










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.28 no.3 Bogotá Sept./Dec. 2010
FITOMEJORAMIENTO, RECURSOS GENÉTICOS Y BIOLOGÍA MOLECULAR
Análisis de la interacción soya-cepa (Bradyrhizobium japonicum) x ambiente, en oxisoles de la Orinoquia colombiana
Analyses of soybean-stain (Bradyrhizobium japonicum) x environment interaction, in oxisols of the colombian Orinoquia
Rubén A. Valencia R.1 y Gustavo Ligarreto M.2
1 Centro de Investigación La Libertad, Investigador de la Corporación Colombiana de investigación Agropecuaria (Corpoica). Villavicencio (Colombia).2 Departamento de Agronomía, Facultad de Agronomía, Universidad Nacional de Colombia. Bogotá (Colombia). Correspondencia: rvalencia@corpoica.org.co
RESUMEN
La Orinoquia colombiana y en particular las sabanas de la altillanura, representa una vasta región con ventajas comparativas y competitivas para la producción de soya. La especie se asocia con bacterias del género Bradyrhizobium para suplir, parcial o totalmente, los requerimientos de nitrógeno. Esta asociación es variable y de alta especificidad huésped-rizobio. La investigación tuvo como propósito determinar la interacción existente entre variedades de soya, cepas del género Bradyrhizobium japonicum y ambiente, en oxisoles de la Orinoquia colombiana, para potenciar el efecto de la fijación biológica del nitrógeno (FBN), en la productividad del cultivo. Se evaluaron las variedades Soyica P-34, Orinoquia 3, C. Libertad 4, C. Taluma 5, C. Superior 6 y C. Sabana 7, con las cepas de B. japonicum ICA J-01, J-96, J-98, mezcla de J-01+J-96 y un control con fertilización con nitrógeno de 150 Kg h-1, en ocho ambientes con variabilidad edafoclimática (C.I. La Libertad, E.E. Taluma de Corpoica en tres semestres del 2007 y 2008, y dos fincas comerciales). La interacción variedad-cepa-ambiente para la variable rendimiento de grano fue determinada mediante los modelos AMMI (Additive main effects and multiplicative interaction) y SREG (Sites regression). El rendimiento de grano varió a través de los ambientes, y estuvo influenciado por el tipo de cepa y variedad de soya. Los biplot generados por los modelos AMMI y SREG permitieron identificar la localidad C.I. Libertad 2008A como el ambiente más discriminante y representativo. El ordenamiento de las variedades x cepa y la identificación de mega-ambientes fueron más consistentes con el modelo SREG (r=0.97, P≤0,01). En general, las variedades presentaron una respuesta positiva a las cepas J96, mezcla de J-01+J-96 y J98, y una respuesta negativa con J-01 y aplicación de N. Las variedades más estables y productivas fueron C. Superior 6 y Orinoquia 3 con la cepa J96. En la interacción, se observó una consistente asociación (r=0,92; P≤0,01) entre balance hídrico y el CP1.
Palabras clave: estabilidad, AMMI, SREG, rendimiento.
ABSTRACT
Colombian Orinoquia savannas and in particular the Altillanura, is a vast region with comparative and competitive advantages for the production of soybeans. Soybean is associated with bacteria of the genus Bradyrhizobium to replace partially or fully the requirements of N. This association is variable and highly specific host-rhizobia. The research was aimed to determine the interaction between soybean varieties and strains of B. japonicum, in Oxisols of the colombian Orinoquia, to boost the effect of biological nitrogen fixation (BNF) in crop productivity. The varieties Soyica P-34, Orinoquia 3, C. Libertad 4, C. Taluma 5, C. Superior 6 and C. Sabana 7, were evaluated with strains J01, J96, J98, a mixture of J01 + J96 and a control with N 150 kg h-1 in eight environments of the Orinoquia. Interaction variety-strain-environment for grain yield was determined by AMMI models (Additive main effects and multiplicative interaction) and SREG (Sites regression). Grain yield varied across environments and was influenced by the strain and soybean variety. The biplot generated by AMMI and SREG models helped identify the location Lib08a as the environment more discriminating and representative. However, the ordering of varieties x strain and identification of mega-environments were more consistent with the SREG model. The main effects of variety x strain (CP1) were high and positively correlated with grain yield (r = 0.97, P≤0.01). In general, the varieties showed a positive response to the strains J96, mix, and J98 and a negative response to J01 and application of N. The C. Superior 6 and Orinoquia 3 varieties were more stable and productive with strain J96. In the interaction, a consistent association was observed (r = 0.92, P≤0.01) between water balance and CP1.
Key word: Stability, AMMI, SREG, yield.Introducción
El creciente aumento en la demanda de alimentos alrededor del mundo es un gran reto para los fitomejoradores quienes buscan plantas de alto potencial genético que mejoren la productividad de los suelos con limitaciones para la producción y puedan ser vinculadas en sistemas agrícolas sostenibles. En Suramérica, aproximadamente 250 millones de hectáreas en sabanas tropicales, incluida la Orinoquia colombiana, presentan serios problemas de fitotoxicidad de aluminio (Vera, 2000), generalmente con bajo contenido de materia orgánica y baja capacidad de intercambio catiónico (Borkert y Sfredo, 1995).
Estas sabanas colombianas representan una vasta región con ventajas comparativas y competitivas para la producción de soya (Valencia et al., 2006). Aunque este cultivo es altamente demandante de nitrógeno, las deficiencias de este elemento se suplen, parcial o totalmente, mediante el proceso de fijación biológica del nitrógeno (FBN) con bacterias del género Bradyrhizobium (González, 2002). Sin embargo, esta asociación entre variedades de soya y cepas de B. japonicum, es variable y depende, en gran medida, de la especificidad huésped-rizobio y de su interacción con el ambiente.
Interacción planta–rizobio–ambiente. Grandes esfuerzos han sido emprendidos en el mejoramiento de cultivares de soya y en la selección de estirpes de B. japonicumi (Hungria et al., 2006). La especificidad de cultivares de soya a cepas de B. japonicum ha sido reportada por varios investigadores (Van Jaarsveld et al., 2002; Krishnan et al., 2003), quienes han confirmado que los genotipos de soya interactúan con las cepas de Bradyrhizobium, donde una cepa específica tiene una alta eficiencia simbiótica con un determinado genotipo. Por lo anterior, es posible mejorar la producción de soya seleccionando apropiadamente, por su compatibilidad, el genotipo y la cepa que se deba utilizar. Otros investigadores, como Zhang et al. (2002) y López- García et al. (2009), han reportado la interacción cepa B. japonicum x ambiente.
