










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.28 no.3 Bogotá Sept./Dec. 2010
Population structure of Phytophthora infestans in potato crops from Antioquia, Boyaca, Cundinamarca, and Norte de Santander (Colombia)
Estructura poblacional de Phytophthora infestans en cultivos de papa de Antioquia, Boyacá, Cundinamarca y Norte de Santander (Colombia)
Bernardo Silva A.1, José Miguel Cotes2, Mauricio Marín1
1 Laboratory of Cellular and Molecular Biology, Faculty of Science, Universidad Nacional de Colombia, Medellín (Colombia).
2Department of Science Agronomy, Faculty of Agricultural Sciences, Universidad Nacional de Colombia, Medellín (Colombia).
Corresponding author: mamarinm@unal.edu.co
Recceived for publication: 5 March, 2010. Accepted for publication: 13 December, 2010.
ABSTRACT
Potato late blight (Phytophthora infestans), is this crop's most important disease in Colombia. An alternative in reducing the high dependence on fungicides to control the potato late blight includes developing resistant plant varieties, such as the ones recently delivered in Colombia (Pastusa Suprema and Esmeralda), which show high acceptance levels among growers. However, maintenance of resistance in these materials, as well as the effectiveness of systemic fungicides used to control this disease depends on management practices and evolutionary dynamics of P. infestans in each of the potato-growing regions. This research evaluated such a condition through a population structure study of isolates obtained from the departments of Antioquia, Boyaca, Cundinamarca, and Norte de Santander, using AFLP (Amplified Fragment Length Polymorphism) molecular markers. Results indicated the occurrence of low levels of genetic variability within each of the four populations and an important population differentiation among them, especially regarding the population of P. infestans from Antioquia against the middle-east populations. This suggests that besides the tests usually made in Cundinamarca and Boyaca to evaluate resistant materials for P. infestans, different tests in Antioquia should also be included.
Key words: AFLP, genetic variability, molecular markers, potato late blight.
Resumen
La gota de la papa (Phytophthora infestans) es la enfermedad más limitante del cultivo en Colombia. Una alternativa para reducir la dependencia de los fungicidas para el control de la gota, incluye la generación de variedades con resistencia al patógeno, algunas de las cuales (Pastusa Suprema y Esmeralda) se han entregado a los agricultores con buenos niveles de aceptación. Sin embargo, tanto el mantenimiento de los niveles de resistencia de estos materiales como la efectividad de los fungicidas que se utilizan para su control, dependen de las prácticas de manejo y de la dinámica evolutiva de P. infestans en cada región cultivadora. Este trabajo evaluó dicha dinámica a partir del estudio de la estructura poblacional del patógeno en los departamentos de Antioquia, Boyacá, Cundinamarca y Norte de Santander, mediante el empleo de marcadores AFLP (Amplified Fragment Length Polymorphism). Los resultados indicaron la ocurrencia de bajos niveles de variación al interior de las cuatro poblaciones, con una evidente diferenciación entre la población del patógeno de Antioquia con respecto a las de la región centro-oriente del país. Esto indica que la evaluación de variedades mejoradas de papa con resistencia a gota debe realizarse tanto en la sabana Cundiboyacense, como en las regiones cultivadoras de Antioquia.
Palabras clave: AFLP, variabilidad genética, marcadores moleculares, gota de la papa.
Introduction
One of the most limiting diseases in potato crops (Solanum tuberosum and S. phureja) is the potato blight or late blight, caused by the oomycete Phytophthora infestans (Garelik, 2002). P. infestans is considered a serious threat for the planet's food safety, to the point that its detrimental effect on world agriculture has been estimated close to 5-billion US Dollars per year (Duncan, 1999; Kamoun, 2003).
