










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.28 no.2 Bogotá May/Aug. 2010
Aspectos nutricionales y metabolismo de Stevia rebaudiana (Bertoni). Una revisión
Nutritional aspects and metabolism of Stevia rebaudiana (Bertoni). A review
Alfredo de Jesús Jarma O.1, Enrique Miguel Combatt C.1 y José Alejandro Cleves L.2
1 Departamento de Agronomía, Facultad de Ciencias Agrícolas, Universidad de Córdoba, Montería (Colombia).
2 Facultad Seccional Duitama, Universidad Pedagógica y Tecnológica de Colombia (UPTC), Duitama (Colombia).
3 Autor de correspondencia. ajarma@sinu.unicordoba.edu.co
Fecha de recepción: 9 de abril de 2009. Aceptado para publicación: 28 de julio de 2010
RESUMEN
Estevia es una planta selvática subtropical del alto Paraná, nativa del Noroeste de la provincia de Misiones en el Paraguay. Posee un potente edulcorante que llega a ser hasta 300 veces más dulce que la sacarosa y no contiene calorías. Las moléculas responsables de esta característica son glucósidos de diterpeno que se encuentran en las hojas y, cuando menos en los estadios iniciales, son sintetizados a partir del mevalonato, a través de la misma ruta del ácido giberélico. Los estudios que abordan el tema de la nutrición mineral en estevia en el mundo son escasos y se circunscriben en gran parte, a las investigaciones adelantadas en el Brasil por Malavolta y sus colaboradores en 1997. En Colombia, los estudios científicos sobre el tema se limitan a algunos resultados encontrados por la Universidad de Córdoba y Corpoica en trabajos desarrollados con nitrógeno, fósforo y potasio. Por tal motivo, el presente trabajo revisa la ruta de síntesis de los principales edulcorantes producidos por la planta, discute el importante papel que en ella juegan algunos minerales, y realiza un análisis de los aspectos nutricionales más importantes de esta planta, con base en los estudios realizados en Brasil y Colombia.
Palabras clave: estevia, edulcorantes, nutrición mineral, metabolismo secundario.
ABSTRACT
Stevia is a subtropical wild plant of the upper Parana river, native of the northwest of the province of Missions in Paraguay. The plant contains a potent sweetener that is 300 times stronger than sucrose and supplies no calories. The molecules that determine this feature are diterpene glycosides, which are present in the leaves and, at least during the first stages, are synthesized from mevalonate, following the same path as gibberellic acid. Studies on mineral nutrition in stevia are scarce. They are circumscribed indeed to those conducted in Brazil by Malavolta and collaborators. In Colombia, the scientific studies about the topic are limited to some works by Universidad de Córdoba and Corpoica dealing with nitrogen, phosphorus and potassium. In this framework, the current work reviews the synthesis path of the main edulcorants produced by this plant, discusses the important role played by certain minerals in this process, and based on studies conducted in Brazil and Colombia, analyzes the most important nutritional aspects of this species.
Key words: stevia, edulcorant, mineral nutrition, secondary metabolism.
Introducción
Stevia rebaudiana es una planta herbácea perenne perteneciente a la familia de las Asteraceas, que crece como arbusto salvaje en el suroeste de Brasil y Paraguay, y generalmente se propaga comercialmente de manera asexual (Hearn y Subedi, 2008). Cobra un alto valor entre los vegetales nativos de estos países, debido a que contiene glucósidos de diterpeno bajos en calorías, llamados comúnmente esteviósidos, cuyo poder edulcorante en estado puro y cristalino puede ser 300 veces mayor que el del azúcar de caña. Otros compuestos químicos de interés también han sido aislados de especies pertenecientes al género Stevia, tales como longipinenos provenientes de S. pilosa (Álvarez-García et al., 2005),S. viscida (Román et al., 1995) y S. lucida (Guerra-Ramírez et al., 1998).
Los edulcorantes, en su mayoría concentrados en las hojas, son sintetizados, al menos en los estados iniciales, usando la misma ruta del ácido giberélico a partir del mevalonato (Hsieh y Goodman, 2005; Guevara-García et al., 2005; Kasahara et al., 2002). Varios autores han indicado que la diferencia radica que en estevia, el kaureno, precursor de dichas hormonas, se convierte en esteviol en el retículo endoplásmico (Geuns, 2003; Totté et al., 2000, 2003; Brandle et al., 2002). El edulcorante, cuyos dos glucósidos principales son el esteviósido (110-270 veces más dulce que el azúcar) y rebaudiósido A (180-400 veces más dulce que el azúcar), este último de mayor valor comercial (Liu, 2010), cuya diferencia radica solamente en la presencia de una glucosa (Erkucuk et al., 2009; Carakostas et al., 2008), es un polvo cristalino blanco (RIRDC, 2002; Brandle y Telmer, 2007); los científicos lo llaman una “molécula noble”, debido a que es 100% natural, no tiene calorías, las hojas se pueden utilizar en su estado natural y solo se requieren cantidades pequeñas. Otras ventajas adicionales son que no eleva los niveles de glucosa en la sangre, no aporta calorías al ser metabolizado, es antiácido, cardiotónico, no produce caries al no ser fermentado por las bacterias orales, y se distingue de los edulcorantes artificiales por no tener sabor metálico y no ser cancerígeno (Tadhani et al., 2007).
La seguridad y las bondades de los edulcorantes de estevia en el metabolismo animal se han demostrado (Jeppesen et al., 2002; Geuns et al., 2003; Koyama et al., 2003; Wong et al., 2004; Narissara et al., 2004), así como sus efectos positivos en la salud del hombre, en las que se reportan propiedades anti-rotavirus (Takahashi et al., 2001), mejoramiento de la hipertensión (Hsieh et al., 2003), tratamiento de la diabetes mellitus tipo 2, al estimular la secreción de insulina actuando sobre las células b del páncreas (Jeppesen et al., 2000; Reziwanggu et al., 2004; Gregersen et al., 2004).