La compatibilidad entre el genotipo y la nodulación efectiva de una determinada cepa, tiene control genético parcial (Devine, 1984). Prithiviraj et al. (2000), afirman que la interacción de variedades de soya con las cepas de rizobio, es un proceso de dos pasos: el primero es la relación de las señales moleculares planta-bacteria dado de maneara usual por flavonoides específicos, y el segundo paso, es la liberación de señales moleculares bacteria-planta las cuales son reguladas por los lipo-quito-oligosacáridos (LCOs), también llamados factores Nod.
Las cepas pueden diferir en su capacidad para infectar y fijar N (Neves et al., 1985; George et al., 1987). La infectividad y la efectividad están reguladas genéticamente, pero también están vinculadas con los factores ambientales que afectan a la planta (Fernández, 2003). A pesar de la selección de cepas para mejorar la capacidad de fijación de nitrógeno y la capacidad competitiva, la inoculación de rizobios no mejora el rendimiento de los cultivos, por varios factores como: suelos, clima y cultivo que afectan esta simbiosis (Hunt et al., 1990; Thies, 1990), o por la presencia de cepas naturalizadas altamente competitivas pero poco efectivas para fijar N (Fernández, 2003).
El aumento de la eficacia de la fijación simbiótica de N se puede lograr no sólo mediante la selección de los genotipos superiores, sino mediante la selección de la mejor combinación de genotipos y bacterias fijadoras de N (Rengel, 2002). Además, Kober et al. (2004), sostienen que en la selección de cepas se deben considerar características simbióticas deseables que incluyan alta fijación de nitrógeno, competitividad para formar nódulos y estabilidad genética. En trabajos de investigación de Embrapa se identificaron dos nuevas cepas de B. japonicum, más eficientes y competitivas para los cultivos de soya, en sabanas como la CPAC 7 y CPAC 15. Estas cepas de uso comercial en Brasil, tuvieron su origen en variantes de las cepas SEMIA 5080 y SEMIA 5079 (Franco et al., 1993; Hungria et al., 2001).
Interacción genotipo-ambiente. Los fitomejoradores en sus procesos de selección enfrentan con regularidad el fenómeno complejo conocido como interacción GA (interacción del genotipo - ambiente), que reduce la correlación entre fenotipo y genotipo, y la ganancia genética (Kang y Gorman, 1989; Kang y Gauch, 1996; Kang, 2002;), limita lo que la selección y recomendación de cultivares. La interacción GA generalmente hace alusión a la variación que no puede ser explicada por los efectos principales del genotipo (G) o del ambiente (A).
Varios trabajos de investigación se han realizado para definir el patrón de interacción de genotipos de soya con el ambiente (Zobel et al., 1988; Yan y Rajcan, 2002; Bhardwaj, 2005; Dardanelli et al., 2006), en los que se ha confirmado la presencia de una significativa interacción GA como consecuencia de la respuesta diferencial de los genotipos a cambios ambientales. Kang (2002), sostiene que el efecto combinado del genotipo y de la interacción puede representar entre el 40 y 90% del rendimiento de grano en algunas gramíneas. Por ello, el fitomejorador debe recurrir a los ensayos en multiambientes (EMA) para evaluar el comportamiento relativo de los genotipos e identificar cultivares superiores para una región objetivo (Dehghani et al., 2006).
La interacción GA que tiene un impacto negativo sobre la heredabilidad, ocurre cuando diferentes cultivares o genotipos responden diferencialmente a diversos ambientes. Surge entonces la necesidad de identificar y seleccionar genotipos estables. Desde el punto de vista dinámico o estabilidad agronómica, los genotipos estables en cada ambiente presentan un comportamiento que corresponde al nivel estimado o predicho (Kang, 2002; Yan y Kang, 2003), es decir, que una mejora en el índice ambiental representaría una mejora en el comportamiento fenotípico.
La estabilidad depende de las características que presente la planta como la resistencia a plagas, a enfermedades y a factores de estrés ambiental. Al determinar los factores responsables de la interacción GA o estabilidad/inestabilidad, los fitomejoradores pueden mejorar la estabilidad. Si por ejemplo, la inestabilidad fue causada por susceptibilidad a enfermedades, la consecución de resistencia podría reducir las pérdidas y mejorar la estabilidad (Kang, 2002). Teniendo en cuenta las consideraciones anteriores y la necesidad de generar soluciones tecnológicas para la producción competitiva y sostenible del cultivo, la investigación tuvo como propósito determinar la interacción existente entre variedades de soya, cepas del género B. japonicum y ambiente para la variable rendimiento de grano, en oxisoles de la Orinoquia colombiana como mecanismo para potenciar el efecto de la fijación biológica del nitrógeno (FBN), en la productividad del cultivo.
Materiales y métodos
Localización. Los ensayos multiambientes (EMA), de variedades de soya y cepas de B. japonicum, se realizaron durante los años 2007 y 2008, en oxisoles de la Orinoquia colombiana, en latitudes entre 04°03' N; 73°29' W. Las Localidades fueron identificadas como Lib, Centro de Investigación La Libertad (subregión Piedemonte) y Tal, Estación Experimental Taluma (subregión Altillanura) de la Corporación Colombiana de Investigación Agropecuaria - Corpoica. Cada localidad fue evaluada durante tres semestres (A y B), para un total de 6 ambientes más las localidades Scruz (finca Santa Cruz) y Hor (finca Hortensias) ambas de subregión Altillanura, para un total de 8 ambientes.
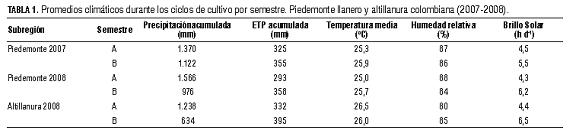
La precipitación acumulada por ciclo de cultivo (mm), la evapotranspiración potencial (ETP) acumulada (mm), la temperatura media (oC), el promedio de la humedad relativa (%) y el brillo solar promedio (h d-1), por semestre y año, se describen en la Tab 1. Se realizaron cálculos acumulados o promedios de cada 4 d de los datos climatológicos desde el día de la siembra hasta la cosecha. La evapotranspiración potencial (ETP) diaria fue estimada mediante el software Daily ver 3 (Granfield University, UK), con base en datos de temperatura mínima y máxima, viento, humedad relativa y brillo solar.