In Colombia, potato blight is limiting in potato crops because of favorable climatic conditions for its development and the cultivation of materials highly susceptible to the pathogen populations (Jaramillo, 2004). Management of this disease has been primarily based on the rotation of fungicides with protective and systemic action (Erwin and Ribeiro, 1996). Nevertheless, the effectiveness of some of these products has been gradually reduced as a result of the onset of resistance in the pathogen populations, given the pressure for selection exerted by current agricultural systems (Gisi and Cohen, 1996; Riveros et al., 2003). Different studies have shown that presence of specific mutations in genes codifying for target proteins of the fungicide action and of certain nucleotide sequences located in said gene promoters, are associated to the occurrence of specific mutations resistant to chemical molecules and that the onset sequence of these genetic changes is directly associated to the recombination rate in the sexual and parasexual cycles presented by fungi and oomycetes, as well as by the genetic structure of populations (Milgroom and Peever, 2003; Damicone, 2004).
A viable alternative to reduce the limitation imposed by the potato blight in the national potato production corresponds to the generation of improved varieties resistant against the pathogen, of which there are success cases like the "Monserrate" variety and the more recent market introductions: "Pastusa Suprema" and "Esmeralda" (Santos et al., 2010). Nevertheless, maintenance of resistance levels of these materials and of others generated in the future will depend on the P. infestans population dynamics.
Traditionally, the populations of P. infestans have been characterized based on different phenotypic and genotypic markers, especially the electrophoretic pattern generated with the RG57 probe in RFLPs (restriction fragment length polymorphism), analysis of Gpi and Pep isoenzymes, level of sensitivity to metalaxyl fungicide, presence of virulent genes determined from differential hosts with R genes, mating type, and evaluation of the mitochondrial haplotypes (Fry et al., 1991; Goodwin et al., 1994; Griffith and Shaw, 1998; Grunwald and Flier, 2005). Using these methodologies has led to determining that the populations of P. infestans are sub-structured from clonal lineages.
Recent studies in Colombia have determined that the population of P. infestans predominant in potato crops presents the IIa Mitochondrial haplotype and the A1 mating type, which is associated to the EC-1 clonal lineage (Raigosa et al., 2009; Gilchrist et al., 2009; Silva et al., 2009). Recently, Vargas et al. (2009) conducted a characterization study of isolations of P. infestans from different solanaceae in the department of Cundinamarca and to a lesser extent in Antioquia, finding isolation with the A2 mating type from Physalis peruviana plants, besides a very low variation level in the population analyzed. In spite of the predominantly clonal condition of the P. infestans populations determined via phenotypic and genotypic evaluations previously described, studies using neutral molecular markers like RAPDs (Random Amplification of Polymorphic DNA) and AFLPs, have revealed the existence of an appreciable degree of genetic variability within the lineages of the oomycete (Mahuku et al., 2000; Páez et al., 2004). A specially interesting work was done by Abu-El Samen et al. (2003), whose RAPD and AFLP electrophoretic patterns (80 RAPD primers and 18 pairs of AFLP primers) from 32 monozoosporic isolates, asexually derived from a parental isolation, presented a high degree of variation, for which the authors suggest that events of mitotic recombination, parasexuality, and spontaneous mutations, play a fundamental role in generating new P. infestans genotypes. Also, studies by Mesa et al. (2008) and Raigosa et al. (2009) using molecular markers in P. infestans populations from different hosts in Colombia have determined the existence of variation levels within the clades representing oomycete genetic lineages reported in the country.
The basis for design of new strategies of chemical or genetic control of potato blight in the country must be founded on studies defining the characteristics of the P. infestans populations in the different potato growing regions. This situation constituted the central objective for this research, when studying the P. infestans populations from the departments of Antioquia, Boyacá, Cundinamarca, and Norte de Santander by using AFLP molecular markers.
Materials and methods
Isolations
We gathered symptomatic samples from plants from different kinds of potato varieties from S. tuberosum ssp. andigena and S. phureja, in the main grower municipalities in the departments of Antioquia, Boyaca, and Cundinamarca.
Additionally, DNA was included of isolates previously collected and stored in the Laboratory of Cellular and Molecular Biology at Universidad Nacional de Colombia in Medellin, from the three departments mentioned and Norte de Santander.
