










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.28 no.3 Bogotá Sept./Dec. 2010
Artificial microRNAs and their applications in plant molecular biology
MicroARNs artificiales y sus aplicaciones en biología molecular de plantas
Álvaro Luis Pérez-Quintero1, Camilo López1
1 Department of Biology, Faculty of Science, Universidad Nacional de Colombia, Bogota (Colombia).
Corresponding author: celopezc@unal.edu.co
Recceived for publication: 23 March, 2010. Accepted for publication: 13 December, 2010.
ABSTRACT
Artificial microRNAs (amiRNAs) are modified endogenous microRNA precursors in which the miRNA:miRNA* duplex is replaced with sequences designed to silence any desired gene. amiRNAs are used as part of new genetic transformation techniques in eukaryotes and have proven to be effective and to excel over other RNA-mediated gene silencing methods in both specificity and stability. amiRNAs can be designed to silence single or multiple genes, it is also possible to construct dimeric amiRNA precursors to silence two non-related genes simultaneously. amiRNA expression is quantitative and allows using constitutive, inducible, or tissue-specific promoters. One main application of amiRNAs is gene functional validation and to this end they have been mostly used in model plants; however, their use can be extended to any species or variety. amiRNA-mediated antiviral defense is another important application with great potential for plant molecular biology and crop improvement, but it still needs to be optimized to prevent the escape of viruses from the silencing mechanism. Furthermore, amiRNAs have propelled research in related areas allowing the development of similar tools like artificial trans-acting small interference RNAs (ta-siARNs) and artificial target mimicry. In this review, some applications and advantages of amiRNAs in plant molecular biology are analyzed.
Key words: miRNAs, amiRNAs, viruses, gene silencing, RNA interference.
RESUMEN
Los microARNs artificiales (amiARNs) consisten en precursores de microARNs (miARNs) nativos en los que se reemplazan las secuencias del dúplex miARN:miARN* con secuencias diseñadas para silenciar cualquier gen deseado. Los amiARNs son utilizados en transformación genética de eucariotas y han mostrado ser efectivos, superando otros métodos de silenciamiento génico mediado por ARN en cuanto a especificidad y estabilidad. Los amiARNs pueden ser diseñados para silenciar uno o múltiples genes, su expresión es cuantitativa y permite la utilización de promotores constitutivos, inducibles o tejido-específicos. Una de sus principales aplicaciones es la validación funcional de genes, y para este fin han sido utilizados hasta ahora principalmente en plantas modelo, pero su uso puede extenderse a cualquier especie o variedad. Otra aplicación con gran potencial es la defensa contra virus mediada por amiARNs, sin embargo, aun debe ser optimizada para prevenir el escape de los virus al mecanismo de silenciamiento. Los amiARNs han impulsado además la investigación en áreas relacionadas y el desarrollo de herramientas como los ta-siARNs (trans actins small interference RNAs) artificiales y la imitación de blancos o target mimicry. En esta revisión se analizan algunas de las aplicaciones y ventajas de los amiARNs en biología molecular de plantas.
Palabras clave
: miARNs, amiARNs, virus, silenciamiento génico, ARN de interferencia.Introduction
Eukaryotes use post transcriptional gene silencing (PTGS) as a mechanism of gene regulation and natural defense against invasive nucleic acids, transposons, and other highly repetitive genomic sequences (Bartel, 2004).This mechanism is mediated by different kinds of small RNAs, among which the most widely studied are microRNAs (miRNAs) and small interference RNAs (siRNAs).
miRNAs are molecules of approximately 22 nt originating from nuclear genes. A gene codifying for a miRNA (MIRNA) is first transcribed by the RNA polymerase II in a primary miRNA (pri-miRNA) whose size can range from 100 nt to several kilobases (kb). It is then processed to an intermediate RNA called miRNA precursor (pre-miRNA) by Dicer-like 1 (DCL1) in plants. Pre-miRNAs have a characteristic secondary "hairpin-like" structure, with high and negative fold-free energy, these can have widely varying sizes, commonly between 70 and 400 nt. Then, the pre-miRNA is processed to amiRNA:miRNA* duplex again by DCL1 in plants. The duplex is recruited by the RNA-Induced Silencing Complex (RISC) where the strand with the least stability in the 5' extreme (the mature miRNA) will be retained by an argonaut protein (AGO), while the passenger strant (miRNA*) will be degraded, then this 14 Agron. Colomb. 28(3) 2011 complex will guide the cleavage ortranslational repression of messenger RNAs complementaryto the mature miRNA (targets) (Bartel, 2004; Zhang et al., 2006; Zhu, 2008).
