










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.29 no.1 Bogotá Jan./Apr. 2011
Evaluation of an Andean common bean reference collection under drought stress
Evaluación de una colección de referencia de fríjol andino bajo condiciones de sequía
Juan Carlos Pérez-Vega1, Matthew W. Blair2,4, Fredy Monserrate1, and Gustavo Ligarreto M.3
1 International Center for Tropical Agriculture (CIAT). Palmira (Colombia).
2 Agricultural Sciences Faculty, Universidad Nacional de Colombia. Palmira (Colombia).
3 Department of Agronomy, Faculty of Agronomy, Universidad Nacional de Colombia. Bogota (Colombia).
4 Corresponding author: mwbeans@gmail.com
Received for publication: 27 November, 2009. Accepted for publication: 2 February, 2011.
ABSTRACT
More than 60% of common bean (Phaseolus vulgaris L.) production worldwide is impacted by the risk of drought. In this study, the goal was to evaluate 64 bush bean genotypes from the CIAT reference collection to identify possible sources of drought resistance in the Andean gene pool. Phenotypic traits such as yield, 100-seed weight (P100) and days to physiological maturity (Dpm) were evaluated on selected accessions of this collection which was grown in an 8x8 lattice with two repetitions under three environments: intermittent drought (SI) and irrigation (R) in Palmira as well as early drought (ST) in Darién, Colombia. The genotypes included 20 from the Nueva Granada 1 (NG1) sub-race, 19 from the Nueva Granada 2 (NG2) sub-race, 10 from race Peru (P), 14 Andean control genotypes and one Mesoamerican check. The variables were analyzed through a combined ANOVA across environments, while simple correlations between yield and others variables were determinate. The genotypes with better adaptation to drought showed higher yields, 100-seed weight and fewer days to physiological maturity. The coefficients of correlations among yield and 100-seed weight were significant and positive, while Dpm showed negative correlation. Fourteen genotypes were identified as drought tolerant: G4001, G5625, G6639, G16115, G17070, G18255, G21210 and G22247 from the NG1 sub-race; G5708, G14253, G18264 and LRK31 from the NG2 sub-race; and DRK47 and G22147 from race Peru.
Key words: andean races, water deficit, phenotypical traits, adaptation.
RESUMEN
Más del 60% de la producción mundial de fríjol común (Phaseolus vulgaris L.) crece bajo riesgo de sequía. Para identificar posibles fuentes de resistencia a la sequía en el acervo genetico andino se indagaron 64 genotipos de fríjol arbustivo de la colección de referencia del CIAT. Se evaluaron atributos fenotípicos como el rendimiento, el peso de 100 semillas (P100) y los días a la madurez fisiológica (Dam) en accesiones seleccionadas de esta colección, los cuales se sembraron en un diseño en lattice 8x8 con dos replicaciones y tres ambientes: sequía intermitente (SI) y riego (R) en Palmira y también sequía temprana (ST) en Darién, Colombia. De los genotipos evaluados, 20 pertenecen a la sub-raza Nueva Granada 1 (NG1), 19 a la sub-raza Nueva Granada 2 (NG2), 10 de la raza Perú (P), 14 son controles andinos y un control mesoamericano. Las variables se analizaron por medio de ANDEVAs combinadas entre los ambientes; además, se determinaron coeficientes de correlación simples entre el rendimiento y las demás variables evaluadas. Los genotipos con mayor adaptación a la sequía presentaron alto rendimiento, mayor P100, además de menor número de días a la madurez fisiológica. Los coeficientes de correlaciones entre el rendimiento y P100 fueron significativos y positivos, mientras que Dam presentó correlación negativa. Se identificaron 14 genotipos como tolerantes a condiciones de sequía: G4001, G5625, G6639, G16115, G17070, G18255, G21210 y G22247 de la sub-raza NG1; G5707, G14253, G18264 y LRK31 de la sub-raza NG2; y DRK47 y G22147 de la raza Perú.
Palabras clave: razas andinas, déficit hídrico, atributos fenotípicos, adaptación.
Introduction
Drought affects more than 60% of world production of common bean (Phaseolus vulgaris L.) (White and Singh, 1991), and the second cause largest to decrease in yield (Singh, 1995). In Latin America about 73% of the common bean crops grow in environments where there is often drought conditions restricting the production (Acosta- Gallegos et al., 1999). In addition, the common bean is a species very susceptible to drought compared with other legumes (Pimentel et al., 2001). The effect of drought on the common bean depends on the type (severe, moderate) and duration (early, intermittent and terminal) and the same stage of development where the cultivation is (Terán and Singh, 2002; Nielsen and Nelson, 1998).