Aunque existe consenso mundial de las ventajas de estevia para la salud humana (Prakash et al., 2008) y que su cultivo es un sistema altamente generador de empleos rurales (Jarma, 2008; Espitia et al., 2009), aún existen grandes vacíos en el conocimiento de esta especie, principalmente en aspectos como la nutrición y las variables ambientales (Das et al., 2007; Gardana et al., 2010; Espitia et al., 2008) y su relación con la síntesis de las principales moléculas edulcorantes (Jaitak et al., 2008). Por lo anterior, el presente artículo pretende revisar la ruta de síntesis de los glucósidos de esteviol, indicar el papel de algunos elementos minerales en la misma y hacer un análisis de los principales reportes en América sobre los requerimientos nutricionales de estevia, haciendo énfasis en los casos particulares de Brasil y Colombia.Síntesis de glucósidos de diterpeno
La ruta de síntesis que da origen a los principales edulcorantes de S. rebaudiana es la del ácido giberélico, en la que la cadena de terpenos se alarga a partir de uniones sucesivas de isopentenil pirofosfato (IPP), considerado como el isopreno activo (Richman et al., 1999; Buchanan et al., 2000; Azcón-Bieto y Talón, 2008). Las enzimas que catalizan el alargamiento de la cadena terpénica son las prenil transferasas, de las cuales se han aislado, hasta ahora, la geranil pirofosfato sintasa, enzima identificada en plastos que requieren Mg2+ o Mn2+ como cofactores (Suga y Endo, 1991; Kim et al., 1996); la farnesil pirofosfato sintasa, que requiere Mg2+ como cofactor, se localiza en el citoplasma y cataliza la formación de farnesil pirofosfato (FPP) en dos etapas; por último, la geranil geranil pirofosfato sintasa, que requiere Mn2+ para su máxima actividad, se localiza en plastos y forma el geranil geranil pirofosfato (GGPP) en tres etapas (Fig. 1). Brandle et al. (2002), quienes han clonado y secuenciado el gen de estevia que codifica para la enzima copalil pirofosfato sintasa, responsable de la conversión de GGPP a CPP, afirman que la hidroxilación del ácido ent-kaurenoico en la posición C13 es el punto de divergencia para la síntesis del esteviol y las giberelinas. Esta hidroxilación, que requiere NADPH y oxígeno molecular proveniente del estroma, es catalizada por la enzima ácido ent-kaurenoico 13-hidroxilasa (Jarma et al., 2010).
Elementos minerales y estevia
Macronutrientes
Nitrógeno
Este elemento es primordial para la planta, ya que forma parte de proteínas y otros compuestos orgánicos esenciales, como enzimas, coenzimas, vitaminas, ácidos nucleicos, clorofila, reguladores de crecimiento, nucleótidos, etc. La importancia de este nutriente, por tanto, es por demás obvia, al estar presente en la mayoría de las reacciones fisiológicas de la célula y de las plantas. Como el nitrógeno se encuentra presente en muchos compuestos esenciales, no sorprende en absoluto que el crecimiento sea lento ante su escasez (Lian et al., 2005 y 2006), aunque las eficiencias en el uso de este elemento difieren de manera significativa entre especies y dentro de estas (Fan et al., 2005). En estas condiciones, las plantas presentan una clorosis general, especialmente en las hojas más viejas.
En trabajos adelantados en nutrición de estevia, De Lima y Malavolta (1997) indican que una deficiencia de N disminuye el tamaño de las células y aumenta el grosor de sus paredes; la división y expansión celular también disminuyen, reduciendo el tamaño de todas las partes morfológicas de la planta, principalmente hojas y frutos. Así mismo, los autores encontraron una disminución en la ramificación como consecuencia de una deficiencia de N, y lo atribuyeron a una probable inhibición en el crecimiento de las yemas axilares.
Fósforo
El P forma parte esencial de muchos glucofosfatos como la uridin difosfato glucosa, UDP-glc, molécula donadora de glucosa en la síntesis de los glucósidos de diterpeno (Shibata et al., 1995), y de otros que participan en la fotosíntesis, la respiración y distintos procesos metabólicos. También forma parte de nucleótidos (como sucede en el ARN y el ADN) y de los fosfolípidos que se encuentran en las membranas. Además, desempeña un papel fundamental en el metabolismo energético, debido a su presencia en las moléculas de ATP, ADP, AMP y pirofosfato (PPi), y determina de manera notable la configuración espacial de la raíz (Liao et al., 2001; Dong et al., 2004; Zhao et al., 2004).
Potasio
El K+ es un activador de muchas enzimas esenciales para la fotosíntesis y la respiración, y también activa enzimas que son necesarias para formar almidón y proteínas. Es uno de los contribuyentes más importantes al potencial osmótico de las células y, por consiguiente, a su presión de turgencia, siendo fundamental en el balance de las cargas negativas de ácidos orgánicos y de aniones tales como el sulfato y nitrato absorbidos por las raíces del medio externo y manteniendo la electro-neutralidad de las células (Taíz y Zeiger, 2006). El mecanismo estomático depende del flujo de iones K+, por lo que plantas que crecen bajo deficiencias de este nutriente presentan daños en esta actividad.
Siendo el K+ activador de varias enzimas, su deficiencia ocasiona disturbios en eventos metabólicos, como por ejemplo, acumulación de compuestos nitrogenados libres o solubles. Estos compuestos pueden ser aminoácidos, amidas y amonio, productos de la descarboxilación de aminoácidos, como putrescina, N-carbamilputrescina y agmatina. De Lima y Malavolta (1997) encontraron menor proporción de ramas en estevia ante deficiencias de K. Una disminución en la proporción de ramas puede generar una menor migración de fotoasimilados a través de los vasos conductores, lo que probablemente esté relacionado con un proceso de fosforilación (producción de ATP), pues cuando el nivel de K es alto, puede ocurrir un estímulo en la producción de ATP.