In contrast, siRNAs originate from transgenes, viruses, and transposons and form perfectly complementary double-strand RNA precursors (dsRNAs) (Aravin et al., 2003; Filipowicz et al., 2005). siRNAs silence the same gene from which they come and, from a siRNA precursor multiple siRNAs can be generated and these can be transmitted among cells (Bartel, 2004).
miRNAs have been identified as important gene expression regulators in plants and animals (Jones-Rhoades et al., 2006). Among the different functions established for miRNAs in plants, there are physiological aspects as important as morphogenesis and organ polarity, identity of floral organs and flowering time, hormone signaling, transition from juvenile to adult states, reproduction, and response to different kinds of biotic and abiotic stress (Dugas and Bartel, 2004; Jones-Rhoades et al., 2006; Mallory and Baucheret, 2006).
miRNAs have been studied extensively during recent years, among other reasons because by it being a very elegant and precise mechanism of gene regulation, its manipulation through different methods would offer new possibilities for genetic improvement of eukaryotes. One way of using the miRNAs path to alter the expression of certain genes is by employing artificial miRNAs (amiRNAs), which are precursors of endogenous miRNAs modified for silencing any desired target. This work will analyze some applications of artificial amiRNAs in plants, including functional validation of genes and defense against viruses.
Design of artificial microRNAs
Artificial miRNAs are designed from a precursor of an endogenous miRNA, used as structural support and in which the region of the mature miRNA is replaced with a specific amiRNA sequence complementary to the desired target sequence (Ossowski et al., 2008; Park et al., 2009). The procedure was first developed in animals (Zeng et al., 2002) and then in plants (Parizotto et al., 2004). A miRNA precursor can be modified without affecting its natural processing, as long as the same secondary structure is maintained (Parizotto et al., 2004; Vaucheret et al., 2004). For this, we must replace the mature miRNA sequence along with that of the complementary strand or miRNA* without changing structural aspects like non-complementary regions (mismatches) orbulges (Ossowski et al., 2008).
The first step in designing amiRNAs consists of selecting an amiRNA sequence complementary to the desired gene, which must be optimal regarding specificity and effectiveness (Schwab et al., 2010). If what is sought is to silence one or multiple genes, it is necessary to verify that there is no random mating with other genome regions. To avoid these types of situations, it is ideal to have an annotation of the genome or a significant collection of Expressed Sequence Tags (ESTs) of the studied species. Optimizing by effectiveness is mainly based on the calculation of hybridization energy between the amiRNA and the desired target, ideally the amiRNAs with the lowest hybridization energy, less than -30kcal/mol should be chosen (Schwab et al., 2010). There is a quick and easy way of designing amiRNAs by using the MicroRNA Designer (WMD) web application (Schwab et al., 2006), which has been widely used in recent years (Khraiwesh et al., 2008; Ossowski et al., 2008; Warthmann et al., 2008; Molnar et al., 2009).
To ensure amiRNA effectiveness, there are other considerations based on in vivo mating of endogenous miRNAs with their targets. Total complementarity is preferred of the ~21nt of the amiRNA with the target; if there are mismatches, these should never be in positions 10 or 11 (5'-3' sense), given that this is the cleavage site of the miRNA: target pair, also, mismatches should preferably not be in the 5' region (Mallory et al., 2004; Schwab et al., 2006). Additionally, some authors consider it important to introduce mismatches in the 3' region (Moissiard et al., 2007; Khraiwesh et al., 2008). It is preferable for the miRNA to have an A or a U in position 10 and a U in the first position given that these nucleotides are overrepresented in natural plant miRNAs (Mallory et al., 2004; Reynolds et al., 2004; Schwab et al., 2006). Also, a higher AU content in the 5' extreme and greater GC content in the 3' extreme yields instability to the 5' extreme, which is considered necessary to incorporate the mature strand to the RISC complex (Schwab et al., 2006).
There might also be important effects caused by the mRNA structure next to the targeted region; thereby, we must bear in mind the energy of the miRNA union to the target, as well as the energy needed to "open" the union site (Kertesz et al., 2007; Warthmann et al., 2008).
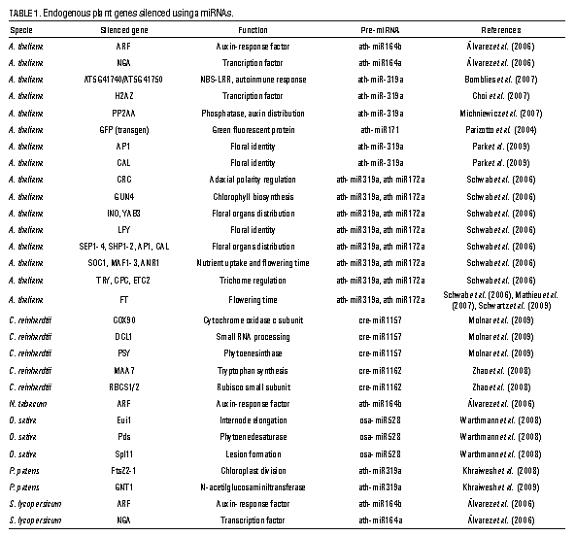
The designed amiRNA and its amiRNA* sequence must replace the miRNA:miRNA* duplex from an endogenous pre-miRNA, some recommended precursors are MIR319a from Arabidopsis, MIR528 from rice, and MIR1157 from Chlamydomonas reinhardtii (tab. 1 ). Once we have the cloned precursors in plasmids, the sequences may be in15 2011 Pérez, López: Artificial microRNAs and their applications in plant molecular biology corporated to the precursors, through guided mutagenesis or overlapping PCR (Schwab et al., 2010). An alternative for replacing the miRNA:miRNA* sequences is the method developed by Molnar et al. (2009), which involves the use of double-strand DNA oligonucleotides to replace DNA from the precursor in the plasmid by using restriction enzymes.