Drought causes reduction in yield, yield components and biomass accumulation (Ramírez-Vallejo and Kelly, 1998; Nielsen and Nelson, 1998, Castañeda et al., 2006, Terán and Singh, 2006; Muñoz-Perea et al., 2006; Gómez et al., 2010). Abebe et al. (1998) found a reduction in yield of 62% in the dry environment compared with the control environment. The greater sensitivity to drought in common bean occurs during the reproductive stage, from pre-flowering to podfilling (Nielsen and Nelson, 1998; Castañeda et al., 2006).
Genotypic and phenotypic differences have been reported for drought resistance in common bean (Abebe et al., 1998), which is necessary for the selection of genotypes with adaptation to drought, is effected through a combination of criteria related to the yield, days to maturity and yield components in different moisture conditions.
Genetic improvement to get drought-resistant crops is a slow and difficult, because other factors such as diseases and heat stress affect the selection of genotypes (Subbarao et al., 1995; Beebe et al., 2008). The use of genetic variability is essential for the improvement of common bean in areas such as increased yield and tolerance to abiotic factors such as drought (Singh, 2001; Terán and Singh, 2002). To find sources of resistance to drought, consider their evolutionary origin and center of domestication understanding that improvement is the basis of genetic diversity (Terán and Singh, 2002), therefore it is important to investigate within germplasm collections common bean.
The core collection is a handy collection consists of 1440 accessions were chosen to represent the genetic diversity of the collection or global basis so they can be used in breeding programs (Brown, 1989; IPGRI, 2000). Meanwhile, the reference collection is even more representative sample of the base collection of only 200 genotypes were selected based on molecular markers to represent the greatest diversity of cultivated common bean shrub found in the core (Blair et al., 2009; CIAT, 2008). The core and reference collections represent two fairly large numbers of genotypes.
The objective of this study was to evaluate 64 genotypes shrub collection from CIAT bean reference, subject to conditions of drought stress to identify sources of resistance to drought in Andean heritage, through the characterization of phenotypic attributes.
Materials and methods
Plant material
The stock Andean bean is divided into races according to agro-morphological characteristics (Singh et al., 1991) that are reflected by polymorphism in molecular markers (Blair et al., 2007). Breeds belonging to this pool identified by Singh et al. (1991) and Blair et al. (2007, 2009), were race Nueva Granada âwhich has two sub-races, New Granada 1 (NG1) and Nueva Granada 2 (NG2)â, race Peru (P) and race Chile, although the latter did not present a clearly distinguished from the other Andean races. For this study, we used 63 genotypes of the reference collection of Andean stock and stock Mesoamerican genotype used as control. Of these 20 genotypes belonging to the Andean sub-race Nueva Granada 1 (NG1), 19 sub-race of Nueva Granada 2 (NG2), 10 race Peru (P), and 14 Andean controls (Blair et al., 2009) (Tab. 1).
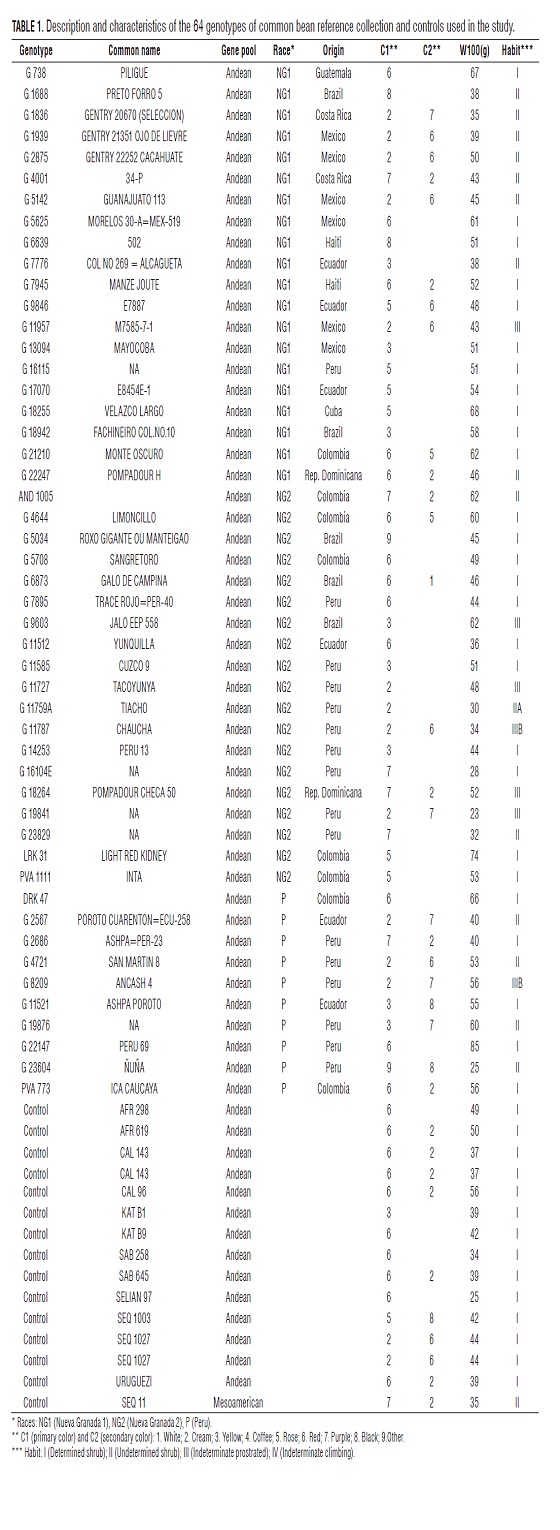
Location of trial
The study was conducted during the dry season from July to October 2008, in the towns of Palmyra and Darien, Colombia. The town of Palmira (3° 29' N, 76° 21' W) is at 965 meters and has an annual rainfall of 839 mm, with average temperature 24° C, soil Mollisol (Aquic Haplustoll), pH 7.7 and 70.5 ppm of phosphorus. The town of Darien (3° 55' N, 76° 28' W), is 1,500 meters and has an average annual rainfall of 1,650 mm, with average temperature 20 °C, Inceptisol soil (typic Dystrandept), pH 5.7 and 4.1 ppm of phosphorus which required soil fertilization with 80 kg of phosphorus.
