










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.29 no.2 Bogotá June/Aug. 2011
PLANT BREEDING, GENETIC RESOURCES & MOLECULAR BIOLOGY
Behavior of introduced regional clones of Theobroma cacao toward the infection Moniliophthora roreri in three different regions of Colombia
Comportamiento de clones regionales e introducidos de Theobroma cacao ante la infección de Moniliophthora roreri en tres regiones diferentes de Colombia
Yeirme Jaimes1,3, Fabio Aránzazu2, Eleonora Rodríguez1, Nubia Martínez2
1Experimental Station La Suiza, Corporación Colombiana de Investigación Agropecuaria (Corpoica). Rionegro (Colombia).2Research Program; Federación Nacional de Cacaoteros-Fedecacao. San Vicente (Colombia).
3Corresponding author. yjaimes@corpoica.org.co Recceived for publication: 23 February, 2010. Accepted for publication: 2 June, 2011.
ABSTRACT
Moniliasis (frosty pod rot), caused by Moniliophthora roreri, is the most limiting disease of cacao bean production in Colombia. Disease control has focused primarily on the implementation of cultural methods, which are inefficient and increase costs of production. A promising alternative for the control of M. roreri is the genetic approach, involving the search for plants with known resistance in the field. The aim of this study was to evaluate the behavior of 13 regional clones of Corpoica Selection Colombia (SCC), ten regional Fedecacao clones (with the initial F), three clones Caucasia (CAU) and 13 introduced clones for resistance to M. roreri, by monitoring symptoms and signs of the pathogen in fruits of the cacao at three experimental sites located in Arauquita (Arauca), San Vicente and Ríonegro (Santander). To estimate the degree of susceptibility or resistance of each clone, the pathogen was inoculated into fruits 75±5 days old. Eight weeks later, the variables Incidence, Internal Severity Index and External Severity Index were evaluated. In the three experimental sites, the ICS clone 95 behaved as a resistant plant with ISI ranging from 1.4 to 1.9. In the other clones tested, susceptibility varied according to altitude and environmental conditions. However, the ICS clone 95 showed partial resistance, varying the spread of the pathogen in the plant tissue.
Key words: moniliasis, corn, cacao, clones, resistance.
RESUMEN
La moniliasis, causada por Moniliophthora roreri, es la enfermedad más limitante de la producción de granos de cacao en Colombia. El control de la enfermedad se ha enfocado principalmente en la implementación de métodos culturales, que resultan ineficientes y elevan los costos de producción. Una alternativa promisoria para el control de M. roreri es el método genético, involucrando la búsqueda de materiales con reconocida resistencia a nivel de campo. El objetivo del presente trabajo fue evaluar el comportamiento de 13 clones regionales Selección Colombia Corpoica (SCC), diez clones regionales Fedecacao (con la inicial F), tres clones Caucasia (CAU) y 13 clones introducidos por resistencia a M. roreri, mediante el seguimiento de síntomas y signos del patógeno sobre frutos de cacao bajo condiciones de tres sitios experimentales ubicados en Arauquita (Arauca), San Vicente y Ríonegro (Santander). Para estimar el grado de susceptibilidad o resistencia de cada clon, se inoculó el patógeno sobre frutos de 75 ± 5 días de edad. Ocho semanas después, se registraron las variables de Incidencia, Índice de Severidad Externa e Índice de Severidad Interna. En los tres sitios experimentales, el clon ICS 95 se comportó como un material resistente, con ISI que varió entre 1,4 a 1,9. En los demás materiales evaluados, la susceptibilidad varió de acuerdo con la altitud y las condiciones medio ambientales. Sin embargo, el clon ICS 95 presentó una resistencia parcial, variando el avance del patógeno en el tejido de la planta.
Palabras clave: moniliasis, mazorca, cacao, clones, resistencia.
Introduction
Cacao (Theobroma cacao) is one of the most important tropical crops: economically, socially and environmentally (Hebbar, 2007). For 2010, FAO reported world production of 4 million t of dry grain, of which over 60% was used in manufacturing in the chocolate industry (FAO, 2011). However, this crop is attacked by a large number of pathogens, which are responsible for up to 30% of global production losses, including Moniliophthora roreri H.C. Evans, Stalpers, Samson & Benny; Moniliophthora perniciosa (Stahel) Aime & Phillips-Mora, comb. nov. and Phytophthora sp. as the most important (Hebbar, 2007). Moniliasis, the common name for the disease caused by M. roreri, is considered a highly destructive disease (Evans, 2007), with its introduction to the African continent considered a major threat to the cacao economy (Ploetz, 2007).