This process, however, may be restricted to small miRNA precursors and has only been implemented until now in Chlamydomonas reinhardtii (Molnar et al., 2009).
A terminator sequence and a promoter sequence must also be added to the plasmid with the precursor. Silencing mediated by amiRNAs occurs in quantitative manner. Constructions containing strong promoters are highly expressed; promoters like 35S from Cauliflower mosaic viruses (CMV) have been broadly used (Ossowski et al., 2008; Zhao et al., 2008; Schwab et al., 2010). However, it is possible to also use inducible or tissue-specific promoters (Schwab et al., 2006). amiRNA can then be inserted in the plant via traditional genetic transformation methods like Agrobacterium-mediated transformation (Schwab et al., 2010).
It is recommended that studies on transformed plants confirm the reduction of transcripts of the desired gene through RT-PCR (Schwab et al., 2010); RACE-PCR could also be used to verify that the mRNA scission occurs in the desired site (Kasschau et al., 2003; Álvarez et al., 2006; Schwab et al., 2010).
Silencing of endogenous amiRNA-mediated genes One of the most common ways to characterize gene function is by analyzing individuals with loss-of-function alleles or mutant genes, in this way the gene's biological activity is inferred from the resulting phenotype (Lukowitz et al., 2000). Several tools have been successful for this type of approach in plants like transpose on tagging, insertional mutagenesis, and tilling, which have been applied successfully, producing extensive and detailed collections of these types of mutants in various species (Alonso and Ecker, 2006; Molnar et al., 2009; Schwab et al., 2010). Nevertheless, saturating complete genomes through this approach requires very large populations of the desired specie and the strategy is normally restricted to certain genetic "contexts" or varieties. Besides, the method can be limited by selectivity in the mutation process and commonly does not permit partial or regulated function loss, which is necessary for studying genes when their null alleles are lethal (Warthmann et al., 2008, Molnar et al., 2009, Schwab et al., 2010).
Alternative methods to obtain loss of gene function are based on RNA-mediated gene silencing (interference RNA or PTGS). RNA transgenes are dominant and can be applied to any variety and to species in which extensive collections of mutants are not yet available. Also, these approaches allow simultaneous silencing of related genes, and gene silencing in an inducible or tissue specific manner (Ossowski et al., 2008; Warthmann et al., 2008).
Common methods of gene silencing are based on the production of dsRNAs that will produce siRNAs to silence the desired gene. The most usual ones consist of 1) the over- expression of a transgene, usually antisense, from the gene sought to be silenced (co-suppression or antisense RNA) (Jorgensen et al., 2006), 2) the sense and antisense expression for the gene to be silenced thus forming a hairpin RNA or hpRNA (Watson et al., 2005), or 3) the insertion of the gene to be silenced in genomes of viruses inoculated on the plant which generate dsRNAs during their replication, this strategy is better known as virus-induced gene silencing (VIGS) (Lu et al., 2003). Although these tools have been broadly used for gene validation and discovery, they still have some difficult ties hindering their broader use in species of agronomic interest (Warthmann et al., 2008). Among its main disadvantages, there are specificity and stability.
These strategies normally use inserts of relatively long size (>200 base pairs), which can generate a great number of siRNAs complementary to distinct regions of the desired gen. Long inserts will most probably generate siRNAs with effective silencing, but will also possibly generate siRNAs complementary or partially complementary to unwanted genome regions (Schwab et al., 2006; Khraiwesh et al., 2008; Warthmann et al., 2008; Molnar et al., 2009). The vast number of possible siRNAs generated by a transgene makes it difficult to predict the unwanted targets (Schwab et al., 2006). It is estimated that between 50 and 70% of the genes in an organism can produce siRNAs complementary to unwanted targets if they are used in interference RNA methods (Xu et al., 2006).Furthermore, the transgenes can be self-silenced, resulting in loss of silencing after several generations (Zhao et al., 2008; Molnar et al., 2009).
amiRNAs become a gene silencing alternative with all the advantages of previous methods of PTGS-mediated silencingand with additional advantages regarding specificity and durability. Successful experiments of amiRNAs-mediated gene silencing have been conducted in dicotyledonous (Arabidopsis, tomato, tobacco) monocotyledonous (rice), mosses (Physcomitrella patens), and algae (Chlamydomonas reinhardtii) (tab.1 ). Different precursors have been used and it has been shown that precursors from different plants 16 Agron. Colomb. 28(3) 2011 can be expressed in others sharing the same miRNA family, producing effective silencing that permits standardizing methods for broad phyologenetic groups (Álvarez et al., 2006; Khraiwesh et al., 2008).
amiRNAs can be designed and optimized to silence one or several genes with similar sequences, including tandem genes, (Álvarez et al., 2006; Schwab et al., 2006; Choi et al., 2007) and have a specificity as high as that of natural plant miRNAs, without formation of secondary siRNAs and without non-autonomous effects (Parizotto et al., 2004; Schwab et al., 2005; Warthmann et al., 2008). Given that an amiRNA produces a unique miRNA with the capacity to silence a specific target, potential unwanted targets can be predicted and avoided from the design (Schwab et al., 2010). In addition, amiRNA transgenes are dominantly inherited and remain stable and active in the progeny (Warthmann et al., 2008; Liu and Chen, 2010). In Chlamydomonas amiRNA activity has remained for up to six months (Zhao et al., 2008) and 500 generations (Molnar et al., 2009). In all, these advantages allow amiRNAs to be considered the most suitable strategy to generate transgenic plants and improve crops (Liu and Chen, 2010).
