, Berkeley, CA. [ Links ]