This pathogen is limited to Spanish-speaking countries (Evans, 2007). Farmers recognize M. roreri primarily by external symptoms on the fruits of cacao, especially by the appearance of signs of the pathogen, such as white mycelium, ashen in appearance, or the complete sporulation of the pathogen on the affected fruit tissue (Phillips-Mora and Wilkinson, 2007). The origin of the disease comes from the northeastern Colombian region where the domestication of cacao involved the association of T. cocoa with other related species such as Theobroma gireli for thousands of years. Hence, at present, the genus Theobroma, like Herrania, is host to M. roreri (Evans, 2007).
In Colombia, moniliasis can cause losses from 40 to 100% of the dry grain, depending on the farmers' practices (Castellanos et al., 2007). Control of M. roreri is mainly based on cultural practices, but this must be supplemented with other control methods because the pathogen inoculum level and aggressiveness, especially in the origin region (Aime and Phillips-Mora et al., 2005), mean that these practices are inefficient. Other studied control methods, rare in Colombia, are biological and chemical control. However, the adoption of these practices or methods is difficult due to the low price of dry grain nationwide. As an alternative to this problem and complementary to other methods, genetic control is promising for the management of the disease (Phillips-Mora et al., 2005), considering that the selection of resistant plants could provide a control method with a longer duration and lower costs, which together with the implementation of other methods would result in the successful control of the disease .
The nature of disease resistance of any plant is present under the cultivation conditions. This resistance can occur as a survival response of the plants to parasitic organisms, involving a wide range of defense mechanisms that stop or prevent the attack of the pathogen. The selection of resistant materials involves measuring the degree of susceptibility to the pathogen; this requires evaluating the direct and indirect effects of the pathogen on the host plant (Ribeiro et al., 2001). According to the above, the objective of this study was to evaluate the behavior of some local plants as well as introduced ones against infection by M. roreri, by monitoring symptoms and signs of the pathogen on fruits of each cacao clone under field conditions.
Materials and methods
Location of experimental sites
Two experimental sites were selected on the Mountain Santandereana and a place in the Llanos Foothills in the department of Arauca. These sites were: Experimental Station La Suiza in Rionegro (Santander, Colombia) Corpoica located at 7° 22' N; 78° 10' W, at an altitude of 500 m, average temperature of 28°C, average annual precipitation of 1,800 mm and average relative humidity of 85%; Villa Monica Farm Fedecacao in San Vicente de Chucuri (Santander, Colombia) located at an altitude of 850 m, with an average temperature of 24°C, average annual precipitation of 1,800 mm and average relative humidity of 82% and Santa Elena Farm Fedecacao in Arauquita (Arauca, Colombia) located at 7° 60' N; 71° 61' W, at an altitude of 162 m, average temperature of 28°C, average annual precipitation of 2,500 mm and average relative humidity of 75%.
Plant material
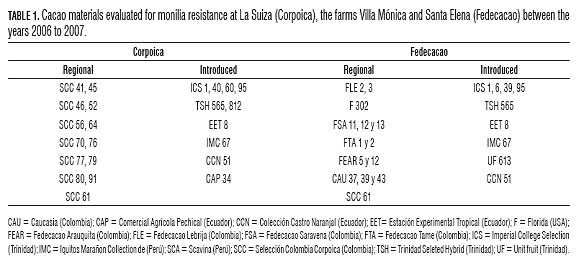
Regional and introduced materials were selected taking into account those most widely used commercially and their availability in each of the three clonal gardens selected as the experimental sites (Tab.1) .
The fruits were obtained by hand pollination of flowers of each material (clone mother) and with clone IMC 67 (clone father). The flowers of each material were covered with high-density polyethylene bags to protect the environment from the pathogen (Phillips-Mora et al., 2005).
Obtaining pathogen inoculum
The pathogen inoculum was obtained from a collection of diseased fruits with symptomatic stains. These were subjected to disinfection with sodium hypochlorite 1%, with subsequent washing with distilled water. The fruits were placed in a humid chamber with environmental conditions equal to those of the test sites for 15 d. After this period, the pathogen spores were obtained by scraping the sporulated fruit.