As with siRNA transgenes, amiRNAs can be expressed in constitutive or inducible manner or via specific tissue according to the promoter used; it has even been possible to generate amiRNAs under the control of the promoter from the same gene sought to be silenced (Schwab et al., 2006). Also, the small size of the precursors used for amiRNAs (<300 base pairs) has allowed for two functional amiRNAs to be simultaneously produced from the same transgene, permitting for two unrelated genes to be simultaneously silenced (Niu et al., 2006; Lin et al., 2009). Given their specificity, amiRNAs can be designed to silence specific alleles or specific forms (E.g., produced via alternative splicing) of certain genes (Ossowski et al., 2008; Schwab et al., 2010).
There is yet no systematic study comparing and showing the advantages of amiRNAs over other interference RNA methods, although Qu et al. (2007) reported a case in which amiRNAs were more effective than hpRNAs in producing silencing. The rate of success of amiRNA-mediated silencing,inferred from published studies, may vary from 90% (Álvarez et al., 2006; Niu et al., 2006; Choi et al., 2007; Mathieu et al., 2007; Qu et al., 2007; Schwab et al., 2006) to 75% (Ossowski et al., 2008). The reasons why some genes cannot be silenced by amiRNAs are not yet clear (Park et al., 2009; Schwab et al., 2010). A possible explanation is the difficulty in accessing the target mRNA by the RISC complex (Ameres et al., 2007; Kertesz et al., 2007; Warthmann et al., 2008; Molnar et al., 2009). This difficulty may, nevertheless, be compensated by thermodynamically modeling the RNA-RNA interactions during the design, as previously described. Another yet unproven possibility may be through negative regulation, such that the silencing produced is compensated with higher rates of transcription that overcome the amiRNA activity (Ossowski et al., 2008).
An additional consideration when working with amiRNAs must be the fact that, although cleavage of the messenger RNA is the predominant mode of action for miRNAs in plants, mRNA translational inhibition has also been reported without knowing yet any adequate way of determining in which instances it occurs (Brodersen et al., 2008). When amiRNA-mediated translational inhibition occurs, the silencing and the phenotypic effects will not be different from those given by mRNA cleavage (Jones-Rhoades et al., 2006; Ossowski et al., 2008). However, this activity may not be monitored and quantified through usual RT-PCR methods (Kasschau et al., 2003; Schwab et al., 2010).
In spite of the potential of the amiRNA method, it has only been used until now in model species (tab. 1 ), particularly in A. thaliana, for which there are even efforts to have a library of amiRNAs covering all the annotated genes (Ossowski et al., 2008), as there is for humans and mice (Chang et al., 2006). Nevertheless, it is only a matter of time for this strategy to expand to other species. Among the Arabidopsis genus amiRNAs have already been used in wild varieties or in non-model lines (Bomblies et al., 2007). Likewise, given the novelty of microRNAs, most studies conducted have concentrated on showing their suitability using genes with already known functions and with those for which we already have null mutants (tab. 1 ).
Some successful and novel functional analysis experiments have been conducted on A. thaliana. For example,by using the tissue-specific expression of amiRNAs, Mathieu et al. (2007) showed that the FT protein and not its mRNAis the mobile signal to induce flowering. Choi et al. (2007) used amiRNAs to demonstrate that H2AZ genes are necessary to activate gene transcription in similar manner as in yeast. Finally, Bomblies et al. (2007) used amiRNAs to show that NBS-LRR type genes AT5G41740/AT5G41750 are involved in the necrosis produced in Arabidopsis hybrid crosses as an autoimmune response.
amiRNA-mediated defense against viruses One of the applications of amiRNAs with the greatest agricultural could be the defense against viruses. Interference 2011 Pérez, López: Artificial microRNAs and their applications in plant molecular biology 17 tab. 1 . Endogenous plant genes silenced using amiRNAs.
RNA methods had already been used in plants to increase their defenses against viruses (Waterhouse et al., 1998; Helliwell and Waterhouse, 2005), in spite of presenting the difficulties mentioned in the case of endogenous genes: poor specificity due to the high probability of unwanted targets and low stability due to self-silencing.
Plants naturally use PTGS to defend themselves against viruses, mainly by siRNA production (Ding et al., 2004; Mahmood-ur-Rahman et al., 2008), and until now, a naturalrole of plant miRNAs for defense against viruses has not been found, as it has been observed in animals (Lecellier et al., 2005). Several authors suggest that plant miRNAs have a potential for defense against viruses (Llave, 2004; Dunoyer and Voinnet, 2005; Lecellier et al., 2005; Simón-Mateo and García, 2006; Lu et al., 2008) and bioinformatics analyses supports this hypothesis (Pérez-Quintero et al., 2010).