Se realizaron análisis físico-químico de suelos por localdiad. Se determinó la resistencia a la penetración de raíces, medida con un penetrógrafo modelo DIK-5520 (DAIKI Rika Kogyo, Tokio). En ninguno de los lotes hubo siembras previas de soya en su historial. tab 1
La siembra de los experimentos se realizó en el sistema de labranza reducida, con un pase de rastra; incorporación de la cal dolomita un mes antes de la siembra, en dosis basada en la saturación de bases según la fórmula de Raij et al. (1985), y llevada al 50%, un pase de cincel rígido y un pase de pulidor. La siembra se realizó siguiendo las recomendaciones técnicas generadas por Corpoica para el cultivo de soya.
Variedades de soya. Las variedades usadas fueron Soyica P-34, Corpoica Superior-6, Corpoica Libertad 4, Corpoica Taluma 5, Corpoica Sabana 7 y Orinoquia 3. Estas variedades desarrolladas por el Instituto Colombiano Agropecuario – ICA y la Corporación Colombiana de Investigación Agropecuaria – Corpoica, son descritas ampliamente por Valencia et al. (2006).
Cepas de B. japonicum. Se utilizó ICA J-01 generada por el ICA (Salamanca y Ramírez, 2000), las cepas J-96 y J-98 del Banco de Germoplasma de Corpoica introducidas del Brasil y la combinación J-01+J-96. Las cepas de B. japonicum del estudio pertenecen a la siguiente nomenclatura: ICA J-98 o SEMIA 5079, también denominada CPAC 15 y conocida en algunos serogrupos como SEMIA 566; ICA J-96 o SEMIA 5080 o variante CPAC 7; ICA J01 o USDA 110, denominada también 3I1b110, TAL102, RCR3427, 61A89 o SEMIA 5032 (Chen et al., 2000). Para determinar la eficiencia simbiótica relativa de las cepas en estudio, se incluyó un control absoluto sin inoculación con la aplicación de nitrógeno a 150 kg ha-1. El nivel de N fue establecido según estudios de Sánchez et al. (1991), realizados en oxisoles de la Orinoquia colombiana, utilizando como fuente la urea, y fraccionado en dos aplicaciones a los 15 y 30 d después de la siembra.
Diseño experimental. Los tratamientos se distribuyeron en un diseño de franjas divididas en dos factores y en cuatro repeticiones por localidad. El factor A (variedad) y el factor B (cepa de rizobio o N), en distribución aleatoria para conformar bloques de 6 x 5 franjas. Las unidades experimentales fueron conformadas por parcelas de 8 surcos, separados a 40 cm y una distancia de 5 cm entre plantas. Este diseño permitió evaluar los efectos individuales de variedades de cepas y, adicionalmente, la interacción variedad x cepa.
Análisis estadístico de la interacción genotipo-cepaambiente. La información registrada de rendimiento de grano fue procesada con el programa de estadística SAS versión 8 (SAS, 2003). Las medias fueron ajustadas utilizando el procedimiento LSMEANS. Para realizar el análisis multivariado de la interacción de variedades, cepas y ambientes, se conformaron tratamientos a partir de la combinación de variedades por cepas-N, para un total de 30 tratamientos (Vc), con efectos principales Vc y de ambientes (A) e interacción VcA. El análisis combinado de varianza permitió determinar la significancia estadística de la interacción Vc x A.
Los estudios de interacción genotipo-ambiente fueron realizados mediante los modelos AMMI (Additive Main effects and Multiplicative Interaction) y SREG (Sites Regression Model). Se empleó el programa SAS para obtener el modelo AMMI según el algoritmo computacional creado por Vargas y Crossa (2000) y del modelo SREG (Burgueño et al., 2002). Con el modelo SREG se realizaron los análisis de estabilidad fenotípica e identificación de ambientes más discriminatorios y representativos.
Modelo AMMI descrito como:
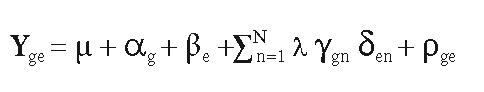

El paquete estadístico y gráfico empleado para procesar ambos análisis multiplicativos fue el propuesto por Burgueño et al. (2002), realizado en SAS y denominado Graphing GE and GGE Biplot (Biometrics and Statistics Unit. CIMMYT. México). El número de términos AMMI y SREG necesarios para una descripción adecuada de la interacción genotipo por ambiente, se evaluó con el método de Gollob (1968).
Resultados y discusión
El análisis combinado de varianza de Vc para rendimiento de grano a través de ambientes, presentó diferencias estadísticas altamente significativas para ambientes (A), var-cepa (Vc) y para la interacción VcA. El rendimiento de grano promedio con las cepas J-96 y J-98 fue superior a J-01 en todas las variedades con excepción de C. Libertad 4, la cual presentó baja respuesta a la inoculación con la cepa J-96 y su combianción con J-01. En la variedad C. Libertad 4, la mejor respuesta se logró con la cepa J-98 y la fertilización nitrogenada. Es de resaltar que con excepción de C. Libertad 4, las otras variedades tuvieron en promedio con las cepas J-96 y J-98, un rendimiento de grano igual o superior a la aplicación de N. La mayor respuesta por ambiente se logró en la Lib2008A con la variedad C. Superior 6 inoculada con la combinación J-01+J-96 (3,64 t ha-1 de rendimiento de grano). Sin embargo, la magnitud de la interacción o la respuesta diferencial a los ambientes hizo que el ranqueo u orden de las variedades y cepas difirieran a través de los ambientes. Por lo anterior, y considerando que el análisis combinado de varianza puede cuantificar la interacción y describir los efectos principales pero no puede explicar la interacción GGA (Kaya et al., 2002), fue importante determinar la contribución de cada componente en la variación total, a través de los modelos AMMI y SREG, los cuales particionan la varianza de la interacción.
Análisis GA del modelo AMMI. El efecto principal del ambiente (A) explicó el 41,9% de la suma de cuadrados total, en tanto que el efecto principal de var-cepa (Vc) explicó el 25,0% de la variación y el 33,0% restante, fue debido a la interacción VcA. Estos resultados indican un alto efecto ambiental sobre el comportamiento de las Vc y que en gran medida, las diferencias en rendimiento de grano se explican por la diversidad de ambientes y su interacción VcA. La alta participación del fenómeno de interacción al ser ésta superior al efecto principal de Vc, demuestra que los Vc (var-cepa) no responden de manera similar a los cambios ambientales. En el modelo AMMI, los dos primeros componentes principales (CP1 y CP2) empleados en la construcción del biplot VcA fueron estadísticamente significativos (P=0,01) y explicaron el 63,6% de la variación de la interacción VcA con el 33,5% de los grados de libertad de la interacción.