Evaluation of cacao clones resistance
Resistance assessments were made on fruits of the cacao at 75±5 d old. For inoculation of the pathogen, dry spores were taken with the tip of an entomological pin, taking approximately 3.4·104 spores/cm2 (Merchán and Restrepo, 1980). The dry spores were dispersed over an area of 4 cm2 on the fruit, moistened with sterile distilled water. The inoculated fruits were covered with high-density perforated bags and moistened absorbent paper for 5 d to create favorable conditions for the onset of the infection process.
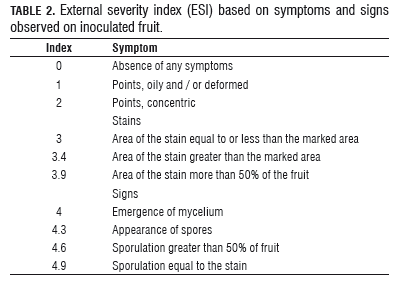
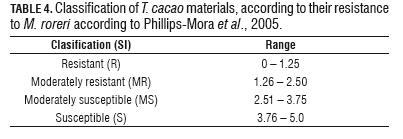
After 8 weeks, the progress of the disease was assessed. For this, the variables recorded were: incidence (I) (% of diseased fruits) external severity index (ESI), using the modified severity scale of five degrees of Sánchez and González (1989) (Tab.2), and internal severity index (ISI) (Tab.3). The classification ISI of the cacao material was based on the scale proposed by Phillips-Mora et al. (2005) (Tab.4).

The materials selected at La Suiza in Corpoica were evaluated twice, once in 2006 and again in 2007. The selected materials in Fedecacao were evaluated three times in each location, once per semester in 2006 and the second half of 2007. We used a completely randomized design, where each clone had a treatment. In each test, we used a sample of 10-15 fruits; each fruit was considered an experimental unit. Each test was considered a repetition over time.
Statistical Analysis
For the statistical analysis, the ISI and ESI variables were analyzed for each region separately, submitting the data for each site for analysis of variance and comparison of means by the Tukey method (P=0.05). The data analysis was performed using SAS software version 8 (SAS Institute, Cary, NC) .
Results
Behavior of materials in the Llanos Foothills At the Santa Elena Farm tested clones had an incidence higher than 76% (Tab.5), with the exception of clones Caucasia, CAU 51 and 43, which had an incidence of 45 and 53% respectively .
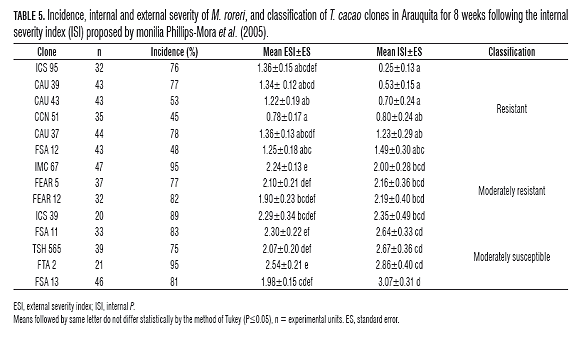
In assessing the severity of the disease internally, ICS materials 95, CAU 39, CAU 43, 51 and CAU 37 behaved as resistant (R) (Tab.5). The Clone of note, the ICS 95 (clone introduced) had the lowest ISI, which was 0.25±0.13, with an average damage of 25%. As for the clones classified as MR, only clone FSA 12 (regional material) presented an ISI with no significant difference with respect to materials R, showing an average internal damage of 29.5%. The other materials showed internal damage exceeding 40%. In assessing the severity of the disease outside, it was observed that clone CCN 51 had the lowest ESI average value, this being 0.78 with the first symptoms of the disease. (Tab.5), followed in ascending order material from the clones, CAU 43, FSA 12, CAU 39, ICS 95 and CAU 37, that presented initial symptoms, such as oily spots, humps or concentric points (Phillips-Mora et al., 2005).
Behavior of Theobroma cacao at Santandereana Mountain
Cacao materials were evaluated under two environmental conditions in the Santandereana mountains on the Villa Monica Farm, San Vicente de Chucurí and La Suiza.