By using amiRNAs, it has been possible to show that the miRNA path can work efficiently in defense against viruses.
Transgenic A. thaliana and N. benthamiana plants were created with resistance to Turnip mosaic virus (TMV), Turnip yellow mosaic virus (TuMV), and Cucumber mosaic virus (CMV) using amiRNAs aimed against sequences of these viruses (tab. 2 ). Inthese cases, it was shown that resistance occurs at cellular level and is inheritable and that amiRNAs can successfully block virus replication and avoid its mobility and translocation (Niu et al., 2006). It 18 Agron. Colomb. 28(3) 2011 has also been shown that it is possible to create transgenic plants expressing dimeric amiRNAs aimed against two types of viruses resulting in plants resistant to a broad viral spectrum (Niu et al., 2006; Lin et al., 2009).

As with amiRNAs aimed against endogenous genes, accessibility to the target mRNA of the virus can affect amiRNA effectiveness (Niu et al., 2006; Simón-Mateo and Garcia, 2006; Duan et al., 2008). The secondary structure of the long viral mRNA in vivo is difficult to predict and model. Duan et al. (2008) suggested an experimental method to design effective amiRNAs ensuring target mRNA accessibility.
For this, a test is carried out by inoculating wild and mutant Arabidopsis plants lacking DCLs (proteins responsible for processing small RNAs) with the unwanted viral RNA, viral RNA cleavage sites are compared and, thus, sensitive sites to siRNA-mediated cleavage are identified; these sites could then be used to design amiRNAs against the virus analyzed.
There are other aspects to consider when applying amiRNAs as a strategy to increase plant resistance against viruses. First, many viruses codify silencing suppressors that directly interfere with the miRNA machinery (Llave, 2004; Zhang et al., 2006; Duan et al., 2008). The relatively simple solution of designing amiRNAs aimed against silencing suppressors has proven to be effective (Niu et al., 2006; Qu et al., 2007). The second objection and perhaps the most important is that viral genomes evolve much faster than plant miRNAs, and it has been noted that eventually viruses can evade amiRNA attacks through mutations or deletions in the target region (Simón-Mateo and García, 2006, Lin et al., 2009). This has also been an obstacle in developing gene therapy against viruses like HIV in humans (Das et al., 2004; Westerhout et al., 2005). It is felt that the key to overcome this obstacle lies in the "polymeric strategy", i.e., create transgenic plants with amiRNAs aimed against several regions of the same virus (and even of several viruses), preferably using highly conserved regions (Niu et al., 2006; Duan et al., 2008; Lin et al., 2009). The effectiveness of this strategy has not yet been shown in plants.
amiRNA sin defense against viruses offer as an advantage the preventive feature of this type of defense, meaning that the plant can be resistant to viruses without ever having found them (Lu et al., 2008). Besides, it has been suggested that amiRNA-mediated silencing would pose less problems regarding bio-safety and environmental security with respect to other strategies (Liu and Chen, 2010), bearing in mind that the size of the inserts is relatively small, reducing the probabilities for horizontal transfer of genes and that viral genomes are not used in the transformation.
Production of new allergens or toxic proteins in transgenic plants is equally less likely than with other strategies (Niu et al., 2006). However, studies have not been conducted to extensively assess the safety of this strategy.
Perspectives on research with amiRNAs in plants
The field of plant transformation with amiRNAs is quite novel and we are perhaps far from obtaining plants with desirable agronomical traits through this strategy. Applications in functional gene validation are more immediate and the method is already being accepted as an usual and effective tool in research. There are still many aspects to be clarified on the function of amiRNAs in plants, among others, the way of evaluating and differentiating mRNA cleavage and translational inhibition. For the case of resistance to viruses, we must evaluate the durability of such in transgenic plants obtained with the polymer strategy.
Research with amiRNAs has also influenced development of related techniques like artificial trans-acting small interference RNAs (ta-siRANs) orata-siRNAs (Gutierrez-Nava et al., 2008). ta-siRNAs are a third type of small RNA involved in PTGS whose processing and activity involves siRNA and miRNAs machinery. ta-siRNAs are coded by nuclear genes, whose transcript mRNAis a target for miRNAs, one of the mRNA segments resulting from miRNA scission will produce a dsRNA that will then be processed to produce several siRNAs, which will have as target nuclear genes different to that from which they originate; hence, it is said they act in trans. Gutierrez-Nava et al. (2008) managed to replace the siRNA region in the ta-siRNA coding gene TASc1 (which requires activity from miRNA miR173 to produce siRNAs), successfully silencing the gene from the Fatty Acid Desaturase 2 (FAD2). Gutierrez-Nava et al. (2008) stated that with this strategy multiple genes could be silenced with a sole construct in a simpler manner than with amiRNAs and that for this strategy the secondary structure considerations are not critical.
Likewise, another promising strategy is target mimicry, which consists of inserting a non-cleavable RNA that will act in non-productive manner with a miRNA inhibiting its activity (Franco-Zorrilla et al., 2007). This strategy will permit modulating the level of endogenous microRNAs and allow greater expression of genes that are normally under microRNA regulation and, thus, perhaps improve some agronomic features. This application, however, has not yet been extensively evaluated (Franco-Zorrilla et al., 2007; Liu and Chen, 2010).