Los coeficientes del CP1 y los promedios de rendimiento de grano de los Vc tuvieron una alta correlación (r=0,75; P=0,01), por lo tanto, el CP1 representó significativamente el rendimiento de los Vc y así el biplot pudo ser utilizado para identificar los Vc superiores, como lo sugieren Crossa et al. (2002). Una baja correlación entre los coeficientes de CP1 y los promedios de los genotipos puede ser atribuida a una fuerte y compleja interacción GA (Yan et al., 2001). El rendimiento promedio mostró un amplio rango entre los Vc, de 1,24 a 2,38 t ha-1. Este rango de promedios de rendimiento de grano fue muy amplio, lo que demuestra la presencia de efectos genético-ambientales severos sobre el comportamiento de las variedades y cepas. Por ello, es importante realizar evaluaciones espacio-temporales antes de generar cualquier recomendación sobre variedades.
Las localidades quedaron incluidas en cuatro de cinco sectores del biplot (fig 1). Las variedades Orinoquia 3 y C. Taluma 5 con la cepa J-01 (Vc 2 y Vc 4) tuvieron el mejor comportamiento en el primer semestre en Santa Cruz (Scruz 2008A) y Estación Experimental Taluma (Tal 2008A); de igual manera, la variedad C. Superior 6 con J-98 (Vc 17), en el ambiente Lib 2008A y con la combinación J-01 y J-96 (Vc 20) en el ambiente Lib 2007B, y la variedad C. Libertad 4 con N (Vc 27), en Hor 2008B, son los de mejor comportamiento promedio en los ambientes mencionados. Estos corresponden a los vértices del polígono, y su cercanía a un vector ambiental define su mejor condición ambiental (Yan, 2002). Las var-cepa (Vc), ubicadas cerca al origen (0,0), fueron las que presentaron menor respuesta de interacción que los ubicados en los vértices. La localidad más discriminante fue Lib 2008A, ya que según Yan et al. (2000), estos ambientes presentan un alt o CP1 y un CP2 cercano a cero.
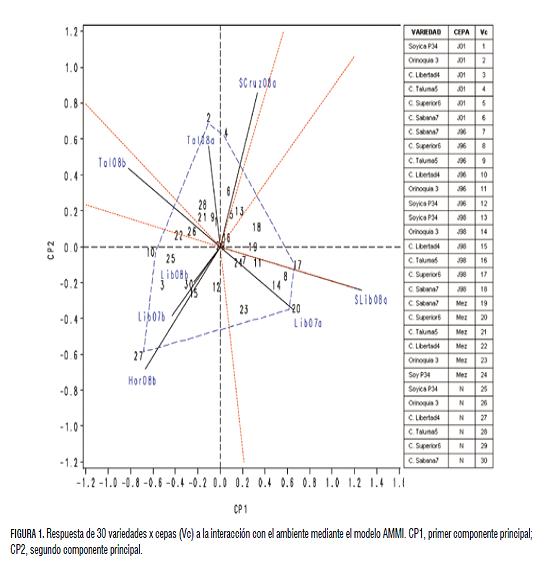
En la localidad Lib2008A más discriminante, se obtuvo el segundo promedio más alto de rendimiento de grano entre localidades (2,29 t ha-1) y el mejor promedio por variedadcepa para C. Superior 6 con la mezcla de J-01+J-96 (3,64 t ha-1), aunque sin diferencias con la J-96 y J-98 (3,38 y 3,32 t ha-1, respectivamente). Por lo anterior, el C.I. La Libertad en evaluaciones de primer semestre sería la condición ideal para procesos de selección de genotipos superiores.
Los mejores Vc por mayor CP1 (atributo de potencial de rendimiento) y bajo CP2 (atributo de estabilidad), fueron: 8, 11, 14, 17 y 20, en todos los casos con rendimientos promedios superiores al promedio general. Estos tratamientos corresponden al comportamiento de las variedades C. Superior 6 y Orinoquia 3 con las cepas J-96, J-98 y la mezcla J-01 y J-96.
Análisis GGA del modelo SREG. Los resultados obtenidos con la aplicación del modelo AMMI tuvieron muchas similitudes a los logrados con el modelo SREG, particularmente al separar los Vc sobre la abscisa (CP1) con una asociación de r=0,89. Las diferencias ambientales entre semestres con gran impacto sobre las variedades y cepas, fueron agrupadas con mayor precisión con el SREG. Estos resultados demuestran la aplicabilidad del SREG para definir los mega-ambientes. Es de resaltar, que en este análisis el biplot GGA despliega simultáneamente el efecto principal de un genotipo (G) y la interacción (GA) de un genotipo x ambiente en una tabla de doble entrada (Yan et al., 2000, 2001; Yan y Kang, 2003), mientras que el AMMI lo hace sólo con la interacción GA.
Con este modelo, el polígono se dividió en siete sectores, en cuatro de los cuales fueron incluidos todos los ambientes evaluados, donde cada sector comparte condiciones ambientales similares. El biplot explicó el 72,9% del GGA, donde el CP1 aportó el 62,1%. Los otros componentes presentaron una participación inferior al 7,6%. Por lo anterior, los dos primeros componentes fueron considerados para la generación del biplot GGA, como lo sigieren Dardanelli et al. (2006). Los efectos principales de Vc (CP1), fueron alta y positivamente correlacionados con rendimiento de grano r=0,97, mientras que con el AMMI esta r=0,75. Yan et al. (2001), mencionan que este valor de asociación debe ser r>0,95 para identificar con certeza los genotipos superiores.
Este tipo de asociación observado también por Yan y Hunt (2001), Crossa et al. (2002), Yan (2002) y Dardanelli et al. (2006), permite el uso del biplot para identificar los Vc superiores y su estabilidad, así como los ambientes más discriminantes y de respuesta similar. Los Vc correspondientes a todas las combinaciones con la cepa J-01, la variedad C. Libertad 4 en todos los casos y con N, presentaron coeficientes negativos. Los demás, presentaron coeficientes CP1 positivos, incluso C. superior 6 con N (fig 2 ).