Thus, a total of 40 isolates were analyzed from each of the departments for a total of 160 isolates. From these isolates, 71 were collected from S. tuberosum "Parda Pastusa" variety, 24 from S. tuberosum "Diacol Capiro" variety, 17 from S. phureja "Criolla" Colombia variety, 15 from S. tuberosum ICA "Puracé" variety, and the remaining 33 from other tetraploid regional varieties.
The P. infestans isolates collected in this work were obtained from leaflets and petiole portions with symptoms of potato blight in potato crops. To purify the strains, symptomatic tissues were placed on "Tuquerreña" variety potato slices 2011 Silva Aguilar, Cotes, and Marín: Population structure of Phytophthora infestans... 139 to induce mycelium growth, which was transferred to a rye agar medium (25 g rye meal, 18 g of agar, 20 g of sugar, 1 L distilled water) and kept at 4°C.
AFLP Tests
DNA extraction from the isolates was carried out by following the methodology described by Silva et al. (2009). For the initial standardization of the AFLPs, five isolates were taken from different locations, evaluating 15 primer combinations derived from the restriction sites: EcoRI +0 (5´-GACTGCGTACCAATT-3´) and MseI +0 (5´-GATGAGTCCTGAGTA- 3´): EA-MA, EA-MG, EC-MA, ECMC, EC-MAC, EC-MCC, EC-MG, EC-MCT, EC-MCG, EA-MC, EA-MAC, EA-MCC, EA-MCT, EA-MCG, and EAT-MA. Of these combinations, the most polymorphic and which were consequently selected for the general population analysis corresponded to the primers: EC-MA, EC-MCG, EC-MCC, and EC-MCT.
Thereafter, double digestions were performed with the Eco- RI/MseI enzymes from 5 mL DNA [approx. 100 ng mL-1], 2X Tango Y Buffer + 1.25 U of the restriction enzymes, 1 mM of Dithiothreitol (DTT) to fit a total volume of 12.5 mL. The mixture was incubated at 37°C during 2 h and the reactions were halted at 70°C for 15 min. The effectiveness of the restriction was monitored in agarose gel at 2% for ligation of Ad-MseI adaptors: 5´-GACGATGAGTCCTGAG-3´, 5´-TACTCAGGACTCAT-3´ and Ad-EcoRI: 5´-CTCGTAGACTGCGTACC- 3´, 5´-AATTGGTACGCAGTCTAC-3´.
The ligation reactions consisted in 25 mL of the final volume, including the 12.5 mL of digestion solution, 1 U of the T4 DNA ligase enzyme, 5 pmoles/mL of EcoRI adaptor, 50 pmoles/mL of MseI adaptor, 1 X of enzyme buffer, 0.5 mM of DTT, and 0.4 mM of ATP. The mixture was incubated at 20°C for 2 h and stored at -20°C until its use. To visualize the digestion-ligation, 5 mL of the solution were taken and electrophoresis was conducted in agarose gel at 1.5%.
Then, amplifications were carried out via PCR by using the sets of primers previously indicated. For the final analysis of the AFLP markers, electrophoresis was conducted on polyacrylamide gels at 6% + 7.5 M of urea, using a Biorad electrophoresis chamber with 25 cm gels. This process was begun with a pre-run at 2.300 V and 55°C, to then load 5 mL of the previously denatured sample at 95°C for 5 min and 2 mL of load buffer (100 mL: 0.05 g Bromophenol Blue, 0.05 g Xylene Cyanol, 10 mL Tris/Borate/EDTA10 X, 90 mL Formamide) and it was run at 2.300 V for 80 min. The visualization of the profiles was done by following the methodology of silver staining proposed by Sambrook and Russell (2001).
Data analysis
For each primer combination a binary matrix was constructed based on the presence/absence of each band in the isolates evaluated. For the clustering analysis, a matrix of distances was obtained based on the Jaccard dissimilarity coefficient, and the dendrogram was carried out by using the Unweighted Pair Group Method with Arithmetic Mean (UPGMA) algorithm, validating the structure of the tree based on the value of p (value of p-AU – Approximately Unbiased – 0.05 significance level) and probability of occurrence obtained through bootstrap, using 10,000 resamplings. For this, the pvclust package was used (Susuki and Shimodaira, 2006) from the R statistical program (R development Core Team, 2006).