The discovery of gene silencing mechanisms mediated by small RNAs, and their implication in defense processes against viruses and regulation of endogenous genes in plants has permitted designing new approaches like amiRNAs to modify gene expression in plant sand, as a result, allow functional gene validation, without needing large populations or completely sequenced genomes for their application. This is of special importance in countries like Colombia where efforts are currently underway for genetic improvement of plants with practically unknown genomes. amiRNAs will surely contribute largely to the development of plant genetics being that it is a simple and effective mechanism, with advantages on specificity, stability, and environmental safety; besides, it is expected that it will allow the development of improved varieties in crops of agricultural interest in a world where increased population and climate change phenomena make up heavy pressure towards safe, innovative, and productive means of agriculture.
Acknowledgments
The authors thank the Research Division in Universidad Nacional at Bogota and Colciencias for the financial support.
Literature cited
Alonso, J.M. and J.R. Ecker. 2006. Moving forward in reverse: genetic technologies to enable genome-wide phenomic screens in Arabidopsis. Nat. Rev. Genet. 7, 524-536. [ Links ]
Álvarez, J., P.I. Pekker, A. Goldshmidt, E. Blum, Z. Amsellem, and Y. Eshed. 2006. Endogenous and synthetic microRNAs stimulate simultaneous, efficient, and localized regulation of multiple targets in diverse species. Plant Cell 18, 1134-1151. [ Links ]
Ameres, S.L., J. Martinez, and R. Schroeder. 2007. Molecular basis for target RNA recognition and cleavage by human RISC. Cell 130, 101-112. [ Links ]
Aravin, A.A., M. Lagos-Quintana, A. Yalcin, M. Zavolan, D. Marks, B. Snyder, T. Gaasterland, J. Meyer, and T. Tuschl. 2003. The small RNA profile during Drosophila melanogaster development. Dev Cell 5, 337-350. [ Links ]
Bartel, D.P. 2004. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116, 281-297. [ Links ]
Bomblies, K., J. Lempe, P. Epple, N. Warthman, C. Lanz, J. Dangl, and D. Weigel. 2007. Autoimmune response as a mechanism for a Dobzhansky-Muller-type incompatibility syndrome in plants. PLoS Biol. 5(9), e236. [ Links ]
Brodersen, P., L. Sakvarelidze-achard, M. Bruun-rasmussen, P. Dunoyer, Y.Y. Yamamoto, L. Sieburth, andO. Voinnet. 2008. Widespread translational inhibition by plant miRNAs and siRNAs. Science 320(5880), 1185-1190. [ Links ]
Chang, K., S.J. Elledge, and G.J. Hannon. 2006. Lessons from nature, microRNA-based shRNA libraries. Nat. Methods 3, 707-714. [ Links ]
Choi, K., C. Park, J. Lee, M. Oh, B. Noh, and I. Lee. 2007. Arabidopsis homologs of components of the SWR1 complex regulate flowering and plant development. Development 134, 1931-1941. [ Links ]
Das, A.T., T.R. Brummelkamp, E.M. Westerhout, M. Vink, M. Madiredjo, R. Bernards, and B. Berkhout. 2004. Human immunodeficiency virus type 1 escapes from RNA interference mediated inhibition. J. Virol. 78, 2601-2605. [ Links ]
Ding, S.W., H. Li, R. Lu, F. Li, and W.X. Li. 2004. RNA silencing, a conserved antiviral immunity of plants and animals. Virus Res. 102, 109-115. [ Links ]
Duan, C.G., C.H. Wang, R.X. Fang, and H.S. Guo. 2008. Artificial microRNAs highly accessible to targets confer efficient virus resistance in plants. J. Virol. 82,11084-11095. [ Links ]
Dugas, D. and B. Bartel. 2004. MicroRNA regulation of gene expression in plants. Curr. Opin. Plant Biol. 7, 512-520. [ Links ]
Dunoyer, P. and O. Voinnet. 2005. The complex interplay between plant viruses and host RNA-silencing pathways. Curr. Opin. Plant Biol. 8, 415-423. [ Links ]
Filipowicz, W., L. Jaskiewicz, F.A. Kolb, and R.S. Pillai. 2005. Post-transcriptional gene silencing by siRNAs and miRNAs. Curr. Opin. Struct. Biol. 15(3), 331-41. [ Links ]
Franco-Zorrilla, J.M., A. Valli, M. Todesco, I. Mateos, M.I. Puga, I. Rubio-Somoza, A. Leyva, D. Weigel, J.A. García, and J. Paz-Ares. 2007. Target mimicry provides a new mechanism for regulation of microRNA activity. Nat. Genet. 39(8), 1033-1037. [ Links ]