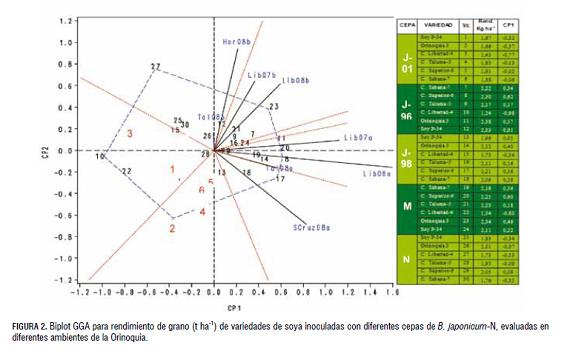
Mejor genotipo por ambiente. Los genotipos que caen sobre el vértice del polígono en un sector determinado, presentan los mejores rendimientos en los ambientes de dicho sector (Yan, 2002; Yan y Hunt, 2002). En la fig 2. , los marcadores de los Vc más alejados en el espacio bidimensional (4, 2, 27, 23, 11, 20, 8 y 17), conectados mediante líneas rectas, configuraron los vértices del polígono. Estos Vc vértices son los que más aportan a la interacción, es decir, los de más alto y más bajo rendimiento. Los Vc ubicados en el margen izquierdo del biplot sobre el eje de la abscisa, presentan baja adaptación a los ambientes evaluados, tal como lo menciona Medina et al. (2001). Los Vc del lado derecho, presentan una relación positiva con los ambientes considerados o que tienden a presentar una adaptación amplia. Estos se encuentran ubicados en el ángulo formado por Scruz 2008A y Hor 2008B. Los genotipos con un alto coeficiente CP1 tienden a dar rendimientos promedios superiores, y localidades con un alto CP1 y un CP2 cercano a cero, facilitan la identificación de tales genotipos (Yan et al., 2000). Los Vc 23, 11, 20, 8 y 17 son los de mejor comportamiento general en los ambientes evaluados, particularmente en el C.I. La Libertad durante el primer semestre del año (A). Los mejores promedios los obtuvieron Vc 8 y 20 en el ambiente Lib 2008A, con rendimientos promedios de grano de 3,38 y 3,64 t ha-1, respectivamente, que corresponden a la variedad C. Superior 6 con la cepa J-96 y la combinación J-01 y J-96. De forma similar, Vc 11 (var. Orinoquia con la cepa J96), se destacó por su alto rendimiento promedio general de 2,38 t ha-1, con su mejor respuesta en la localidad de Scruz 2008A con 2,82 t ha-1. Por otra parte, los Vc 3, 10, 22 y 27 presentan, en general, bajos rendimientos promedios en los ambientes evaluados (1,43; 1,24; 1,34 y 1,73 t ha-1-1, respectivamente), y representan el comportamiento de la variedad C. Libertad 4 con la cepa J-01, J-96, la combinación J-01 y J-96, y N. Esta variedad tuvo su mejor comportamiento en la localidad Hor 2008B con la cepa J-98 y la aplicación de N (2,61 y 2,89 t ha-1, respectivamente).
Relación entre ambientes. Los Vc, correspondientes a un vértice del polígono que se halla comprendido dentro de un sector, tuvieron el mejor rendimiento en los ambientes que se encuentran dentro de ese mismo sector como lo describió Yan et al. (2000). Los ambientes fueron separados por semestre; las localidades del primer semestre Lib 07A, Lib 08A, Tal 08A y Scaruz 08A guardan una estrecha relación entre ellas; así mismo, los ambientes Lib 07B, Lib 08B, Tal 08B y Hor 08B. La mayor correlación se obtuvo entre las localidades Lib07a y Lib08a, o Lib07b y Lib08b, mientras que los más contrastantes fueron Tal08b y Scruz 08a, como se puede observar en el biplot GGA. Esta observación coincide con los coeficientes de correlación, donde las localidades de Lib 07A y Lib 08A del primer semestre tuvieron un r=0,75 (P=0,01), Lib08a con Tal 08A r=0,67 (P=0,01), Lib 07B con Hor 08B r=0,45 (P=0,05). Según Yan (2002) se pueden esperar algunas inconsistencias porque el biplot no explica el 100% de la variación GGA. Por lo anterior, no sobra utilizar el modelo AMMI para reducir el sesgo. Los resultados obtenidos con ambos modelos sugieren una diferencia marcada entre los ambientes, atribuida en gran medida por la época de siembra, más relacionada con la disponibilidad de agua lluvia o balance hídrico entre semestre.
Estabilidad fenotípica del rendimiento de grano. Los coeficientes CP1 y CP2 de los ambientes fueron promediados para crear el punto medio que, al pasar por el origen (0,0), conformó el eje X o CP1 virtual del nuevo biplot Cp1v el cual constituye el potencial de producción, y su perpendicular que pasa por el origen (0,0) conformó el nuevo CP2 virtual Cp2v o vector de estabilidad de los Vc. Yan et al. (2000), sugieren que un genotipo productivo y estable podría tener un alto CP1v pero un CP2v cercano a cero.
Una gran distancia del coeficiente CP2v desde el origen (0,0) del biplot significa alta interacción VcA y, por lo tanto, baja estabilidad. Sobre el margen derecho de la ordenada se ubican las mejores opciones para la selección, destacándose por su amplia adaptación y alto rendimiento los Vc 8, 11 y 20, y con adaptación específica y alto rendimiento los Vc 17 y 23. En contraste, los Vc 3, 10 y 22 obtuvieron los rendimientos promedios más bajos a pesar de su alta estabilidad, lo que significa que son malos en todos los ambientes. El comportamiento típico de un Vc con baja estabilidad y bajo rendimiento es el 27, por lo que se deduce que es un Vc malo en algunos sitios (Tab 2, fig 3 ).

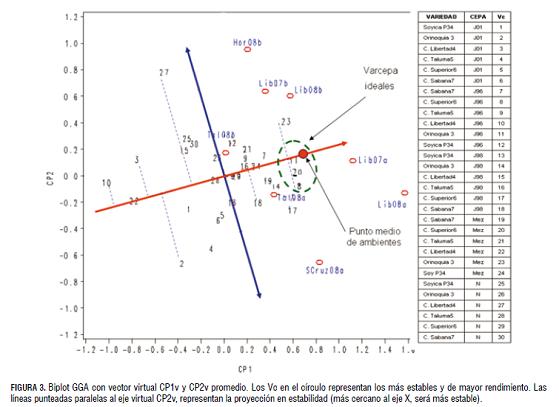
Los modelos AMMI y SREG no permiten establecer las diferencias estadísticas entre genotipos o tratamientos en los biplot, razón por la cual, mediante análisis de contrastes, se seleccionaron las mejores combinaciones de la interacción entre variedades y cepas de B. japonicum, para generar la respectiva recomendación técnica. Las variedades Orinoquia 3 y C. Superior 6 con la cepa J96 fueron las más estables y rendidoras, aunque estadísticamente iguales en comportamiento con la cepa J98. Estas se constituyen en las mejores alternativas para los productores de soya, en oxisoles de la Orinoquia colombiana.