Agricultural practices to control pests, weeds and diseases were performed manually, chemical and mechanical. the seed was treated with captan carboxin and also benomyl applications were made, and oxicarboxin inorganic sulfur to control diseases. For weed control conducted a preemergent chemical control with s-metolachlor linuron and then at 25 and 62 days after planting (Dap), control was performed with ammonium and glyphosate glufocinato. Manual-mechanical control of weeds was done at 14, 25, 38, 51 and 62 Dap. Insecticides were applied according to the degree of infestation and phenological stage for insect pest control with the active ingredients: carbaryl, milbemectin imidacloprid, deltamethrin, as recommended by the manufacturer. In addition, there were two foliar fertilizations with zinc and boron (220 g ha-1 and 400 g ha-1, respectively) at 14 and 33 Dap, to avoid possible deficiencies due to the basic pH of the Palmira soils.
Experimental design
Design was used in 8 x 8 lattice partially balanced with 2 repetitions. Environments evaluated were irrigation (R), intermittent drought (SI) and early drought (ST). The first two are located in the town of Palmyra and the latest in Darien, Colombia. For R, a total of six irrigations of 31 mm each were applied, while for SI only two irrigations were necessary to ensure the establishment of the crop. ST was not carried out in supplementary irrigation. The experimental unit consisted of two rows of 3 m in length; with a row spacing of 0.6 m planting density were 15 seeds per linear meter.
Variables evaluated
The variables evaluated were yield, number of days to physiological maturity (Dam) and weight of 100 seeds (P100). The number of days to maturity was estimated by means of daily observations by evaluating the number of days from planting until at least 50% of the experimental unit pods present a green discoloration of the characteristic color of maturity in each genotype.
To determine the weight of 100 seeds were harvested randomly five plants per experimental unit and based on the seed obtained, we calculated this variable. The whole plot was harvested, we determined the weight and seed moisture at harvest, and then make a correction to 14% moisture and express seed yield kg ha-1.
Statistical analysis
The variables studied were subjected to analysis of variance between environments combined and determined the least significant difference (LSD) between treatments. Also, the coefficients were determined simple correlations between performance and the other variables assessed. We used the GLM procedure of SAS® statistical software (version 9.1.3) (SAS Institute, 2004).
Results and discussion
In Palmira maximum and minimum temperature recorded during the crop cycle was 33°C and 16°C respectively. The potential evaporation was 416 mm, while precipitation totaled 163 mm, distributed unevenly. The data of precipitation and evaporation with rainfall distribution indicate that there was an intermittent type of stress during the period of crop growth and development (Ludlow and Muchow, 1990). Meanwhile, in Darien there was a maximum and minimum temperature of 27°C and 13°C respectively. The precipitation was 280 mm, no evaporation data were obtained but the precipitation data, along with its distribution suggest that introduced drought early in the crop because during the first trifoliate leaf stage to third trifoliate leaf rainfall was 5.3 mm (Fischer and Maurer, 1978).