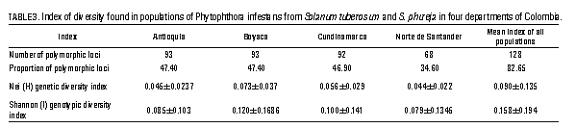
Then, the genotype groups were established based on their region of origin to conduct a molecular analysis of variance (ANOVA) proposed by Excoffier et al. (1992), using the Arlequin v3.1 software (Institute of Ecology and Evolution, University of Bern, Switzerland). A hypothesis test was performed for the components of variance among and within populations using 1.000 permutations. Finally, the number of polymorphic loci was calculated, along with the Nei (H) genetic diversity index, the Shannon (I) genotypic diversity index, and the population sub-structuring statistics: F-statistic (Fst), Nei's population differentiation coefficient (Gst), and the number of average migrants per generation (Nm) through the POPGENE software (University of Alberta, Canada).
Results
The initial evaluation of the 15 primer combinations in five P. infestans isolates indicated a high level of monomorphism, selecting the four combinations resulting slightly polymorphic (EC–MA, EC-MCG, EC-MCC, EC-MCT).
For the construction of the binary matrix, 196 amplicons were selected, distributed thus: EC-MA: 55, EC-MCG: 50, EC-MCC: 47, and EC-MCT: 44 amplicons. The size of these products ranged between 80 and 1.100 bp. In spite of having used the methodology in the 160 isolates, the final analysis was done based on the 148 isolates amplifying for the four pairs of primers (Antioquia: 37, Boyaca: 35, Cundinamarca 39, and Norte de Santander: 37).
The genetic analysis of populations generated a value of Fst = 0.7 and Gst = 0.42 (P = 0.00001), with the genetic distance being greater among the population obtained in Antioquia and that from the Cundinamarca and Boyaca savannah and Norte de Santander (tab. 1 ). The number of average migrants per generation was 0.67. When the
population from Antioquia was not included, the value of Nm increased to 0.98 migrants/generation and the degree of sub-structuring diminished to 0.34. The populations from Boyaca and Cundinamarca presented the least genetic differentiation (Gst = 0.26) and the greatest rates of migration (Nm = 1.35) (tab. 2 ).

The percentage of polymorphic loci ranged between 34 and 47% (82.65% among the four populations), Nei's genetic diversity index was between 0.044 and 0.073 (H = 0.090 for the whole population) and Shannon's genotypic diversity index was between 0.079 and 0.12 (I = 0.158, for the whole population) (tab. 3 ). ANOVA indicated that the total genetic variation of the population under study can be explained in 47.2% by differences among the groups, while 52.8% was due to the variance among the groups.
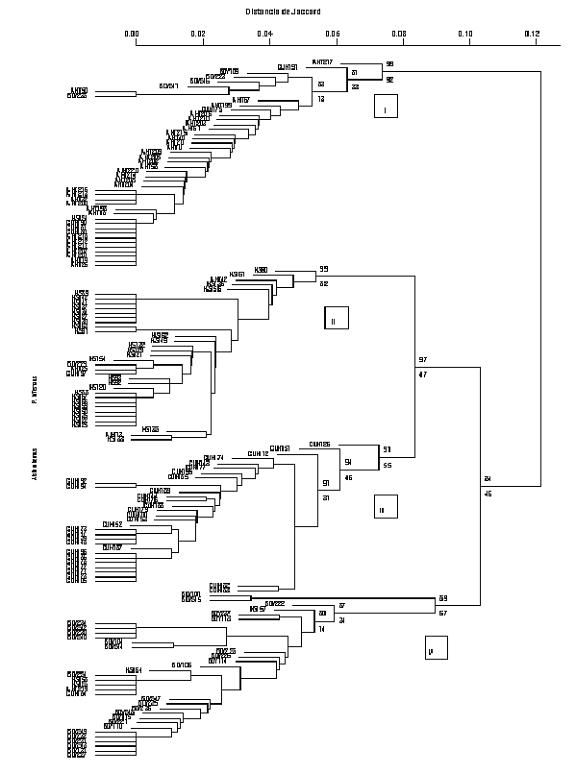
Based on the dendrogram generated, isolates were separated in four groups (I, II, III, and IV) with a maximum distance of 0.12, while within each group there was a very high level of genetic similarity with values of 0.93, 0.94, 0.92, and 0.91 for groups I, II, III, and IV, respectively (fig. 1 ). The statistical analysis supporting these groupings resulted high when using the P value (group I: 99%, group II: 100%, group III: 91%, and group IV: 89%), but not for bootstrap values that presented a range of 56 to 96% (Fig. 1). The first group (I) included 44 isolates that were mostly from the department of Antioquia, although five isolates from Boyaca and five from Cundinamarca were presented. Group II contained 37 isolates, 33 from Norte de Santander, and a very low number from other regions (one from Cundinamarca, one from Boyaca and two from Antioquia); while group III included only isolates from Cundinamarca (32) and group IV was made up of 29 isolates from Boyaca, one from Cundinamarca, one from Antioquia, and four from Norte de Santander.