Ambiente más discriminante y representativo. El sector de mayor interés por discriminar los Vc a través de ambientes son aquellos con los valores altos en CP1 y bajos en CP2. Según Yang et al. (2009), los vectores ambientales con CP1 más alejados del origen discriminan mejor los genotipos que aquellos que están cerca del origen. En este sentido, el vector con mayor habilidad discriminante fue la localidad de Lib08a, resultado similar al obtenido con el modelo AMMI.
El menor ángulo de los vectores ambientales formado contra el eje virtual CP1v se presentó con las localidades Lib 07A y Lib 08A, las cuales conforman los ambientes más representativos. En este proceso de evaluación de ambientes, el ambiente ideal altamente discriminante y representativo en promedio fue Lib 08a, donde se logró establecer con mayor claridad el ranqueo de genotipos o Vc. Así, los Vc 11, 20, 8 y 17 se destacaron por su mejor comportamiento en el ambiente Lib 08A. Aunque los resultados fueron similares al modelo AMMI, éste no los ranqueó claramente por su potencial de rendimiento como el SREG. Estos resultados sugieren que el modelo SREG es altamente efectivo para la selección de genotipos estables y altamente promisorios, y que en la localidad del C.I La Libertad en siembras de primer semestre, es posible obtener los mayores beneficios en un programa de mejoramiento genético.
Asociación de PC1 genotípicos (Vc) y las covariables agronómicas. Una asociación positiva y significativa, casi perfecta, se presentó entre los coeficientes CP1 y el efecto del rendimiento de grano de los Vc a través de ambientes, con r=0,97 y significancia P=0,01, por ello, estos CP1 pueden ser interpretados por su efecto principal de Vc, lo que permite deducir que las variables agronómicas que correlacionaron con estos CP1, también están asociadas e interactúan con la producción de grano de las variedades a través de los ambientes en estudio. Las variables asociadas con valores positivos y significativos fueron: vainas totales (VT) y dentro de éstas, las vainas con 3 granos (V3), la altura de planta (ALM) y el número de nudos (NM), el número de nódulos (NN), el peso seco de nódulos (PSN), el contenido total de nitrógeno (CN) en el tejido aéreo y el índice de fijación biológica de nitrógeno (IRFBN). Estos resultados demuestran la importancia de la asociación simbiótica en la producción de grano de soya en el que, cualquier factor externo que afecte su normal desempeño también afectará su rendimiento y sus componentes.
Entre las variables dinámicas, la reacción a enfermedades tuvo un significativo efecto sobre la interacción, particularmente con la presencia de Cercospora sojina, la cual se asocia de manera negativa (r=-0,69; P=0,01), con el CP1 de los Vc (Var x cepa), y con respuesta diferencial entre variedades y localidades, más no entre cepas. Según Kang y Gauch (1996) y Yan y Kang (2003), la resistencia a enfermedades podría incrementar la estabilidad de los cultivares. Con excepción de C. Libertad 4, las otras variedades presentaron tolerancia a este patógeno.
Asociación de PC1 ambientales y las covariables edafoclimáticas. Para establecer la relación, se asumió el criterio expuesto por Yan (2002), donde el ángulo formado entre los vectores de dos ambientes en un biplot GGA está relacionado con el coeficiente de correlación entre ellos. Los suelos en evaluación fueron de texturas francas, con mayor o menor contenido de arcillas. Se encontró asociación negativa entre el contenido de arcillas y el CP1 (-0,72; P=0,01). En este sentido, Santos et al. (2008) encontraron que a mayores incrementos en los contenidos de arcilla en suelos francos, menores serán los rendimientos. En general, la mayor asociación entre ambientes y las covariables climáticas se presentó con el balance hídrico acumulado en el período R3/R6 (de formación a llenado de vainas), con una correlación positiva y altamente significativa (r=0,92). Aún cuando el volumen de precipitación durante el ciclo del cultivo fue superior al requerimiento hídrico total, se presentaron limitantes en la frecuencia con que ocurrió la precipitación. Un déficit hídrico en las etapas críticas del cultivo se presentó en la localidad Lib 07A, al caer la precipitación del 1 al 14 de julio de 2007, donde el balance hídrico fue negativo en 9 de los 14 d, para un bance negativo total de -7,8 mm. Este período coincidió con la fase de llenado de grano y por ello, los rendimientos de grano promedio fueron muy bajos (1,44 t ha-1). King y Purcell (2001), encontraron que el déficit hídrico redujo la fijación simbiótica y, en consecuencia, limitó el rendimiento de grano y de proteína en soya, al favorecerse el aborto y caída de flores, el vaneamiento y caída de vainas. En general, la máxima actividad fijadora ocurre cuando el suelo está en condiciones de capacidad de campo (FAO, 1995). Almanza et al. (2006), determinó que el consumo de agua por el cultivo es de 4,5 mm día-1 y que en el suelo se agotaría su existencia en aproximadamente 4 d, por lo que la ausencia de lluvia durante 4 d en la Altillanura hace necesario el riego.
En la resistencia a la penetración promedia (RP), aunque no se observaron diferencias estadísticas en los coeficientes de correlación, fue notoria la menor resistencia a la penetración a 20 cm de profundidad del suelo en las localidades Lib 07B y Lib 2008A con valores de 0,73 y 1,01 Mpa, respectivamente, mientras que en las otras localidades, estos valores estuvieron entre 1,36 y 2,31 Mpa. Las dos localidades con la menor RP, presentaron los mayores rendimientos de grano promedios (2,46 y 2,29 t ha-1, respectivamente). Bonel y Montico (2007), encontraron que una reducción en rendimiento se relacionaba estrechamente con el aumento de la resistencia a la penetración. La densidad aparente aunque no presentó asociación con rendimiento de grano en los tratamientos evaluados, sus valores promedios fueron iguales o superiores a 1,4 g cm3 en todos los ambientes, afectando el normal crecimiento y desarrollo de las plantas.