Performance
The mean squares of environments, genotypes and genotype by environment interaction showed highly significant differences (P <0.001) for performance (Tab. 2). The least significant difference (LSD), allowed separating environments with differences between the environments of drought and irrigation (Tab. 3). The yield coefficient of variation was 23.1%.
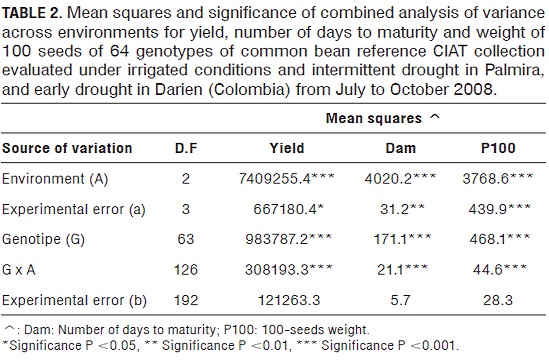
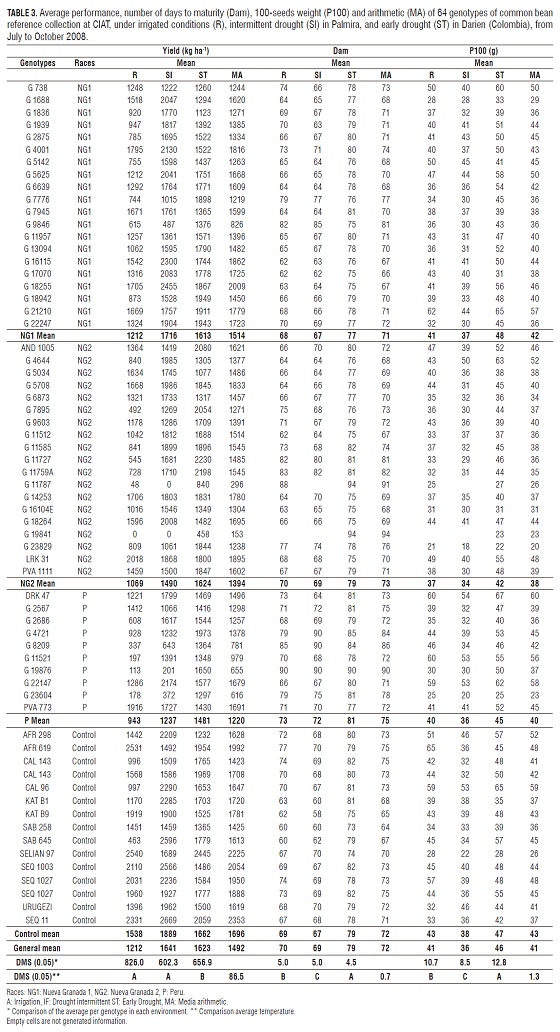
In Palmira, under the conditions of SI was the performance range of 0 - 2669 kg ha-1 with an overall average of 1,644 kg ha-1. For R the range was from 0 - 2,540 kg ha-1 with an overall average of 1,213 kg ha-1, negatively affected by the fungus Sclerotium rolfsii Sacc., which can cause considerable losses to crops in dry seasons, hot and heavy soils (Abawi, 1994). Meanwhile in ST, the performance range was 458 to 2,444 kg ha-1 with an overall average of 1,614 kg ha-1.
The controls showed the highest values for the mean arithmetic (MA) of performance defined as the average among the three environments genotype evaluation, followed by genotypes of the sub-race-group NG1. The high performance of NG1 may have been for the best adaptation of these genotypes to tropical and subtropical environments of high temperatures, while P genotypes of race had the lowest MA, possibly because of their limited adaptation to different environments where normally takes place in high altitude cold areas (Singh et al. 1991).
By yield, the 64 genotypes could be classified into four groups. The first group corresponds to the genotypes that had high performance in the three environments, such as controls SEQ11, SEQ1027, together with the genotypes G18255, G16115, G21210, G22247, G17070, G5625, G6639 and G4001 (in the sub-race NG1) LRK31, G5708, G14252 and G 18 264 (sub-race of the NG2) and DRK47 and G22147 (breed P) (Tab. 3). The good fit of the controls may have been caused by being improved genotypes to withstand drought conditions while the accessions may be due to race are mainly from New Granada which is adapted to dry conditions and higher temperatures (Singh et al., 1991).