Calcio
El calcio resulta esencial para las funciones normales de la membrana citoplasmática en cualquier célula, seguramente a modo de enlazador de fosfolípidos, tanto entre sí como a proteínas de membrana. Al parecer, el Ca2+ no puede cargarse en las células transportadoras del floema, y por esta razón los síntomas de deficiencia son más marcados en los tejidos jóvenes. Últimamente el calcio está recibiendo más atención porque en la actualidad se reconoce que todos los organismos mantienen concentraciones inesperadamente bajas de Ca2+ libre en el citosol, habitualmente menores del 4 mM (Marschner, 2002). Esto resulta cierto incluso cuando el calcio es muy abundante en muchas plantas.
La mayor parte del calcio que contienen las plantas se encuentra en las vacuolas centrales, y en las paredes celulares se encuentra unido a ciertos polisacáridos llamados pectatos. En las vacuolas, el calcio suele precipitarse en forma de cristales insolubles de oxalatos, y también, en algunas especies, en forma de carbonato, de fosfato o de sulfato insolubles. Parece ser que las concentraciones bajas, casi micromolares, de Ca2+ en el citosol deben mantenerse en parte para impedir la formación de sales de calcio insolubles, obtenidas partiendo del ATP y de otros fosfatos orgánicos. Además, las concentraciones de Ca2+ por encima del margen micromolar inhiben la corriente citoplásmica. Aunque se activan unas cuantas enzimas mediante Ca2+, muchas otras quedan inhibidas, lo que hace más necesario todavía que las células mantengan concentraciones muy bajas de Ca2+ en el citosol, donde existen muchas enzimas.
Una parte considerable del calcio existente en el citosol se une de forma irreversible a una pequeña proteína llamada calmodulina; esta unión modifica su estructura, activando varias enzimas. En la actualidad se está investigando intensamente la relación que tiene el calcio y la calmodulina con la actividad enzimática de las plantas. Es muy posible que el Ca2+ efectúe una función de activador enzimático, especialmente cuando el ion se encuentra unido a la calmodulina o a proteínas muy afines. Adicionalmente, es reconocida su importancia como segundo mensajero en las respuestas de las plantas a señales ambientales y hormonales (White y Broadley, 2003).
En estevia, Utumi et al. (1999), al inducir deficiencias de Ca2+, hallaron síntomas de necrosis apical en primordios foliares, que concluyó en muerte descendente, decrecimiento en el contenido de esteviósidos, las nuevas ramas formadas fueron quebradizas y las raíces presentaron reducción en su longitud y grosor.
Magnesio
Además de su presencia de clorofila, el magnesio resulta un elemento esencial porque se combina con el ATP, permitiéndole participar en muchas reacciones, como la fosforilación del AMV para formar IPP, primer compuesto isoprenoide en la síntesis de los glucósidos de esteviol. Seguidamente, en la misma ruta de síntesis, actúa como cofactor de la enzima citoplasmática farnesil pirofosfato sintasa, que cataliza la formación del FPP. Además, el Mg2+ activa otras enzimas necesarias en el proceso de la fotosíntesis, la respiración y la formación de ADN y de ARN (Taíz y Zeiger, 2006).
Azufre
En algunas especies vegetales, el S no se redistribuye con facilidad desde los tejidos maduros, así que la deficiencia se nota antes en las hojas más jóvenes, aunque en otras especies la mayor parte de las hojas se vuelven cloróticas casi al mismo tiempo o, a veces, hasta en las hojas más antiguas primero. Muchas plantas cultivadas, incluyendo la raíz, contienen aproximadamente una quinceava parte de azufre respecto a la cantidad de nitrógeno (en peso), lo que parece ser una clave interesante para evaluar necesidades nutritivas.
La mayor parte del azufre en las plantas se encuentra en las proteínas, más precisamente en los aminoácidos cisteína y metionina, que son constituyentes de las proteínas. Las vitaminas tiamina y biotina, así como la coenzima A (compuesto esencial para la respiración, la síntesis y la degradación de ácidos grasos, y del acetil CoA en la síntesis de glucósidos de diterpenos), son otros compuestos esenciales que contienen azufre. Por ello, no es extraño observar decrecimientos en el contenido de esteviósidos ante deficiencias de S, tal como lo reportaron Utumi et al. (1999), además de otros síntomas como clorosis y reducción en el número de hojas nuevas.
Micronutrientes
Boro
El rango de concentración de B en los tejidos de los vegetales es muy amplio, pudiendo variar de 5 a 300 mg kg-1 con base en la masa seca, y generalmente estos valores son superiores en dicotiledóneas respecto a monocotiledóneas (Epstein y Bloom, 2005). Los vegetales con deficiencia de boro pueden tener una amplia variedad de síntomas (Yan et al., 2006), dependiendo de la especie y la edad de la planta, aunque el primer síntoma suele ser la falta de crecimiento y el alargamiento anormal en las puntas de la raíz, junto con la inhibición de la síntesis de ADN y ARN. También se desactiva la división celular en el ápice del tallo y en las hojas más jóvenes. Du et al. (2002) indican que cultivares ineficientes en el uso del B registran una producción menor de carbohidratos y aminoácidos libres.
Aunque se ha publicado que el boro tiene una función esencial en la elongación de los tubos de polen, sus funciones bioquímicas son poco claras, en parte porque no se sabe cómo se modifica el B(OH)3 en las células, y en parte porque realmente puede cumplir muchas funciones. Quizá gran parte de este ácido débil se una, para formar complejos de borato cis-diol, con grupos hidroxilo contiguos que provienen de la manosa, y con otros azúcares de los polisacáridos de la pared celular (O'Neill y York, 2003), aunque no sucede con la fructuosa, la glucosa, la galactosa y la sacarosa, que no tienen distribuciones cis-diol de grupos hidroxilo. Aún no se ha identificado ninguna función específica con seguridad, pero existen evidencias que indican la participación especial del boro en la síntesis de los ácidos nucleicos, que es esencial para la división en los meristemos apicales; existe también consenso general en que hace parte de la estructura de la pared celular y de las sustancias pépticas asociadas a ella.