Discussion
This research presents a genetic study of characteristics of P. infestans populations in potato crops in Colombia, which included four populations from main potato-growing regions in Antioquia, Boyaca, Cundinamarca, and Norte de Santander, whose cultivated area corresponds to 77% of the total crops with this tuber in the country (122,617 ha) (MADR, 2006). Our experimental development was based on the use of AFLP markers, a methodology that is broadly used in these types of studies (Cooke and Lees, 2004; Gotoh et al., 2005) and, hence, constitutes a strong support for the findings in this research.
Work done by Gilchrist et al., (2009) in 2001; Raigosa et al., (2009); Silva et al., (2009) and Vargas et al. (2009) show the occurrence of only one type mating and of one mitochondrial haplotype predominant in the potato P. infestans populations; nevertheless, Vargas et al. (2009) reported in Colombia the presence of an isolate with the A2 type from P. peruviana plants. The genetic variability analysis performed via AFLP markers revealed variation among and within the four populations evaluated. Although said variation levels were low (H = 0.090; I = 0.158), all the isolates evaluated correspond to unique genotypes, which in many cases only diverge by a few loci. These variation levels indirectly indicate the absence of sexual reproduction of this pathogen in Colombia, and support the hypothesis that other mechanisms like parasexuality, high rates of mutations, and presence of extra-chromosomal elements are important in defining P. infestans populations. These mechanisms have already been experimentally detected when evaluating the genotypes derived from monozoosporic crops through RAPD and AFLP molecular markers (Abu-El Samen et al., 2003).
These authors found a considerable level of molecular and phenotypic variation in the monozoosporic populations evaluated, suggesting that genetic changes in asexual P. infestans populations are possible and that these affect the resulting phenotypes, both at the level of pathogenicity, as well as of sensitivity to some fungicides. This situation was confirmed by Gilchrist et al., (2009) in a population of 28 P. infestans isolates from potato crops in Antioquia, which represented 18 pathotypes different from the oomycete and contained in all 10 of the 11 factors of virulence identified in the world to define the P. infestans-S. tuberosum interaction. The studies conducted with molecular markers like AFLPs and microsatellites in P. infestans populations have permitted inferring with greater precision the levels of variability presented by this pathogen and that through the conventional methods defining the genetic lineages (RFLP with the RG57 probe, analysis of Gpi and Pep isoenzymes, mating type, resistance to metalaxyl and mitochondrial haplotypes) were generally masked (Cooke and Lees, 2004). Thus, for example, Cooke et al. (2003) evaluated the P. infestans populations in Scotland between 1995 and 1997, and found that from a sample of 500 isolates, there were only two mitochondrial haplotypes (Ia and IIa) and six RG57 electrophoretic profiles, while the AFLP analysis detected high levels of variability (H= 0.39; I= 0.57) and the presence of at least half of the population evaluated with unique AFLP patterns. Similarly, Knapova and Gisi (2002), when studying P. infestans populations collected between 1996 and 1997 in France and Switzerland, found that for every nine isolates analyzed with microsatellites, one corresponded to a different genotype and that said proportion was two to one, when using AFLP markers.