In the second group are low-yielding genotypes in three environments: G11787, G19841 (sub-race of the NG2), G23604 and G8209 (breed P) (Tab. 3), ie the lowest degree of adaptation to the environments tested. In the third group are the genotypes that responded to the drought conditions but had low performance in R as the control and accessions SAB645 G11585, G5625, G11512, and G11759A (sub-race of the NG2) (Tab. 3), favoring dry conditions return. In the latter group, the genotypes are presented in irrigation performance, but with below average performance in drought, such as G7945, G1688 (the sub-race NG1), G14253, G5034, G18264 (sub-race of the NG2) and G2567 (breed P), which had little or no adaptation to drought, that is they are susceptible genotypes. Muñoz-Perea et al. (2006), performed a similar grouping according to the degree of adaptation of common bean to drought conditions. Only considered genotypes adapted to this condition and the division between these genotypes was based on the yield in nonstress conditions, whereas in our study were considered the multiple responses of genotypes to the environments evaluated for grouping.
The performance is the best selection criteria used to determine the drought resistance of a genotype (Terán and Singh, 2002). White and Singh (1991) and Abebe et al. (1997), report the use of MA as a criterion to identify genotypes with high yield under drought conditions. Based on the foregoing, the genotypes that are mentioned in the first group would be selected for the possible onset of breeding programs, but according to Beebe et al. (2008), now other attributes are also considered varietal selection of sources of resistance to drought and ability to mobilize photosynthates. Unfortunately the irrigation, could not be used as a control environment, for which he was regarded as an evaluation environment. The high temperatures that occurred in Palmira during the growing season caused a heat stress. Root growth, reduces the size of the seed and the abortion of flowers and pods were affected for stress (Shonnard and Gepts, 1994; Rao, 2001). Besides this, the fungus Sclerotium rolfssi Sacc, affected the performance in this environment; the attack of this fungus was favored by the environmental conditions of R, since temperature and high relative humidity, followed by dry periods favor the establishment and disease development (Abawi, 1994; Blum-B. et al., 2003).
Number of days to physiological maturity
The mean square of the number of days to maturity (Dam) were highly significant between environments, genotypes and genotype by environment interaction (Tab. 2) and comparing the averages using the DMS environments, each environment formed a distinct group (Tab. 3), indicating that this variable was affected by the concentration of water in the soil. The coefficient of variation was low with a value of 3.4%.
In R, Dam was the range of 60 to 90 d, with an average of 70 d. Whereas in SI, the range was from 58 to 90 d, with an average of 69 d and finally in ST the range was from 73 to 94 days with an average of 79 d. There was a reduction in the average of 1 d (1.5%) compared IS and R. Also there was a reduction in the average of 10 d (13%) in SI compared with ST in contrast there was an increase in the average of 10 d (11%) in Dam when comparing ST with R being in different locations. Terán and Singh (2002), in Mesoamerican heritage accessions and breeding lines, found a 3% decrease in the number of days to maturity between irrigation environments and drought in Palmira attributed to high temperatures that occurred on the site of study.
Singh (1995), says that drought accelerates the maturity as a mechanism of resistance to drought escape, especially when stress increases after flowering. Singh (2007) reported a reduction of up to 4 d, in three accessions and 17 cultivars of race Durango. Contrary to the above, Muñoz-Perea et al. (2006) reported an increase in the number of days maturity intermittent drought conditions Creole cultivars and genotypes Kimberly Mesoamerican heritage, USA, supported in that this type of stress, plants exhibit a disruption of metabolic processes before recovering step of modifying the reproductive vegetative state. The Dam increased from ST and R, is given by the environmental conditions at Darien, as a lower temperature reduces the metabolic activity of the plant, resulting in a longer duration of phenological stages (Lambers et al., 2000).
In general, the genotypes of the sub-race reached physiological maturity NG1 fastest followed by the controls (Tab. 3), explained why the race Nueva Granada as reported by Singh et al. (1991), is the race earlier in the Andean stock, while the genotypes of P is the race that the longer it takes to mature.