Conclusiones
La respuesta diferencial de variedades de soya a la asociación simbiótica con cepas de B. japonicum, abre la posibilidad de seleccionar cepas altamente infectivas y efectivas para potenciar el rendimiento de las variedades de soya, en oxisoles de la Orinoquia colombiana.
Las variedades C. Superior 6 y Orinoquia 3 se constituyen en excelentes alternativas de producción para los oxisoles de la Orinoquia colombiana por su efectiva asociación simbiótica con cepas de B. japonicum, particularmente con J96, su alto potencial de rendimientos de grano y tolerancia a enfermedades.
El modelo SREG fue altamente efectivo para establecer los mega-ambientes y el ambiente más discriminante para ordenar las relaciones variedad-cepa en función de la expresión de su potencial de rendimiento.
Por su alto poder discriminante, el Centro de Investigaciones La Libertad (Villavicencio), se constituye en el mejor ambiente para procesos de selección de genotipos superiores en asocio con cepas específicas de B. japonicum, especialmente en evaluaciones en el primer semestre. Los factores más relevantes de la interacción genotipoambiente (GA) y que de alguna manera afectaron la relación simbiótica y el rendimiento de grano, fueron en su orden: variedad, déficit hídrico, compactación de suelos y enfermedades, particularmente la presencia de Cercospora sojina.
Literatura citada
Almanza M., E.F., J.O. Arguello T., J.H. Bernal R. y S. Caicedo G. 2006. Requerimientos hídricos del cultivo de la soya en la Altillanura. Corpoica, Villavicencio, Colombia. [ Links ]
Bonel, B. y S. Montico. 2007. Las posibilidades del escarificado en suelos del sur de Santa Fe. Rev. Agromensajes 21(4), 26-36. [ Links ]
Borkert, C.M. y G.J. Sfredo. 1995. La fertilización de los suelos tropicales para el cultivo de la soya. pp. 175-200. En: El cultivo de la soya en los trópicos. Mejoramiento y producción. Colección FAO, Producción y Protección Vegetal No. 27. Embrapa; CNPSo, Roma. [ Links ]
Burgueño, J., J. Crossa y M. Vargas. 2002. SAS Programs for graphing GE and GGE biplots. En: Biometrics and Statistics Unit, Cimmyt, http://www.cimmyt.org [Dic.2010]. [ Links ]
Chen, L.S., A. Figueredo, F.O. Pedrosa y M. Hungria. 2000. Genetic characterization of soybean rhizobia in Paraguay. Appl. Environ. Microb. 66(11), 5099-5103. [ Links ]
Crossa, J., P.L. Cornelius y W. Yan. 2002. Biplots of linear-bilinear models for studying crossover genotype x environment interaction. Crop Sci. 42, 619-633. [ Links ]
Dardanelli, J., M. Balzarini, M.J. Martínez, M. Cuniberti, S. Resnik, S. Ramunda, R. Herrero y H. Baigorri. 2006. Soybean maturity groups, environments and their interaction define megaenvironments for seed composition in Argentina. Crop Sci. 46, 1939-1947. [ Links ]
Dehghani, H., A. Ebadi y A. Yousefi. 2006. Biplot analysis of genotype x environment interaction for Barley yield in Iran. Agron. J. 98, 388-393. [ Links ]
Devine, T.E. 1984. Inheritance of soybean nodulation response with a fast-growing strain of Rhizobium. J. Hered. 75(5), 359-361. FAO. 1995. Manual técnico de la fijación biológica del nitrógeno: Leguminosa/Rhizobium. Roma. [ Links ] [ Links ]
Fernández C., M.V. 2003. Manual de nodulación. Editorial Hemisferio Sur, Buenos Aires. [ Links ]
Franco, M. G.A. Ocampo, E. Melo y R. Thomas. 1993. Catalogue of Rhizobium strains for 172 tropical forage legumes. 5a ed. CIAT, Cali, Colombia. [ Links ]
George, T., B.B. Bohlool y P.W. Singleton. 1987. Bradyrhizobium japonicum-environment interactions: nodulation and interstrain competition in soils along an elevational transect. Appl. Environ. Microbiol. 53(5), 1113-1117. [ Links ]
Gollob, H. 1968. A statistical model which combines features of factor analytic and analysis of variances techniques. Psychometrika 33, 73-115. [ Links ]
González, N.S. 2002. Algunos elementos de juicio para interpretar el fenómeno de la nodulación en soja. Publicación de las Jornadas de Cosecha Gruesa. INTA; CIAM, Mar del Plata, Argentina. [ Links ]
Hungria, M., R.J. Campo y I.C. Mendes. 2001. Fixação biológica do nitrogêno na cultura da soja. Embrapa Soja Circular Técnica 35. Embrapa, Londrina, Brasil. [ Links ]
Hungria, M., R.J. Campo, I.C. Mendes y P.H. Graham. 2006. Contribution of biological nitrogen fixation to the nitrogen nutriton of grain crops in the tropics: the success of soybean (Glycine max L. Merr.) in South america. pp. 43-93. En: Singh, R.P., N. Shankar y P.K. Jaiwal (eds.). Nitrogen nutrition in plant productivity. Studium Press, LLCC, Houston, TX. [ Links ]
Hunt, P.G., M.J. Kasperbauer y T.A. Matheny. 1990. Influence of Bradyrhizobium japonicum strain and far-red/red canopy light ratios on nodulation of soybean. Crop Sci. 30(6), 1306-1308. [ Links ]
Kang, M.S. 2002. Quantitative genetics, genomics, and plant breeding. CABI, Wallingford, UK. [ Links ]
Kang, M.S. y D.P. Gorman. 1989. Genotype x environment interaction in maize. Agron. J. 81, 662-664. [ Links ]
Kang, M.S. y H.G. Gauch. 1996. Genotype-by-environment interaction. CRC Press, Boca Ratón, FL. [ Links ]
Kaya, Y., Ç. Palta y S. Taner. 2002. Additive main effects and multiplicative interactions analysis of yield performances in bread wheat genotypes across environments. Turk. J. Agric. For. 26, 275-279. [ Links ]
King, C.A. y L.C. Purcell. 2001. Soybean nodule size and relationship to nitrogen fixation response to water deficit. Crop Sci. 41, 1099-1107. [ Links ]