Hierro
El hierro acumulado en las hojas más antiguas se encuentra relativamente inmóvil en el floema, igual que en el suelo, probablemente porque se precipita internamente en las células de las hojas en forma de óxido insoluble o de compuestos de fosfato férrico, orgánicos o inorgánicos. Hay pocas evidencias directas de que se formen esos precipitados; quizás se puedan formar otros compuestos insolubles, que nos resultan desconocidos, pero son similares. Su transporte a larga distancia puede darse en forma oxidada en un complejo Fe-citrato (Hell y Stephan, 2003). En los cloroplastos se almacena una forma estable y abundante de hierro existente en las hojas, en forma de complejo de hierro y proteína denominado fitoferritina. El acceso de hierro a la corriente de transporte del floema probablemente se minimiza debido a la formación de esos compuestos insolubles, aunque parece ser que la fitoferritina representa un almacén de hierro.
El hierro es esencial porque forma parte de algunas enzimas y numerosas proteínas (constituyendo grupos hemo), que trasladan electrones durante la fotosíntesis y la respiración (Ben-Shem et al., 2003). Experimenta una oxidación y una reducción alternativas, entre los estados de Fe2+ y Fe3+, cuando se comporta como portador de electrones en las proteínas. Un complejo hemo es el grupo prostético de varias proteínas que incluyen la leghemoglobina y las catalasas (Epstein y Bloom, 2005). Estas últimas catalizan la reacción H2O2 →2H2O2 + ½O2 que detoxifica el peróxido de hidrógeno, un proceso importante en la fotorrespiración y en muchas situaciones de estrés.
Manganeso
La microscopía electrónica de los cloroplastos obtenidos de hojas de espinaca muestra que la ausencia de Mn2+ genera la desorganización de las membranas tilacoidales, pero tiene poco efecto sobre la estructura de los núcleos y las mitocondrias. Esta situación, junto con la investigación bioquímica, indica que el elemento tiene una función estructural en el sistema de membranas del cloroplasto, y que una de sus funciones más importantes es, al igual que en el caso del cloro, ayudar a la disociación fotosintética de la molécula de agua. El ion Mn2+ activa numerosas enzimas y es constituyente integral de otras, tales como la Mn-superóxido dismutasa, que provee protección de los radicales libres de oxígeno (Marschner, 2002; Epstein y Bloom, 2005); es cofactor de las prenil transferasas, geranil pirofosfato sintasa y geranil geranil pirofosfato sintasa, las cuales, como se mencionó antes, están involucradas en el alargamiento de la cadena terpénica dentro de la ruta de síntesis de los glucósidos de esteviol.
Zinc
Los inconvenientes producidos por las deficiencias de zinc se dan como resultado de la disminución de crecimiento de las hojas jóvenes y los entrenudos del tallo. Los bordes foliares suelen presentar distorsiones y pliegues. Frecuentemente se produce una clorosis intervenal en gramíneas y frutales, lo que indica que este elemento participa en la formación de la clorofila o bien impide su destrucción. El retraso en el crecimiento del tallo (Epstein y Bloom, 2005) se debe en parte a que quizás sea necesario para producir el ácido indolacético (auxina). Se conocen más de 80 enzimas que contienen zinc unido tan fuertemente, que resulta esencial para su correcto funcionamiento. Adicionalmente, Takatsuji (1999) señala que el Zn2+ hace parte de proteínas activas en la transcripción del DNA.
Cobre
En ausencia de cobre, las hojas jóvenes suelen tomar un color verde oscuro y se presentan arrugadas o deformes; muchas veces tienen manchas necróticas (Mengel y Kirby, 2001). Sin embargo, este elemento en su forma iónica es altamente tóxico a concentraciones elevadas. El cobre está presente en diversas enzimas o proteínas implicadas en los procesos de oxidación y reducción (Fraústo y Williams, 1991; Epstein y Bloom, 2005). Dos ejemplos notables son la citocromo oxidasa, una enzima respiratoria que se encuentra en las mitocondrias, y la plastocianina, una proteína de los cloroplastos. Adicionalmente el cobre es requerido para la fijación de nitrógeno.
Nutrición y producción de hoja seca
A continuación se discuten algunos resultados de trabajos de investigación en requerimientos nutricionales de estevia, haciendo énfasis en los reportados en Brasil para N, P y K por De Lima et al. (1997); se analizan estos resultados con los encontrados por la Universidad de Córdoba (Espitia et al. 2008).
Caso Brasil
Contenido de macronutrientes
Al analizar el contenido de macronutrientes por órgano, De Lima et al. (1997a) determinaron que, en términos generales, durante los primeros 30 d, la mayor tasa de absorción de P, K, Ca, Mg y S se presenta en las ramas, en tanto que el N se acumula preferencialmente en las hojas (Fig. 2). De los 30 a los 60 d, se presenta una uniformidad en cuanto al direccionamiento de los nutrientes entre los órganos de la planta, siendo mayor en flores y frutos, excepto para el S que, entre los 45 y 60 d después del transplante (ddt), presenta una asimilación en las raíces semejante a la de los órganos reproductivos.
Con base en la cantidad y distribución de los nutrientes observados en el trabajo de Brasil, la extracción de los mismos por cada 1.000 kg de hojas secas a los 60 ddt (kg ha-1) será: N: 130; P: 18,8; K: 131,5; Ca: 43,7; Mg: 8,3 y S: 9,7. Si se considera un cultivo comercial, programado para colectas de 2.500 kg ha-1 de hoja seca, se requerirán (kg ha-1): N: 325; P: 47; K: 330; Ca: 109; Mg: 21 y S: 24.