Villa Monica Farm
On the Villa Monica Farm, there was a higher incidence, up to 81%, in most of the materials tested, with the noted exceptions of clone CAU 43, CCN 51 and CAU 39 with an incidence of 39, 61 and 69%, respectively (Tab.6). Regarding the assessment of the severity of the disease, only clone ICS 95 was classified as a durable material with an internal severity index average of 0.82, i.e. an internal damage average of 16.4%. Among the materials evaluated, the following were notable for their behavior as moderately resistant (MR) FLE regional clones FLE 2 and CAU 43 with ISI CAU 1.63 and 1.79 1.63, respectively (Tab.6). The other materials showed damage 43.8% higher, i.e. an average ISI less than 2.19 (Tab.6). In the external assessment of severity, the notable ones were clones CAU 43, ICS 95 and FLE 2 with ESI averages of 1.6, 1.9 and 1.63 (Tab.6), i.e. only initial symptoms presented on these materials (Phillips-Mora et al., 2005).
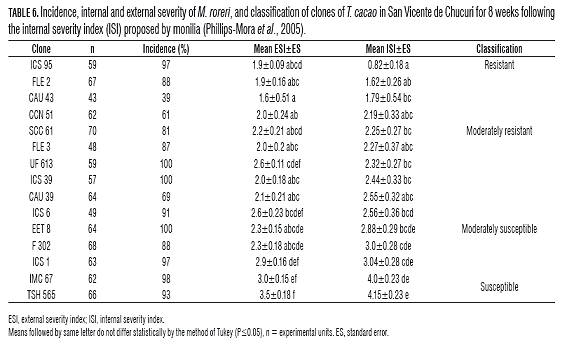
Experimental Station La Suiza
In the E.E. La Suiza, were presented an incidence of 100% in all tested materials (Tabs. 7 and 8). Regarding the severity of the disease in the introduced clones, as at the Villa Monica farm, only clone ICS 95 behaved as a resistant material, with an average ISI of 1.4 (Tab.7), indicating an average internal damage of 16%. The other introduced materials behaved from moderately susceptible to susceptible, with internal damage above 58% (Tab.7). In the external assessment of severity, only clone ICS 95 presented an ESI mean of 1.4, equivalent to the onset of initial symptoms (Tab.7) (Phillips-Mora et al., 2005).
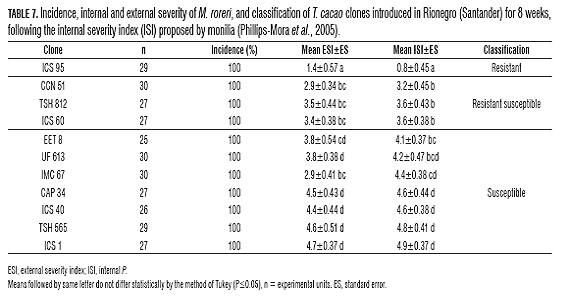
In the evaluation of the regional clones, the ranking ranged from moderately susceptible (MS) to susceptible, presenting an ISI above 2.5, i.e. internal damage exceeding 50% (Tab.8), including the clone SSC 61, which presented an average ISI of 4.8, i.e. an internal damage average of 96%. In most of the clones, a higher than average ESI, 2.5 (Tab.8), indicating that most of the materials developed stain symptoms, especially in clones SCC 45 and 70, which completed the disease cycle, i.e. until sporulation (Tab.8).
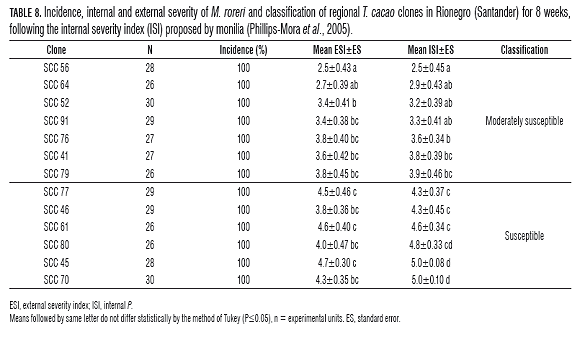
Discussion
In this study, only clone ICS 95 behaved as a resistant material with respect to the other clones evaluated, with internal damage below 20% (Fig.1), coinciding with that reported by other authors (Tab.9) (Aránzazu, 2003; Phillips-Mora et al., 2005; Rodríguez and Medina, 2005). The same results found by Phillips-Mora et al. (2005) evaluated this material against seven strains of M. roreri, noting that the material retained its behavior as resistant material. Other materials that showed partial resistance under environmental conditions at the Santa Elena and Villa Monica were the clones, CCN 51 (Fig.1), CAU 37, CAU39, CAU 43 and IMC 67 (Fig.1) (Tab.9). These materials had atypical symptoms, such as subsidence, green ripening, wilting in the affected area and the formation of green islands on senescent plant material (Fig.2A). In the specific case of the green islands, this is because of the pathogens hemibiotrophics such as M. roreri that invade living cells, reverse the metabolismimpleof the host to favor their growth and reproduction. That is when the green islands form on the senescent plant material, in order to ensure their survival (Hammond-Kosack and Jones, 1997).