Genotypes had lower number of days to maturity and performance in the three environments were: G18255, G17070, G16115, G1688, G5625 and G6639 (in the subrace NG1) LRK31, G11512, G14253 and G18264 (from the sub NG2-race), G22147 (breed P) and controls KATB9, KATB1, SAB645, and SEQ11 SEQ1003. Meanwhile, G19876, G23604, G8209 (breed P), G7677 (the sub-race NG1), G19841 and G11787 (sub-race of the NG2) showed the highest Dam accompanied by low yields in some cases without forming pods, showing its poor adaptation to the environment assessment. Muñoz-Perea et al. (2006), claim that precocious cultivars have a lower net water requirement throughout the entire life cycle, compared with those who mature later, this means a reduction in the effect of drought on the crop. Studies White and Singh (1991) and Beebe et al. (2008) reported that late maturing lines, suffer a greater decrease in performance under drought stress. According to Acosta-Diaz et al. (2004), lower high-performance Dam in drought conditions is associated with a high capacity to mobilize photosynthates to pods.
Weight of 100 seeds
The mean square of the 100-seeds weight (P100) was highly significant between environments, genotypes, and significant genotype by environment interaction (Tab. 2). Comparing the averages for the environment and DMS, found that each environment P100 formed in a different group (Tab. 3), indicating a direct effect of moisture content on this variable. The coefficient of variation was 13.2%; usually, this is less than the coefficients of variation of other components of performance due to its higher heritability.
In R the range of P100 was 21 to 65 g, with an average of 41 g, whereas in SI the range was from 18 to 54 g, with an average of 36 g and, finally ST range was from 22 to 67 g, with an average of 46 g (Tab. 3). There was a reduction in the average weight of 100 seeds of 5 g (11%) in SI compared with R and 10 g (22%) in SI compared with ST. Singh (2007), reported a reduction in 14% P100 genotypes of the Mesoamerican heritage, under conditions of moderate type drought. Muñoz-Perea et al. (2006) found a 22% reduction in the P100 in 13 cultivars and three accessions of Mesoamerican heritage under intermittent drought conditions and terminal. The reduction in P10 is caused by a decrease in photoassimilate and water that goes to the seed filling period, the effect of drought (Muñoz-Perea et al., 2006). The controls had higher P100 in MA with 43 g, followed by NG1 with 42g, while P showed P100 race with 38 g lower in MA (Tab. 3).
This feature made a big difference between the sites of study, so their results are presented separately. Genotypes showed the highest P100 high performance under SI and R in Palmira, were: G21210, G18255, G17070, G4001, G7945, G16115, G22247, G5625, G6639 (the sub-race NG1) LRK31, G5708, G4644, G14253 and G18264 (sub-race of the NG2), and G22147 DRK47 (breed P) and controls AFR619, SEQ1003, SEQ1027, AFR298 and KATB9. SEQ11 SELIAN97 and controls showed low levels of P100 but had higher yields in R and SI respectively. In ST (Darien) most P100 genotypes and high performance were: G21210, G18255, G5625, G6639, G1842 (the sub-race NG1), AND1005, G11727, G14254 (sub-race of the NG2), G4721 of (Race P) and controls SAB645, and SEQ1003 CAL143. According to Gebeyehu (2006), the resistant genotypes have a strong relationship and good translocation landfill source of carbohydrates that allows them to maintain high values of P100 regardless of moisture conditions.
Correlations between performance and phenotypic variables
Was presented positive correlations and highly significant between yields in the three environments (Tab. 4). This result indicates that genotypes with high performance in SI, probably had a high-performance R and ST. This result accords with what was suggested by Schneider et al. (1997), where the selection of genotypes with resistance to drought, is equally effective under different stress levels. In the study, the highest correlation was obtained between the environments of Palmira, as would be expected to be made in the same locality.
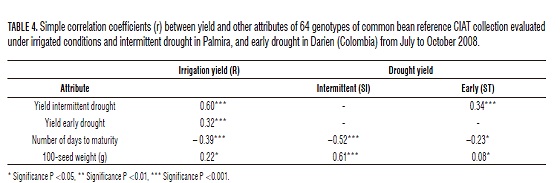
There was a negative correlation between yield and highly significant Dam IS and R, and significant ST (Tab. 4). This result agrees with that presented by Abebe et al. (1998) and Singh (1995), where the genotypes had lower Dam under drought, reached the highest yields. Muñoz-Perea et al. (2006), say a quick physiological maturity intermittent drought is associated with the escape mechanism.
The correlation between performance and P100 was positive, being highly significant in itself, significant R and without significance in ST. This compares with Muñoz- Perea et al. (2006), who report a significant positive correlation but intermittent drought conditions. Terán and Singh (2002) found a negative correlation under drought and irrigated in Mesoamerican heritage accessions and breeding lines from crosses race, this supported where performance is a function of the number of seed and plant to increase its number compensation ago, reducing the size of the seed. By contrast, in this study the genotypes had a larger P100 of seed were generally indicating high performance in the Andean heritage works a different mechanism for obtaining high performance.