In the Andean region, two studies have been published on P. infestans populations using AFLP markers. The first of these, corresponds to that conducted by Adler et al. (2004) with a population of the pathogen from different hosts (S. andreanum, S. colombianum, S. caripense, S. muricatum, S. tuberosum, S. betaceum, Brugmansia sanguinea, among others) and which permitted the separation of the population into several clades representing the EC-1, US-1, EC-3 lineages, besides the Ic and Ia haplotypes, associated to new lineages in S. muricatum and in Solanum species from the Anarrhichomenum section, respectively. Within each group several sub-clades were presented, revealed variation among the isolates that comprise them. The main clades presented considerable genetic distances that even led to the description of the new P. andina species from the isolates obtained in S. tetrapetalum of Anarrhichomenum (Adler et al., 2004; Gómez et al., 2007). The other study was conducted by Mesa et al. (2008) with two P. infestans populations collected in the department of Nariño and obtained in potato and tree tomato crops. In said work, the populations from both hosts were located in different clades and presented genetic differentiation (Fst= 0.12; P = 0.0001). Additionally, a certain level of genetic variation was found within each group (H= 0.04; I= 0.06). Such a variation was similar to that found within each of the P. infestans populations from the four departments included in this study (H= 0.04 to 0.07), which may indicate that in spite of the differences in each of the study zones, the pathogen in Colombia presents some similar population parameters, with the absence of sexual reproduction in the populations being a determining factor for this situation.
On the other hand, in this study, the analysis of the population structure from the AFLP markers permitted determining the existence of genetic differentiation among the four populations, especially among the P. infestans population from Antioquia and those from the middleeast part of the country, which present a lower degree of sub-structuring (Gst < 0.3), probably as a result of a greater genetic flow of its populations (Nm > 1.21). Said situation can be due to the differences existing in the environmental conditions in both regions, especially those related with rainfall and temperature, besides the important selection effect represented by the cultivation of different varieties and the differences in the management practices by farmers in said zones. In Antioquia predominates the cultivation of the "Diacol Capiro" variety, while in Boyaca, Cundinamarca, and Norte de Santander the "Parda Pastusa" variety is cultivated most, although there is increased cultivation of improved material resistant to P. infestans, like "Pastusa Suprema" (Santos et al., 2010).
2011 Silva Aguilar, Cotes, and Marín: Population structure of Phytophthora infestans... 143 The other mechanism that could explain the sub-structuring found corresponds to the geographical isolation between the two areas. Nevertheless, when considering the flow of seed tuber between these regions and of fresh commercialized potato, this hypothesis loses value and on the contrary reaffirms the importance in both regions of selection factors of genotypes of the pathogen like climate and agronomic management, more so when the grouping of some isolates was detected in representative clades from different places to their place of origin. These isolates can be considered part of founder populations, which are in the process of adapting to the new conditions of the agro-ecosystems and represent a possible risk, given that they introduce new sets of genes to these zones, which can derive into variants that are more pathogenic or resistant to fungicides used to control potato blight. The greatest variability within the P. infestans population in Antioquia and its difference with the rest of the populations, indicate that the varieties improved through resistance to potato blight must be evaluated in the potatogrowing zones of this department, which coincides with field observations made by Henfling (1987), when making evaluations in the CORPOICA research station "La Selva" in Rionegro, with some families from crosses showing resistance to P. infestans.
Conclusions
The characteristics of the P. infestans population structure in four potato-growing departments in Colombia are presented in this study. The validity of their results is supported by the high number of isolates evaluated (148) and by the use of AFLP molecular markers, which revealed a low level of variation within the geographical populations evaluated. The sub-structuring analysis showed differentiation among the P. infestans population from Antioquia and those obtained from the departments of Boyaca, Cundinamarca, and Norte de Santander.