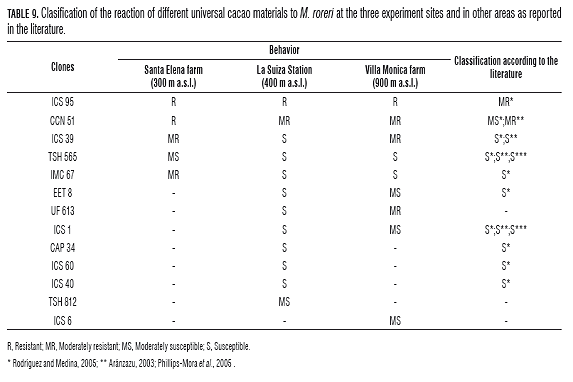

In the specific case of a limited area or minor external damage to the fruit skin, reduction of the amount of inoculum in the environment is guaranteed. However, estimating the amount of affected tissue is generally a good estimate of the amount of the pathogen. But the amount of the pathogen is not just dependent on the level of cultivar resistance, since there are other factors such as interference between plots, the relationship between symptoms of the disease, the number and density of inoculum of the pathogen, and precocity and habitat of the host (Ribeiro et al., 2001). This could elucidate the ICS 95 clone, where this material under the conditions of the E.E. La suiza showed a small affected area, but with full internal tissue damage (Fig.2) in contrast to that observed at the Villa Monica farm and Santa Elena (Fig.2).
According to the above, one can conclude that clone ICS 95 has partial resistance, where the expression of resistance genes can be modified by the action of other genes (epitasis), the state of development of plant tissue or the environment. In the atmosphere, the temperature plays an important role, which has been reported by several authors, where the expression of certain genes depends on the temperature at which they are exposed. This is the case of genes L1, L3, L7, L8, L10 and L 11 in flax for the rust Melampsora lini (Islam and Mayo, 1990), Sr6 in oats for stem rust, LR6 and Lr17 in oats for leaf rust (Browder, 1985), among many other reported genes (Ribeiro et al., 2001).
Significantly, the behavior of clone CCN 51, which is the product of crossing ICS 95 X BMI 67, can show partial resistance to M. roreri as an inherited characteristic. In the case of clone ICS 95, Johnson et al. (2009) highlighted in their study the behavior of these materials, and their source of resistance displayed on TSH materials. This shows that resistance is an inherited trait associated with resistance.
Although partial resistance was observed in clone ICS 95 at the three experimental sites (Tab.9), and to a lesser extent in CCN51 and CAU materials under the conditions of the Villa Monica farm and Santa Elena (Tab.9), the implementation of a process of improvement by conventional methods is a slow process. Furthermore, although there is much genetic variability of clonal material, the majority of improvement projects have used a reduced genetic base (Motamayor et al., 2002; Kuhn et al., 2003). This is why the selection of new materials with resistance to monilia with current methodologies is a challenge. This is because field trials are expensive and take more than 5 years to find results (Kuhn et al., 2003).
In the specific case of clone ICS 95, as a self-compatible material, it could be integrated into a genetic enhancement program by selecting filial generation 2, i.e. F2 the population, allowing the purification of the genes involved in the resistance of the materials. However, keep in mind that the disease resistance of T. cocoa is multigenic, and the associated phenotypic resistance is currently unknown, it is necessary to integrate molecular tools that reduce the cycle of selection in clonal materials with complete resistance to M. roreri (Kuhn et al., 2003).
According to the above, it is important to focus research on the generation of materials resistant to M. roreri in three aspects: (1) Identification of genes or alleles responsible for partial resistance of clone ICS 95 and (2) Searching for new sources of resistance genes in hybrids that would expand the genetic base for resistance sources to moniliasis.
This, given the current trend to expand planting areas, it is a goal both globally and for Colombia as a control measure using genetic improvement programs for the control of disease in cacao producing countries. Incorporation of resistant materials into production systems will result in an environmentally friendly and cost effective alternative, thus allowing the reduction of the spread of M. roreri (Johnson et al., 2009), which is important because the disease is currently invasively and rapidly spreading throughout Mesoamerica.