Gutierrez-Nava, M.L., M.J. Aukerman, H. Sakai, S.V. Tingey, and R.W. Williams. 2008. Artificial trans-acting siRNAs confer consistent and effective gene silencing. Plant Physiol. 147, 543-551. [ Links ]
Helliwell, C.A., and P.M. Waterhouse. 2005. Constructs and methods for hairpin RNA-mediated gene silencing in plants. Meth. Enzymol. 392, 24-35. [ Links ]
Jones-Rhoades, M.W., D.P. Bartel, and B. Bartel. 2006. MicroRNAs and their regulatory roles in plants. Annu. Rev. Plant Biol. 57, 19-53. [ Links ]
Jorgensen, R.A., N. Doetsch, A. Muller, Q. Que, K. Gendler, and C.A. Napoli. 2006. A paragenetic perspective on integration of RNA silencing into the epigenome and its role in the biology of higher plants. Cold Spring Harb. Symp. Quant. Biol. 71, 481-485. [ Links ]
Kasschau, K.D., Z. Xie, E. Allen, C. Llave, E.J. Chapman, K.A. Krizan, and J.C. Carrington. 2003. P1/HC-Pro, a viral suppressor of RNA silencing, interferes with Arabidopsis development and miRNA function. Dev. Cell 4, 205-217. [ Links ]
Kertesz, M., N. Iovino, U. Unnerstall, U. Gaul, and E. Segal. 2007. The role of site accessibility in microRNA target recognition. Nat. Genet. 39, 1278-1284. [ Links ]
Khraiwesh, B., S. Ossowski, D. Weigel, R. Reski, and W. Frank. 2008. Specific gene silencing by artificial microRNAs in physcomitrella patens; an alternative to targeted gene knockouts. Plant Physiol. 148, 684-693. [ Links ]
Lecellier, C.H., P. Dunoyer, K. Arar, J. Lehmann-Che, S. Eyquem, C. Himber, A. Saïb, and O. Voinnet. 2005. A cellular microRNA mediates antiviral defense in human cells. Science 308(5721), 480-481. [ Links ]
Liu, Q. and Y.Q. Chen. 2010. A new mechanism in plant engineering: the potential roles of microRNAs in molecular breeding for crop improvement. Biotechnol. Adv. 28(3), 301-307. [ Links ]
Lin, S., H. Wu, S. Elena, K. Chen, Q. Niu, S. Ye, C. Chen, and N. Chua. 2009. Molecular evolution of a viral non-coding sequence under the selective pressure of amiRNA-mediated silencing. PLoS Pathog. 5(2), e1000312. [ Links ]
Llave, C. 2004. MicroRNAs: more than a role in plant development? Mol. Plant Pathol. 5(4), 361-366. [ Links ]
Lu, R., A.M. Martin-Hernandez, J.R. Peart, I. Malcuit, and D.C. Baulcombe. 2003. Virus-induced gene silencing in plants. Methods 30, 296-303. [ Links ]
Lu, Y., Q. Gan, X. Chi, and S. Qin. 2008. Roles of microRNA in plant defense and virus offense interaction. Plant Cell Rep. 27, 1571-1579. [ Links ]
Lukowitz, W., C.S. Gillmor, and W. Scheible. 2000. Positional cloning in Arabidopsis. Why it feels good to have a genome initiative working for you. Plant Physiol. 123, 795-805. [ Links ]
Mahmood-ur-Rahman, A.I., T. Husnain and S. Riazuddin. 2008. RNA interference: the story of gene silencing in plants and humans. Biotechnol. Adv. 26(3), 202-209. [ Links ]
Mallory, A.C., B.J. Reinhart, M.W. Jones-Rhoades, G. Tang, P.D. Zamore, M.K. Barton, and D.P. Bartel. 2004. MicroRNA control of PHABULOSA in leaf development, Importance of pairing to the microRNA 59 region. EMBO J. 23, 3356-3364. [ Links ]
Mallory, A., and H. Vaucheret. 2006. Functions of microRNAs and related small RNAs in plants. Nat. Genet. 38(7), 850. [ Links ]
Mathieu, J., N. Warthman, F. Kuttner, and M. Schmid. 2007. Export of FT protein from phloem companion cells is sufficient for floral induction in Arabidopsis. Curr. Biol. 17, 1055-1060. [ Links ]
Michniewicz, M., M. Zago, L. Abas, D. Weijers, A. Schweighofer, I. Meskiene, M.G. Heisler, C. Ohno, J. Zhang, F. Huang, R. Schwab, D. Weigel, E.M. Meyerowitz, C. Luschnig, R. Offringa,and J. Friml. 2007. Antagonistic regulation of PIN phosphorylation by PP2A and PINOID directs auxin flux. Cell 130, 1044-1056. [ Links ]
Moissiard, G., E.A. Parizotto, C. Himber, and O. Voinnet. 2007. Transitivity in Arabidopsis can be primed, requires the redundant action of the antiviral Dicer-like 4 and Dicer-like 2, and is compromised by viral-encoded suppressor proteins. RNA 13, 1268-1278. [ Links ]
Molnar, A., A. Bassett, E. Thuenemann, F. Schwach, S. Karkare, S. Ossowski, D. Weigel, and D. Baulcombe. 2009. Highly specific gene silencing by artificial microRNAs in the unicellular alga Chlamydomonas reinhardtii. Plant J. 58(1), 165-174. [ Links ]