The results suggest that the occurrence of parasexual phenomena or high rates of mutation in this pathogen are important in the definition of the population characteristics of P. infestans in Colombia and that it is necessary to include the evaluation of the improved potato materials in the cultivation regions of Antioquia, given the differences of the pathogen populations in this department.
Acknowledgments
This research was funded by the Research Office at Universidad Nacional de Colombia (project: 20101005571) and by DIME (Laboratory Enhancement Project, Medellin). The collection of isolates used in the study was carried out with the support from technical personnel from FEDEPAPA and from Professor Sonia Jaramillo and farmers in Antioquia, Boyaca, Cundinamarca, and Norte de Santander.
Literature cited
Abu-El Samen, F.M., G.A. Secor, and N.C. Gudmestad. 2003. Variability in virulence among asexual progenies of Phytophthora infestans (Mont.) De Bary. Phytopathol. 93, 293-304. [ Links ]
Adler, N.F., L.J. Erselius, M.G. Chacón, W.G. Flier, M.E. Ordóñez, L.P. Kroon, and G.A. Forbes. 2004. Genetic diversity of Phytophthora infestans sensu lato in Ecuador provides new insight into the origin of this important plant pathogen. Phytopathol. 94, 154-162. [ Links ]
Cooke, D.E. and A.K. Lees. 2004. Markers, old and new, for examining Phytophthora infestans diversity. Plant Pathol. 53, 692-704. [ Links ]
Cooke, D.E.L., V. Young, P.R. Birch, R. Toth, F. Gourlay, J.P. Day, S.F. Arnegie, and J.M. Duncan. 2003. Phenotypic and genotypic diversity of Phytophthora infestans populations in Scotland. (1995–97). Plant Pathol. 52, 181-192. [ Links ]
Damicone, J. 2004. Fungicide resistance management. Cooperative Oklahoma Extension Service EPP-7663. Stillwater, OK. Duncan, J.M. 1999. Phytophthora an abiding threat to our crops. Microbiol. Today 26, 114-116. [ Links ]
Erwin, D.C. and O.K. Ribeiro. 1996. Phytophthora diseases worldwide. The American Phytopathological Society, St. Paul, MN. Excoffier, L., P.E. Smouse, and J.M. Quattro. 1992. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genet. 131, 479-491. [ Links ]
Fry, E.W., A. Drenth, L.J. Spielman, B.C. Mantel, L.C. Davidse, and S. Goodwin. 1991. Population genetic structure of Phytophthora infestans in the Netherlands. Phytopathol. 81, 1330-1336. [ Links ]
Garelik, G. 2002. Taking the bite out of potato blight. Science 278, 1702-1704. [ Links ]
Gilchrist, E., S. Jaramillo, L. Afanador, and R. Arango. 2009. Caracterización de las poblaciones de Phytophthora infestans en Antioquia, Colombia. Rev. Fac. Nal. Agr. Medellín 62, 5031-5037. [ Links ]
Gisi, U. and Y. Cohen. 1996. Resistance to phenylamide fungicides: a case study with Phytophthora infestans involving mating type and race structure. Annu. Rev. Phytopathol. 34, 549-572. [ Links ]
Gómez, L.A., I. Carbone, and J.B. Ristaino. 2007. An andean origin of Phytophthora Infestans inferred from mitochondrial and nuclear gene genealogies. Proc. Natl. Acad. Sci. USA 9, 3306-3311. [ Links ]
Goodwin, S.B., B.A. Cohen, and W.E. Fry. 1994. Pan global distribution of a single clonal lineage of the Irish potato famine fungus. Proc. Natl. Acad. Sci. USA 91, 11591-11595. [ Links ]
Gotoh, K., S. Akino, A. Maeda, N. Kondo, S. Naito, M. Kato, and A. Ogoshi. 2005. Characterization of some Asian isolates of Phytophthora infestans. Plant Pathol. 54, 733-739. [ Links ]