Conclusions
The field evaluation of the 63 genotypes of the reference collection at CIAT, identified 14 genotypes that showed a superior adaptation to stress conditions caused by drought.
The materials G18255, G17070, G16115, G21210, G4001, G5625, G6639 and G22247 sub-race of the New Granada 1, LRK31, G14253, G18264 and G5708 of the sub-race Nueva Granada genotypes 2 and G22147 DRK47 of race and Peru, were the most prominent. This result suggests that resistance to drought conditions is not limited to one race or group, which should further investigate the different inputs germplasm Andes.
The adaptation of these genotypes to stress conditions and good response in irrigated conditions were associated with fewer days physiological maturity, high yield and weight of 100 seeds.
By the other hand, the results for the weight of 100 seeds, suggest that the body of an Andean drought tolerance mechanism could be the good grain filling.
In some genotypes, the phenotypic plasticity allowed a better adaptation to environmental conditions by reducing the effects of drought on bean plants.
As presented in the study, it is important to use different environments for evaluation and determination of the different phenotypic attributes for the selection of genotypes resistant to drought conditions, optimizing the selection process.
Acknowledgements
The authors wish to thank Idupulapati Rao, Ph.D., José Polania, IA and Jaime Ricaurte, IA for the advice to the research project. In addition to giving recognition to the valuable assistance of field staff and Andean bean physiology at CIAT. This work was funded GCP/TL1 project.
Literature cited
Abawi, G.S. 1994. Pudriciones radicales. pp. 153-158. En: Pastor- Corrales, M. y M. Schwartz (eds.). Problemas de producción de fríjol en los trópicos. 2nd ed. CIAT, Cali (Colombia). [ Links ]
Abebe, A., M.A. Brick y R.A. Kirkby. 1998. Comparison of selection indices to identify productive beans lines under diverse environmental conditions. Field Crops Res. 58, 15-23. [ Links ]
Acosta-Díaz, E., C. Trejo-López, L. Ruiz-Posadas, J.S. Padilla- Ramírez and J.A. Acosta-Gallegos. 2004. Adaptación de fríjol común a sequía en la etapa reproductiva. Terra Latinoamericana 22, 49-58. [ Links ]
Acosta-Gallegos, J.A y J.W. White. 1995. Phenological plasticity as an adaptation by common bean to rainfed environments. Crop Sci. 35, 199-204. [ Links ]
Acosta-Gallegos, J.A., E. Acosta, S. Padilla, A. Goytia, R. Rosales and E. Lopez. 1999. Mejoramiento de la resistencia a la sequía del fríjol común en México. Agron. Mesoam. 10, 83-90. [ Links ]
Beebe, B., J. Rengifo, E. Gaitan, M.C. Duque and J. Thomes. 2001. Diversity and origin of Andean landraces of common bean. Crop. Sci. 41, 854-862. [ Links ]
Beebe, S.E., I.M. Rao, C. Cajiao and M. Grajales. 2008. Selection for drought resistance in common bean also improves yield in phosphorus limited and favorable environments. Crop Sci. 48, 582-592. [ Links ]
Blair, M.W., J.M. Diaz, R. Hidalgo, L.M. Diaz y M.C. Duque. 2007. Microsatellite characterization of Andean races of common bean (Phaseolus vulgaris L.).Theor Appl Genet. 116, 29-43. [ Links ]
Blair, M.W., L.M. Diaz, H.F. Buendía y M.C. Duque. 2009. Genetic diversity, seed size associations and population structure of core collection of common beans (Phaseolus vulgaris L.). Theor. Appl. Genet. (On line). [ Links ]
Blum-B, L.E., C.V.T. Amarante, C.J. Arioli, L.S. Guimaraes, A. Dezanet, P.H. Neto and F.R. Scheidt. 2003. Recão de genótipos de Phaseolus vulgaris à podriadão do colo e ao Oídio. Fitopatol. Bras. 28(1), 96-100. [ Links ]
Brown, A. 1989. Core collections: a practical approach to genetic resources management. Genome 31, 818-824. [ Links ]
Castañeda, M.A., L. Córdova, V. Gonzales, A. Delgado, A. Santacruz, and G. García. 2006. Respuestas fisiológicas, rendimiento y calidad de semilla en fríjol sometido a condiciones de estrés hídrico. INCI 31(6), 461-466. [ Links ]