Contenido de micronutrientes
Un trabajo de investigación de De Lima y Malavolta et al. (1997b) indica que en los primeros 30 d, la mayor tasa de acumulación en las ramas, en relación con los demás órganos, ocurre para Cu y Zn, en tanto que Fe y Mn se acumulan de manera más importante en las hojas (Fig. 3). En el período de 30 a 60 ddt, se observó que los micronutrientes se direccionaron de manera similar hacia los órganos reproductivos; excepción a esto son el Fe y el Mn, que continúan acumulándose de manera preferencial en las hojas.
Con base en los valores observados, por cada tonelada de hojas secas cosechadas en el momento de la floración, se espera la extracción (g ha-1) de: B: 89; Cu: 26; Fe: 683; Mn: 207 y Zn: 13. En un programa de producción de semillas (60 ddt), para cada tonelada de hojas secas se prevé la extracción de 226, 76, 2.550, 457 y 33 g ha-1 de B, Cu, Fe, Mn y Zn, respectivamente.
Es importante resaltar que los investigadores no informan el genotipo utilizado y registran resultados hasta los 90 ddt (aunque en esta revisión sólo se presentan hasta los 60 ddt para efectos de comparación con los resultados de Colombia), lo que sin duda debe variar significativamente al considerar que el ciclo productivo de la especie puede ser hasta de cinco años dependiendo de las condiciones ambientales y del manejo. Así mismo, las frecuencias de cosecha de la hoja, que pueden sucederse en periodos de 45 a 60 d de intervalo, también deben ser determinantes para establecer los requerimientos por época de cosecha.
Caso Colombia
En un trabajo llevado a cabo por la Universidad de Córdoba en 2003, se cuantificaron los contenidos de N, P y K en plantas del genotipo Morita 2, en diferentes estados de desarrollo sin discriminar el órgano en el cual estaba presente el elemento (Espitia et al., 2008).
Los resultados indicaron que el contenido de los tres macronutrientes se mantienen relativamente estable a través del tiempo, de manera similar a los resultados del trabajo de De Lima et al. (1997a) en Brasil. Si se comparan los contenidos de los elementos a los 60 ddt en los dos países, se encuentran altas similitudes, a excepción del K que en Colombia registra valores de casi la mitad respecto de los observados en Brasil (Tab. 1).
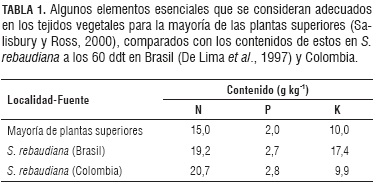
Es probable que el potasio haya sido limitante en los suelos de Colombia respecto a los de Brasil, por lo que los contenidos son menores; otra teoría podría sustentarse en el hecho de que el primer corte de hojas en Colombia se dio a los 90 d, cuando la planta, comparada con la de la otra localidad, presenta un estadio más juvenil y alcanza su madurez a los 120 d, cuando precisamente los contenidos de K por planta alcanzan niveles más cercanos a los observados en Brasil. Sin embargo es importante resaltar que los valores de K observados en la investigación desarrollada por la Universidad de Córdoba son más cercanos a los sugeridos por Salisbury y Ross (2000), quienes reportan contenidos de alrededor de 10 g de K por kg de masa seca de la planta.
Con base en el contenido de nutrientes del trabajo en Colombia a los 60 ddt, la extracción de N, P y K por cada 1.000 kg de hojas secas, será, respectivamente, de 70, 9,2 y 32 kg ha-1 (Jarma, 2008). Al considerar los requerimientos que reportan los investigadores de Brasil para la producción de 1 t ha-1 de hoja seca (respectivamente, 130, 19 y 131 kg ha-1 de N, P y K), se evidencia que las plantas cultivadas bajo la oferta ambiental de Córdoba requieren menor cantidad de los mismos elementos (aproximadamente la mitad de N y P, y la cuarta parte de K). Esto, tal como se discutió anteriormente, puede obedecer, entre otros factores tales como la oferta nutricional del suelo, a la diferencia de estados de desarrollo que registran las plantas a los 60 ddt, ya que, mientras a esa edad las plantas en Brasil están en periodo reproductivo (producción de semilla sexual), en Colombia este estado sólo se alcanza a los 90 d.
Conclusiones
Deficiencias de nitrógeno pueden causar disminución en el crecimiento y desarrollo de todas las partes morfológicas de la planta de estevia, principalmente en hojas, el órgano de interés comercial. El fósforo forma parte esencial de muchos glucofosfatos como la uridin difosfato glucosa, UDP-glc, molécula donadora de glucosa en la síntesis de los glucósidos de diterpeno entre otros procesos metabólicos, y deficiencias de potasio, por su parte, ocasionan reducción en el número de ramas por planta.
Las enzimas que intervienen en la síntesis del ácido ent-kaurenoico (precursor de los principales edulcorantes de las hojas de estevia), son la geranil pirofosfato sintasa, que requiere Mg2+ o Mn2+ como cofactores, la farnesil pirofosfato sintasa, que requiere Mg2+ como cofactor, la geranil geranil pirofosfato sintasa, que requiere Mn2+ para su máxima actividad y la kaureno sintetasa (KS) como regulador en la vía biosintética de los esteviósidos.
El magnesio se combina con el ATP, permitiendo la fosforilación del AMV para formar IPP, primer compuesto isoprenoide en la síntesis de los glucósidos de esteviol. Seguidamente, en la misma ruta de síntesis, actúa como cofactor de la enzima citoplasmática farnesil pirofosfato sintasa, que cataliza la formación del FPP.