Literature cited
Aime, M. and W. Phillips-Mora. 2005. The causal agents of witches' broom and frosty pod rot of cacao (chocolate, Theobroma cacao) form a new lineage of Marasmiaceae. Mycología 97, 1012-1022. [ Links ]
Aránzazu, F. 2003. Evaluación de híbridos y clones de cacao en rendimiento y calidad para la zona marginal baja cafetera. Corporación Colombiana de Investigación Agropecuaria (Corpoica); Luker, Manizales, Colombia. [ Links ]
Browder, L. 1985. Parasite: host: environment specificity in the cereal rusts. Annu. Rev. Phytopathol. 23, 201-222. [ Links ]
Castellanos, O., L. Torres, S. Fonseca, V. Montanez, and A. Sánchez. 2007. Agenda prospectiva de investigación y desarrollo tecnológico para la cadena productiva de cacao-chocolate en Colombia. Grupo de Investigación y Desarrollo en Gestión, Productividad y Competitividad - Biogestión, Universidad Nacional de Colombia; Ministerio de Agricultura y Desarrollo Rural - MADR, Bogota. [ Links ]
Evans, H. 2007. Cacao diseases - the trilogy revisited. Phytopathol. 97, 1640-1643. [ Links ]
FAO, Food and Agriculture Organization of the United Nations. 2010. Faostat. In: http://faostat.fao.org; consulted: June, 2011. [ Links ]
Hammond-Kosack, K. and J. Jones. 1997. Plant disease resistance genes. Annu. Rev. Plant Physiol. Plant Mol. Biol. 48, 575-607. [ Links ]
Hebbar, P. 2007. Cacao diseases: a global perspective from an industry point of view. Phytopathol. 97, 1658-1663. [ Links ]
Islam, M. and G. Mayo. 1990. A compendium on host genes in flax conferring resistance to flax rust. Plant Breed. 104, 89-100. [ Links ]
Johnson, E., F. Bekele, S. Brown, Q. Song, D. Zhang, L. Meinhartd, and R. Schnell. 2009. Population structure and genetic diversity of the Trinitarian cacao (Theobroma cacao L.) from Trinidad and Tobago. Crop Sci. 49, 564-572. [ Links ]
Kuhn, D., M. Heath, R. Wisser, A. Meerow, J. Brown, U. Lopes, and R. Schnell. 2003. Resistance gene homologues in Theobroma cacao as useful genetic markers. Theor. Appl. Genet. 107, 191-202. [ Links ]
Merchán, V. and A. Restrepo. 1980. Calibration of a method of inoculation for Moniliophthora roreri. Annual Activity Report 1979 B - 1980A. Instituto Colombiano Agropecuario - (ICA), Bogota. [ Links ]
Motamayor, J., A. Risterucci, A. Lopez, C. Ortiz, A. Moreno, and C. Lanaud. 2002. Cacao domestication I: the origin of the cacao cultivated by the Mayas. Heredity 89, 380-386. [ Links ]
Phillips-Mora, W., J. Castillo, U. Krauss, E. Rodríguez, and M. Wilkinson. 2005. Evaluation of cacao (Theobroma cacao) clones against seven Colombian isolates of Moniliophthora roreri from four pathogen genetic groups. Plant Pathol. 54, 483-490. [ Links ]
Phillips-Mora, W. and M. Wilkinson. 2007. Frosty pod of cacao: a disease with limited geographic range but limited potential for damage. Phytopathol. 97, 1644-1647. [ Links ]
Ploetz, R. 2007. Cacao diseases: important threats to chocolate production worldwide. Phytopathol. 97, 1634-1639. [ Links ]
Ribeiro, F., J. Parlevleliet, and L. Zambolim. 2001. Concepts in plant disease resistance. Fitopatol. Bras. 26, 577-589. [ Links ]
Rodríguez, E. and J. Medina. 2005. Caracterización de clones de cacao por respuesta a Monilia, Moniliophthora roreri (Cif), en Santander. Fitopatol. Colomb. 28. 2. [ Links ]
Sánchez, J. and L. González. 1989. Metodología para evaluar la susceptibilidad a moniliasis en cultivares de cacao (Theobroma cacao). Turrialba 39(4), 461-468. [ Links ]