Griffith, G.W. and D.S. Shaw. 1998. Polymorphisms in Phytophthora infestans: four mitochondrial haplotypes are detected after PCR amplification of DNA from pure cultures or from host lesions. Appl. Environ. Microbiol. 10, 4007-4014.144 Agron. Colomb. 28(3) 2011 [ Links ]
Grunwald, N. and W.G. Flier. 2005. The biology of Phytophthora infestans at its center of origin. Annu. Rev. Phytopathol. 43, 171-190. [ Links ]
Henfling, J.W. 1987. El tizón tardío de la papa: Phytophthora infestans. Boletín de Información Técnica 4. CIP, Lima. [ Links ]
Jaramillo, S. 2004. Monografía sobre Phytophthora infestans (Mont.) de Bary. Universidad Nacional de Colombia. Medellin, Colombia. [ Links ]
Kamoun, S. 2003. Molecular genetics of pathogenic oomycetes. Eukar. Cell 2, 191-199. [ Links ]
Knapova, G. and U. Gisi. 2002. Phenotypic and genotypic structure of Phytophthora infestans populations on potato and tomato in France and Switzerland. Plant Pathol. 51, 641-653. [ Links ]
Mahuku, G., R.D. Peters, H.W. Platt, and F. Daayf. 2000. Random amplified polymorphic DNA (RAPD) analysis of Phytophthora infestans isolates collected in Canada during 1994 to 1996. Plant Pathol. 49, 252-260. [ Links ]
Mesa, V., M.F. Mideros, S. Jaramillo-Villegas, J.M. Cotes, L.E. Lagos, R. Pineda, and M. Marín. 2008. Variabilidad genética de aislamientos de Phytophthora infestans procedentes del suroeste de Colombia. Rev. Iberoam. Micol. 25, 167-172. [ Links ]
Milgroom, M.G. and T.L. Peever. 2003. Population biology of plant pathogens: the synthesis of plant disease epidemiology and population genetics. Plant Dis. 87, 608-617. [ Links ]
MADR, Ministerio de Agricultura y Desarrollo Rural. 2006. Cadena productiva de la papa en Colombia. Bogota. [ Links ]
Páez, O., R. Valverde, L. Gómez, and A. Brenes. 2004. Diversidad genética de aislamientos de Phytophthora infestans en plantaciones de papa en Costa Rica con el uso de RAPDs. Agron. Costarric. 29, 41-55. [ Links ]
Raigosa, N., M.C. Amaya, S. Jaramillo, L.E. Lagos, and M. Marín. 2009. Variabilidad genética de aislamientos colombianos de Phytophthora infestans (Mont.) de Bary. Rev. Fac. Nal. Agr. Medellín 62, 4761-4771. [ Links ]
Riveros, F., R. Sotomayor, V. Rivera, G. Secor and B. Espinoza. 2003. Resistance of Phytophthora infestans (Montagne) de Bary to metalaxyl in potato crops in Northern Chile. Agric. Téc. 63, 39-46. [ Links ]
Sambrook, J. and D.W. Russell. 2001. Molecular cloning: a laboratory manual. 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. pp. 5.26-5.50. [ Links ]
Santos, M., M. Segura, and C.E. Ñústez. 2010. Análisis de crecimiento y relación fuente-demanda de cuatro variedades de papa (Solanum tuberosum L.) en el municipio de Zipaquirá (Cundinamarca, Colombia). Rev. Fac. Nal. Agr. Medellín 63, 5253-5266. [ Links ]
Silva, B., S. Jaramillo, and M. Marín. 2009. Caracterización genética de aislamientos de Phytophthora infestans de papa cultivada en Antioquia, Boyacá, Cundinamarca y Norte de Santander (Colombia). Actual. Biol. 31, 5-20. [ Links ]
Susuki, R. and H. Shimodaira. 2006. Pvclust : An R package for assessing the uncertainty in hierarchical clustering. Bioinform. 22, 1540-1542. [ Links ]
Vargas, A.M., L.M. Quesada-Ocampo, M.C. Céspedes, N. Carreño, A. González, A. Rojas, A.P. Zuluaga, K. Myers, W.E. Fry, P. Jiménez, A.J. Bernal, and S. Restrepo. 2009. Characterization of Phytophthora infestans populations in Colombia: First report of the A2 mating type. Phytopathol. 99, 82-88. [ Links ]