Aunque no se reportan muchos trabajos de nutrición en estevia con elementos menores, el papel que desempeñan estos en procesos metabólicos fundamentales, como el desarrollo de la raíz y hojas jóvenes (boro y zinc), la cadena de transporte de electrones (hierro) y procesos de óxido-reducción (cobre), entre otros, presupone la importancia de suplirlos en forma oportuna en programas de nutrición de esta especie.
La extracción de los macronutrientes N, P y K en Brasil y Colombia varían de manera significativa como consecuencia de diversos factores diferenciales (características del suelo, oferta de variables ambientales, genotipo y estados de desarrollo fenológico, entre otros). Como consecuencia de esto, para obtener 1 t ha-1 de hoja seca, en Colombia se requiere la mitad del N y P que reporta Brasil (respectivamente, 70,0 y 9,2 kg ha-1), y la cuarta parte del K (32 kg ha-1).
Literatura citada
Álvarez-García, R., J. Torres-Valencia, L. Román, J. Hernández, C. Cerda-García-Rojas y P. Joseph-Nathan. 2005. Absolute configuration of the a-methylbutyryl residue in longipinene derivatives from Stevia pilosa. Phytochem. 66, 639-642. [ Links ]
Azcón-Bieto, J. y M. Talón. 2008. Fundamentos de fisiología vegetal. McGraw-Hill Interamericana de España, Madrid. [ Links ]
Ben-Shem, A., F. Frolow y N. Nelson. 2003. Crystal structure of plant photosystem I. Nature 426, 630-635. [ Links ]
Brandle, J.E. y P.G. Telmer. 2007. Steviol glycoside biosynthesis. Phytochem. 68, 1855-1863. [ Links ]
Brandle, J.E., A. Richman, A. Swanson y B. Chapman. 2002. Leaf ESTs from Stevia rebaudiana: a resource for gene discovery in diterpene synthesis. Plant Mol. Biol. 50, 613-622. [ Links ]
Buchanan, B., W. Gruissem y R. Jones. 2000. Biochemestry and molecular biology of plants. American Society of Plant Physiologists, Waldorf, MD. [ Links ]
Carakostas, M.C., L.L. Curry, A.C. Boileau y D.J. Brusick. 2008. Overview: the history, technical function and safety of rebaudioside A, a naturally occurring steviol glycoside, for use in food and beverages. Food Chem. Toxicol. 46 (Suppl. 7), 1-10. [ Links ]
Das, K., R. Dang, T.N. Shivananda y N. Sekeroglu. 2007. Influence of bio-fertilizers on the biomass yield and nutrient content in Stevia (Stevia rebaudiana Bert.) grown in Indian subtropics. J. Med. Plants Res. 1(1), 5-8. [ Links ]
Das, K., R. Dang y T.N. Shivananda. 2008. Influence of bio-fertilizers on the availability of nutrients (N, P and K) in soil in relation to growth and yield of Stevia rebaudiana grown in South India. Int. J. Appl. Res. Natl. Prod. 1(1), 20-24. [ Links ]
Davies, T., J. Ying, Q. Xu, Z. Li, J. Li y R. Gordon-Weeks. 2002. Expression analysis of putative high-affinity phosphate transporters in Chinese winter wheat. Plant Cell Environ. 25, 1325- 1340. [ Links ]
De Lima, O. y E. Malavolta. 1997. Sintomas de desordens nutricionais em estévia Stevia rebaudiana (Bert.) Bertoni. Sci. Agric. 54(1-2), 53-61. [ Links ]
De Lima, O., E. Malavolta, J. De Sena y J. Carneiro. 1997a. Absorção e acumulação de nutrientes em estévia Stevia rebaudiana (Bert.) Bertoni: I. Macronutrientes. Sci. Agric. 54(1-2), 23-30. [ Links ]
De Lima, O., E. Malavolta, J. De Sena y J. Carneiro. 1997b. Absorção e acumulação de nutrientes em estévia Stevia rebaudiana (Bert.) Bertoni: II. Micronutrientes. Sci. Agric. 54(1-2), 14-22. [ Links ]
De Lima, O., E. Malavolta y H. Yabico. 1997. Influència de estresses nutricionais no teor e produçâo de esteviosídeo durante o desenvolvimento da Stevia. Pesq. Agrop. Bras. 32(5), 489-494. [ Links ]
Dong, D., X. Peng y X. Yan. 2004. Organic acid exudation induced by phosphorus deficiency and/or aluminium toxicity in two contrasting soybean genotypes. Physiol. Plant. 122, 190-199. [ Links ]
Du, C., Y. Wang, F. Xu, Y. Yang y H. Wang. 2002. Study on the physiological mechanism of boron utilization efficiency in rape cultivars (Brassica napus L.). J. Plant. 25, 231-244. [ Links ]
Epstein, E. y A. Bloom. 2005. Mineral nutrition of plants: principles and perspectivas. Sinauer Associates, Sunderland, MA. [ Links ]
Erkucuk, A., I. H. Akgun y O. Yesil-Celiktas. 2009. Supercritical CO2 extraction of glycosides from Stevia rebaudiana leaves: Identification and optimization. J. Supercrit. Fluids 51, 29-35. [ Links ]
Espitia, M., R. Montoya y A. Jarma. 2008. Stevia en el Caribe colombiano. Universidad de Córdoba, Montería, Colombia. [ Links ]
Espitia, M., R. Montoya y L. Atencio. 2009. Rendimiento de Stevia rebaudiana Bert., bajo tres arreglos poblacionales en el Sinú Medio. Rev. UDCA Actual. Divulg. Cient. 12(1), 151-161. [ Links ]
Fan, X., Q. Shen, H. Zhu, Z. Ma, X. Yin y A. Miller. 2005. A comparison of nitrate transport in four different rice (Oryza sativa L.) cultivars. Science in China Series C Life. Sci. 48(Special issue), 897-911. [ Links ]