Kober, M.V., E.L. Saccol de Sá, J.R.J. Freire y A. Giongo. 2004. Characterization of variants of Bradyrhizobium elkanii and B. japonicum and symbiotic behaviour in soybeans. Ciênc. Rural 34(5), 1459-1464. [ Links ]
Krishnan, H.B., J. Lorio, W.S. Kim, G. Jiang, K.Y. Kim, M. DeBoer y S.G. Pueppke. 2003. Extracellular proteins involved in soybean cultivar-specific nodulation are associated with pilus-like surface appendages and exported by a type III protein secretion system in Sinorhizobium fredii USDA257. Mol. Plant Microbe Interact. 16(7), 617-625. [ Links ]
López-García, S.L., A. Perticari, C. Piccinetti, L. Ventimiglia, N. Arias, J.J. De Battista, M.J. Althabegoiti, E.J. Mongiardini, J. Pérez-Giménez, J.I. Quelas y A.R. Lodeiro. 2009. In-furrow inoculation and selection for higher motility enhances the efficacy of Bradyrhizobium japonicum nodulation Agron. J. 101(2), 357-363. [ Links ]
Medina, M., T. Abadie, D. Vilaró y S. Ceretta. 2001. Estudio metodológico de adaptación de cultivares de maíz para silo a las condiciones de Uruguay. Agrociancia 5(1), 23-31. [ Links ]
Neves, M.C.P., A.D. Didonet, F.F. Duque y J. Döbereiner. 1985. Rhizobium strain effects on nitrogen transport and distribution in soybeans. J. Exp. Bot. 36, 1179-1192. [ Links ]
Prithiviraj, B., A. Souleimanov, X. Zhou y D.L. Smith. 2000. Differential response of soybean (Glycine max (L.) Merr.) genotypes to lipo-chito-oligosaccharide Nod Bj V (C18:1 MeFuc). J. Exp. Bot. 51(353), 2045-2051. [ Links ]
Raij, B. Van, N.M. Silva, O.C. Bataglia, J.A. Quaggio, R. Hiroce, H. Cantarella, J.R. Belinazzi Jr., A.R. Dechen y P.E. Trani. 1985. Recomendações de adubação e calagem para o Estado de São Paulo. Boletín Técnico No. 100. Instituto Agronómico de Campinas (IAC), Campinas, Brasil. [ Links ]
Rengel, Z. 2002. Breeding for better simbiosis. Plant Soil 245(1), 147-162. [ Links ]
Salamanca S., C.R. y G.M. Ramírez. 2000. ICA J-01 Biofertilizante para soya. Inoculante de rizobio para los Llanos Orientales. Corpoica; Ministerio de Agricultura; FOCC, Villavicencio, Colombia. [ Links ]
Sánchez S., L.F., J.E. Baquero P. y F. Munevar M. 1991. Limitantes nutricionales para el establecimiento del cultivo de la soya en oxisoles de los llanos Orientales. Suelos Ecuat. 20, 5-16. [ Links ]
Santos, F.C., R.F. Novais, J.C.L. Neves, J.M. Foloni, M.R. Albuquerque Filho y J.C. Ker. 2008. Produtividade e aspectos nutricionais de plantas de soja cultivadas em solos de cerrado com diferentes texturas. Rev. Bras. Ciênc. Solo 32(5), 2015-2025. [ Links ]
SAS. 2003. Statistical analysis systems user's guide. SAS Institute Inc., Cary, NC. [ Links ]
Thies, J.E. 1990. Modeling ecological determinants of the symbiotic performance of introduced rhizobia in tropical soils. Tesis de doctorado. University of Hawaii, Honolulu, HI. [ Links ]
Valencia R., R.A., H. Carmen, H. Vargas y G. Arrieta. 2006. Variedades mejoradas de soya para zonas productoras actuales y potenciales de Colombia. Innovación y Cambio Tecnológico - Corpoica. 4(2-3), 7-15. [ Links ]
Van Jaarsveld, C.M., M.A. Smit y G.H.J. Kruger. 2002. Interaction amongst soybean [Glycine max (L.) Merrill] genotype, soil type and inoculant strain with regard to N2 fixation. J. Agron. Crop Sci. 188(3), 206-211. [ Links ]
Vargas, M. y J. Crossa. 2000. The AMMI analysis and the graph of the Biplot in SAS. En: Biometrics and Statistics Unit, Cimmyt, http://www.cimmyt.org/es/programas-y-unidades/unidades-y-laboratorios/laboratorio-de-informatica-aplicada-a-cultivos/gruposunidades-disciplinarias/unidad-de-biometria-y-estadistica; consulta: diciembre de 2010. [ Links ]
Vera, R.R. 2000. Investigación en sistemas agropastoriles: antecedentes y estrategias. pp. 1-8. En: Guimarães, E.P., J.I. Sanz, I.M. Rao, M.C. Amézquita y E. Amézquita. Sistemas agropastoriles en sabanas tropicales de America Latina. CIAT; Embrapa, Cali, Colombia. [ Links ]
Yan, W. 2002. Singular-value partitioning in biplot analysis of multienvironment trial data. Agron. J. 94, 990-996. [ Links ]
Yan, W. y I. Rajcan. 2002. Biplot evaluation of test sites and trait relations of soybean in Ontario. Crop Sci. 42(1), 11-20. [ Links ]
Yan, W. y L.A. Hunt. 2001. Interpretation of genotype x environment interaction for winter wheat yield in Ontario. Crop Sci. 41, 19-25. [ Links ]
Yan, W. y L.A. Hunt. 2002. Biplot analysis of diallel data. Crop Sci. 42, 21-30. [ Links ]
Yan, W., L.A. Hunt, Q. Sheng y Z. Szlavnics. 2000. Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Sci. 40, 597-605. [ Links ]
Yan, W. y M.S. Kang. 2003. GGE biplot analysis: a graphical tool forbreeders. CRC Press, Boca Raton, FL. [ Links ]
Yan, W., P. Cornelius, J. Crossa y L.A. Hunt. 2001. Two types of GGE biplots for analyzing multi-environment trial data. Crop Sci. 41, 656-663. [ Links ]
Yang, R.-C., J. Crossa, P.L. Cornelius y J. Burgueño. 2009. Biplot analysis of genotype × environment interaction: proceed with caution. Crop Sci. 49, 1564-1576. [ Links ]
Zhang, H., T.C. Charles, B.T. Drisco, B. Prithiviraj y D.L. Smith. 2002. Low temperature–tolerant Bradyrhizobium japonicum strains allowing improved soybean yield in short-season areas. Agron. J. 94, 870-875. [ Links ]
Zobel, R., M. Wright y H. Gauch. 1988. Statistical análisis of a yield trial. Agron. J. 80, 388-393. [ Links ]