CIAT - Centro Internacional de Agricultura Tropical. 2008. Bean Project Annual Report 2007-2008, CIAT. Cali, Colombia. Pp. 55-56. [ Links ]
Fischer, R.A. and R. Maurer. 1978. Drought resistance in spring wheat cultivar I. Grain yield responses. Augt. J. Agr. Res. 29, 897-912. [ Links ]
Gebeyehu. S. 2006. Physiological response to drought stress of common bean (Phaseolus vulgaris L.) genotypes differing in drought resistance. Tesis de doctorado. Universidad de Liebig- Giessen, Giessen (Germany). [ Links ]
Gomez, A., H. Rojas, F.A. Vallejo and E.I. Estrada. 2010. Determinación del requerimiento hídrico del pimentón en el municipio de Candelaria, departamento del Valle del Cauca. Acta Agronómica 59(4), 442-448. [ Links ]
IPGRI. 2000. Core collections of plant genetic resources, IPGRI. Rome, Italy. [ Links ]
Lambers, H., F.S. Chapin and T.L. Pons. 2000. Chapter 7: Growth and allocation. pp. 323. In: Plant physiological ecology. Springer- Verlag, New York. [ Links ]
Muñoz-Perea, C.G., H. Terán, R. Allen, J.L. Wright, D.T. Westermann and S.P. Singh. 2006. Selection for drought resistance in dry bean landraces and cultivar. Crop Sci. 46. 2111-2120. [ Links ]
Muñoz-Perea, C.G., R.G. Allen, D.T. Westermann, J.L. Wright y P.S. Singh. 2007. Water use efficiency among dry bean landraces and cultivars in drought-stressed and non-stressed environments. Euphytica 155, 393-402. [ Links ]
Nielsen, D.C. y N. Nelson. 1998. Black bean sensitivity to water stress at various growth stages. Crop Sci. 38, 422-427. [ Links ]
Pimentel, C., A.A. Gomes, and T.B. Ottoni Filho. 2001. Trocas gaseosas em folhas de feijoeiro de plantas hidratadas e teores de carbohidratos e aminoácidos livres sob deficiência hídrica. Agronomia 35, 66-70. [ Links ]
Ramírez-Vallejo, P. and J.D. Kelly. 1998. Traits related to drought resistance in common bean. Euphytica 99, 127-136. [ Links ]
Rao, I. M. 2001. Role of physiology in improving crop adaptation to abiotic stresses in the tropics: the case of common bean and tropical forages. Pp. 583-613. En: M. Pessarakli (ed.). Handbook of plant and crop physiology. 2a ed., Marcel Dekker, Inc., New York. [ Links ]
SAS Institute Inc. 2004. SAS/STAT(tm) 9.1 User's guide. SAS Institute Inc., Cary NC, USA. [ Links ]
Schneider, K.A., R. Rosales-Serna, F. Ibarra-Pérez, B. Cáceres- Enrique, J.A. Acosta-Gallegos, P. Ramírez-Vallejo, N. Wassimi, J.D. Kelly. 1997. Improving common bean performance under drought stress. Crop Sci. 37, 43-50. [ Links ]
Shonnard, G.C. and P. Gepts. Genetic of heat tolerance during reproductive development in common bean. 1994. Crop. Sci. 34, 1168-1175. [ Links ]
Singh, S.P., P. Gepts and D.G. Debouck. 1991. Races of common bean (Phaseolus vulgaris, Fabaceae). Econom. Bot. 45(3), 379-96. [ Links ]
Singh, S.P. 1995. Selection for water-stress tolerance in interracial population of common bean. Crop Sci. 35, 153-165. [ Links ]
Singh, S.P. 2001. Broadening the genetic base of common bean cultivars: a review. Crop Sci. 41, 1659-1675. [ Links ]
Singh, S.P. 2007. Drought resistance in the race Durango dry bean landraces and cultivars. Agron. J. 99, 1219-1225. [ Links ]
Subbarao, G.V., C. Johansen, A.E. Slinkhard, R.C. Nageswara Rao, N.P. Saxena and Y.S. Chauchan. 1995. Strategies for improving drought resistance in grain legumes. Crit Rev Plant Sci. 14, 469-523. [ Links ]
Terán, H. and S.P. Singh. 2002. Comparison of sources and lines selected for drought resistance in common bean. Crop Sci. 41, 64-70. [ Links ]
White, J.W., and S.P. Singh. 1991. Breeding for adaptation to drought. pp. 501-560. In: A. van Shoonhoven and O. Voysest (eds.). Common beans: research for crop improvement. CABI, Wallingford (UK) and CIAT, Cali (Colombia).
[ Links ]