Fraústo da Silva, J. y R. Williams. 1991. The biological chemistry of the elements: the inorganic chemistry of life. Clarendon Press, Oxford, UK. [ Links ]
Gardana, C., M. Scaglianti y P. Simonetti. 2010. Evaluation of steviol and its glycosides in Stevia rebaudiana leaves and commercial sweetener by ultra high performance liquid chromatography-mass spectrometry. J. Chromatogr. A 1217(9), 1463-70. [ Links ]
Geuns, J.M. 2003. Stevioside. Phytochem. 64, 913-921. [ Links ]
Geuns, J., P. Augustijns, R. Mols, J. Buyse y B. Driessen. 2003. Metabolism of stevioside in pigs and intestinal absorption characteristics of stevioside, rebaudioside A and steviol. Food Chem. Toxicol. 41, 1599-1607. [ Links ]
Gregersen, S., P. Jeppesen, J. Holst y K. Hermansen. 2004. Antihyperglycemic effects of stevioside in type 2 diabetic subjects. Metabolism. 53(1), 73-76. [ Links ]
Guerra-Ramírez, D., C. Cerda-García-Rojas, A. Puentes y P. Joseph-Nathan. 1998. Longipinene diesters from Stevia lucida. Phytochem. 48(1), 151-154. [ Links ]
Guevara-García, A., C. San Román, A. Arroyo, M. Cortés, M. de la Luz Gutiérrez-Nava y P. León. 2005. Characterization of the arabidopsis clb6 mutant illustrates the importance of posttranscriptional regulation of the methyl-D-erythritol 4-phosphate pathway. Plant Cell 17(2), 628-643. [ Links ]
Hearn, L.K. y P.P. Subedi. 2008. Determining levels of steviol glycosides in the leaves of Stevia rebaudiana by near infrared reflectance spectroscopy. J. Food Comp. Anal. 22(2), 165-168. [ Links ]
Hell, R. y U. Stephan. 2003. Iron uptake, trafficking and homeostasis in plants. Planta 216, 541-555. [ Links ]
Hsieh, M. y H. Goodman. 2005. The Arabidopsis IspH homolog is involved in the plastid nonmevalonate pathway of isoprenoid biosynthesis. Plant Phys. 138(2), 641- 653. [ Links ]
Hsieh, M., P. Chan, Y. Sue, J. Liu, T. Liang, T. Huang, B. Tomlinson, M. Chow, P. Kao e Y. Chen. 2003. Efficacy and tolerability of oral stevioside in patients with mild essential hypertension: a two-year, randomized, placebo-controlled study. Clinical Therapeutics 25(11), 2797-2808. [ Links ]
Jaitak, V., A. Gupta, V. Kaul y P. Ahuja. 2008. Validated high-performance thin-layer chromatography method for steviol glycosides in Stevia rebaudiana. J. Pharm. Biomed. Anal. 47, 790-794. [ Links ]
Jarma, A. 2008. Estudios de adaptación y manejo integrado de estevia (Stevia rebaudiana Bert.): nueva alternativa agroindustrial del Caribe colombiano. Una revisión. Rev. Colomb. Cienc. Hortíc. 2(1), 109-120. [ Links ]
Jarma, A., M. Espitia y G. Fischer. 2010. Síntesis de esteviósidos en estevia (Stevia rebaudiana Bert.). Acta Biol. Colomb. 15(1), 263-268. [ Links ]
Jeppesen, P., S. Gregersen, R. Poulsen y K. Hermansen. 2000. Stevioside acts directly on pancreatic 13 cells to secrete insulin: actions independent of cyclic adenosine monophosphate and adenosine triphosphate-sensitive K+-channel activity. Metabolism 149(2), 208-214. [ Links ]
Jeppesen, P., S. Gregersen, K. Alstrup y K. Hermansen. 2002. Stevioside induces antihyperglycaemic, insulinotropic and glucagonostatic effects in vivo: studies in the diabetic Goto-Kakizaki (GK) rats. Phytomed. 9, 9-14. [ Links ]
Kasahara, H., A. Hanada, T. Kuzuyama, M. Takagi, Y. Kamiya y S. Yamaguchi. 2002. Contribution of the mevalonate and methylerythritol phosphate pathways to the biosynthesis of gibberellins in arabidopsis. J. Biol. Chem. 277(47), 45188-45194. [ Links ]
Kim, K., Y. Sawa y H. Shibata. 1996. Hydroxylation of ent-Kaurenoic acid to steviol in Stevia rebaudiana Bertoni - purification and partial characterization of the enzyme. Arch. Biochem. Biophys. 332(2), 223-230. [ Links ]
Koyama, E., N. Sakai, Y. Ohori, K. Kitazawa, O. Izawa; K. Kakegawa; A. Fujino y M. Ui. 2003. Absorption and metabolism of glycosidic sweeteners of stevia mixture and their aglycone, steviol, in rats and humans. Food Chem. Toxicol. 41, 875-883. [ Links ]
Lian, X., Y. Xing, H. Yan, C. Xu, X. Li y Q. Zhang. 2005. QTLs for low nitrogen tolerance at seedling stage identified using a recombinant inbred line population derived from an elite rice hybrid. Theor. Appl. Genet. 112, 85-96. [ Links ]
Lian, X., S. Wang, J. Zhang, Q. Feng, L. Zhang y D. Fan. 2006. Expression profiles of 10,422 genes at early stage of low nitrogen stress in rice assayed using a cDNA microarray. Plant Mol. Biol. 60, 617-631. [ Links ]
Liao, H., G. Rubio, X. Yan, A. Cao, K. Brown y J. Lynch. 2001. Effect of phosphorus availability on basal root shallowness in common bean. Plant Soil 232, 69-79. [ Links ]