Niu, Q., S. Lin, J. Reyes, K. Chen, H. Wu, S. Ye, and N. Chua. 2006. Expression of artificial microRNAs in transgenic Arabidopsis thaliana confers virus resistance. Nat. Biotechnol. 24(11), 1420-1427. [ Links ]
Ossowski, S., R. Schwab, and D. Weigel. 2008. Gene silencing in plants using artificial microRNAs and other small RNAs. Plant J. 53,674-690. [ Links ]
Parizotto, E.A., P. Dunoyer, N. Rahm, C. Himber, and O. Voinnet. 2004. In vivo investigation of the transcription, processing, endonucleolytic activity, and functional relevance of the spatial distribution of a plant miRNA. Genes Dev. 18, 2237-2242. [ Links ]
Park, W., J. Zhai, and J. Lee. 2009. Highly efficient gene silencing using perfect complementary artificial miRNA targeting AP1 or heteromeric artificial miRNA targeting AP1 and CAL genes. Plant Cell Rep. 28, 469-480. [ Links ]
Pérez-Quintero, A.L., R. Neme, A. Zapata, and C. López. 2010. Plant microRNAs and their role in defense against viruses, a bioinformatics approach. BMC Plant Biol. 10(1), 138-150. [ Links ]
Qu, J., J. Ye, and R. Fang. 2007. Artificial microRNA-mediated virus resistance in plants. J. Virol. 81, 6690-6699. [ Links ]
Reynolds, A., D. Leake, Q. Boese, S. Scaringe, W.S. Marshall, and A. Khvorova. 2004. Rational siRNA design for RNA interference. Nat. Biotechnol. 22, 326-330. [ Links ]
Schwab, R., J. Palatnik, M. Riester, C. Schimmer, M. Schmid, and D. Weigel. 2005. Specific effects of microRNA on the plant transcriptome. Dev. Cell 8, 517-527. [ Links ]
Schwab, R., S. Ossowski, M. Riester, N. Warthmann, and D. Weigel. 2006. Highly specific gene silencing by artificial microRNAs in Arabidopsis. Plant Cell 18, 1121-1133. [ Links ]
Schwab, R., S. Ossowski, N. Warthman, and D. Weigel. 2010. Directed gene silencing with artificial microRNAs. pp. 71-89. In: Meyers, B.C. and P.J. Green (eds.). Plant microRNAs, methods in molecular biology. Vol. 592. Humana Press, Clifton, NJ.Schwartz, C., S. Balasubramanian, N. Warthman, T.P. Michael, J. Lempe, S. Sureshkumar, Y. Kobayashi, J. Maloof, J.O. Borevitz,J. Chory, and D. Weigel. 2009. Cis-regulatory changes at FLOWERING LOCUS T mediate natural variation in flowering responses of A. thaliana. Genetics 183, 723-732. [ Links ]
Simón-Mateo, C., and J. García. 2006. MicroRNA-guided processing impairs plum pox virus replication, but the virus readily evolves to escape this silencing mechanism. J. Virol. 80 (5), 2429-2436. [ Links ]
Vaucheret, H., F. Vazquez, P. Crete, and D.P. Bartel. 2004. The action of ARGONAUTE1 in the miRNA pathway and its regulation by the miRNA pathway are crucial for plant development. Genes Dev. 18, 1187-1197. [ Links ]
Warthmann, N., H. Chen, S. Ossowski, D. Weigel, and P. Herve. 2008. Highly specific gene silencing by artificial miRNAs in rice. PLoS ONE 3, e1829. [ Links ]
Waterhouse, P.M., M.W. Graham, and M.B. Wang. 1998. Virus resistance and gene silencing in plants can be induced by simultaneous expression of sense and antisense RNA. Proc. Natl. Acad. Sci. USA 95, 13959-13964. [ Links ]
Watson, J.M., A.F. Fusaro, M. Wang, and P.M. Waterhouse. 2005. RNA silencing platforms in plants. FEBS Lett. 579, 5982-5987. [ Links ]
Westerhout, E.M., M. Ooms, M. Vink, A. Das, and B. Berkhout. 2005. HIV-1 can escape from RNA interference by evolving an alternative structure in its RNA genome. Nucl. Acids Res. 33, 796-804. [ Links ]
Xu, P., Y. Zhang, L. Kang, M.J. Roossinck, and K.S. Mysore. 2006. Computational estimation and experimental verification of offtarget silencing during posttranscriptional gene silencing in plants. Plant Physiol. 142, 429-440. [ Links ]
Zeng, Y., E.J. Wagner, and B.R. Cullen. 2002. Both natural and designed microRNAs can inhibit the expression of cognate mRNAs when expressed in human cells. Mol. Cell 9, 1327-1333. [ Links ]
Zhang, B., X. Pan, G.P. Cobb, and T. Anderson. 2006. Plant microRNA,a small regulatory molecule with big impact. Dev. Biol. 289, 3-16. [ Links ]
Zhao, T., W. Wang, X. Bai, and Y. Qi. 2008. Gene silencing by artificial microRNAs in Chlamydomonas. Plant J. 58(1), 157-164. [ Links ]
Zhu, J. 2008. Reconstituting plant miRNA biogenesis. Proc. Natl.Acad. Sci. USA 105(29), 9851-9852. [ Links ]