Liu, J., L. Jin-wei y T. Jian. 2010. Ultrasonically assisted extraction of total carbohydrates from Stevia rebaudiana Bertoni and identification of extracts. Food Bioprod. Process. 88(2-3), 215-221. [ Links ]
Marschner, H. 2002. Mineral nutrition of higher plants. 2nd ed. Academic Press, London. [ Links ]
Mengel, K. y E. Kirby. 2001. Principles of plant nutrition. Kluwer Academic Publ., Dordrecht, The Netherlands. [ Links ]
Narissara, L., V. Saengsirisuwan, J. Sloniger, C. Toskulkao y E. Henriksen. 2004. Effects of stevioside on glucose transport activity in insulin-sensitive and insulin-resistant rat skeletal muscle. Metabolism 53(1), 101-107. [ Links ]
O'Neill, M. y W. York. 2003. The composition and structure of plant primary cell walls. pp. 237-263. En: Rose, J.K.C. (ed.). The plant cell wall. Blackwell, Oxford, UK. [ Links ]
Prakash, I., G.E. DuBois, J.F. Clos, K.L. Wilkens y L.E. Fosdick. 2008. Development of rebiana, a natural, noncaloric sweetener. Food Chem. Toxicol. 46(Suppl. 1), 75-82. [ Links ]
Reziwanggu, A., P. Jeppesen, S. Rolfsen, J. Xiao y K. Hermansen. 2004. Rebaudioside A potently stimulates insulin secretion from isolated mouse islets: studies on the dose-, glucose-, and calcium-dependency. Metabolism 53(10), 1378-1381. [ Links ]
Richman, A., M. Gijzen, A. Starratt, Z. Yang y J. Brandle. 1999. Diterpene synthesis in Stevia rebaudiana: recruitment and up-regulation of key enzymes from the gibberellin biosynthetic pathway. Plant J. 19(4), 411-421. [ Links ]
Román, L., G. Morán, J. Hernández, C. Cerda-García-Rojas y P. Joseph-Nathan. 1995. Longipinane derivatives from Stevia Viscida. Phytochem. 38(6), 1437-1439. [ Links ]
RIRDC, Rural Industries Research and Development Corporation. 2002. A new rural industry - Stevia - to replace imported chemical sweeteners. Web Publication No. W02/022. En: http://www.rirdc.gov.au; consulta: julio de 2010. [ Links ]
Salisbury, F.B. y C.W. Ross (eds.). 2000. Fisiología de las plantas 1. Células: agua, soluciones y superficies. Editorial Paraninfo Thomson Learning, Madrid. [ Links ]
Shibata, H., Y. Sawa, T. Oka, S. Sonoke, K. Kim y M. Yoshioka. 1995. Steviol and Steviol-Glycoside: glucosyltransferase activities in Stevia rebaudiana Bertoni- purification and partial characterization. Arch. Biochem. Biophys. 321(2), 390-396. [ Links ]
Su, J., Y. Xiao, M. Li, Q. Liu, B. Li e Y. Tong. 2006. Mapping QTLs for phosphorus-deficiency tolerance at wheat seedling stage. Plant Soil 281(1-2), 25-36. [ Links ]
Suga, T. y T. Endo. 1991. Geranyl diphosphate sinthase in leaves of Pelargonium roseus. Phytochem. 30(6), 1757-1761. [ Links ]
Tadhani, M., V. Patel y S. Rema. 2007. In vitro antioxidant activities of Stevia rebaudiana leaves and callus. J. Food Compos. Anal. 20, 323-329. [ Links ]
Berry, W. 2006. Essensial nutrients, deficiencies and plant dissorders. En Taíz, L. y E. Zeiger (eds.). Plant physiology. 4th ed. Sinauer Associates, Inc., Sunderland, MA. [ Links ]
Takahashi, K., M. Matsuda, K. Ohashi, K. Taniguchi, O. Nakagomi, Y. Abe, S. Mori, N. Sato, K. Okutani y S. Shigeta. 2001. Analysis of anti-rotavirus activity of extract from Stevia rebaudiana. Antiviral Res. 49, 15-24. [ Links ]
Takatsuji, H. 1999. Zinc fingers proteins: the classic zinc fingers emerges in contemporary plant sciences. Plant Mol. Biol. 39, 1073-1078. [ Links ]
Totté, N., L. Charon, M. Rohmer, F. Compernolle, I. Baboeuf y J. Geuns. 2000. Biosynthesis of the diterpenoid steviol, an entkaurene derivative from Stevia rebaudiana Bertoni, via the methylerythritol phosphate pathway. Tetrahedron Lett. 41, 6407-6410. [ Links ]
Totté, N., W. van den Ende, E. van Damme, F. Compernolle, I. Baboeuf y J. Geuns. 2003. Cloning and heterologous expression of early genes in gibberellin and steviol biosynthesis via the methylerythritol phosphate pathway in Stevia rebaudiana Bertoni. Can. J. Bot. 81(5), 517-522. [ Links ]
Utumi, M., P. Monnerat, P. Gomes, P. Resende y V. Campos. 1999. Deficiência de macronutrientes em estévia: sintomas visuais e efeitos no crescimento, composição química e produção de esteviosídeo. Pesq. Agropec. Bras. 34(6), 1039-1043. [ Links ]
White, P. y M. Broadley. 2003. Calcium in plants. Ann. Bot. 92, 487-511. [ Links ]
Wong, K., P. Chan, H. Yang, F. Hsu, I. Liu, Y. Cheng y J. Cheng. 2004. Isosteviol acts on potassium channels to relax isolated aortic strips of Wistar rat. Life Sci. 74, 2379-2387. [ Links ]
Yan, X., P. Wu, H. Ling, G. Xu, F. Xu y Q. Zhang. 2006. Plant nutriomics in China: an overview. Ann. Bot. 98, 473-482. [ Links ]
Zhao, J., J. Fu, H. Liao, Y. He, H. Nian e Y. Hu. 2004. Characterization of root architecture in an applied core collection for phosphorus efficiency of soybean germplasm. Chinese Sci. Bull. 49, 1611-1620. [ Links ]

