










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.29 n.2 Bogotá maio/ago. 2011
PLANT BREEDING, GENETIC RESOURCES & MOLECULAR BIOLOGY
Identification of radiation induced mutants of cassava (Manihot esculenta Crantz) using morphological and physicochemical descriptors1
Identificación de mutantes radio-inducidos de yuca (Manihot esculenta Crantz) mediante descriptores morfológicos y fisicoquímicos
Adriana Tofiño, 6,Diana Cabal3, Teresa Sánchez4, and Hernán Ceballos5
1Article derived from Ph.D. thesis by first author..2Experimental Station Motilonia, Corporación Colombiana de Investigación Agropecuaria (Corpoica). Codazzi (Colombia).
3Agroindustrial Engineering, Universidad Popular del Cesar. Valledupar (Colombia).
4 Cassava Program, International Center for Tropical Agriculture (CIAT). Cali (Colombia).
5Faculty of Agricultural Sciences, Universidad Nacional de Colombia; Cassava Program, International Center for Tropical Agriculture (CIAT). Cali (Colombia).
6Corresponding author. atofino@corpoica.org.co Recceived for publication: 14 February, 2010. Accepted for publication: 2 June, 2011.
ABSTRACT
For mutants of starch and other desirable traits of interest in cassava, we evaluated the physical-chemical morphology of the root and paste in inbred populations of six M2 families, derived from irradiated seeds. 1,097 plants were established, 829 of Gamma treatment and 268 of neutrons. In the morphological description, most chimeras recorded in M1 were identified except hermaphrodite flowers and bark of different colors on the same plant, new abnormalities in M2 were found, such as root without suberification, no anthesis, early flowering, meristems and changes in the supply-demand relationship. However, mortality, reduced vigor, yield and anthesis loss have also been identified as effects of inbreeding. Simple techniques such as optical microscopy and iodine staining were effective for the identification of mutants of the granule and the results were confirmed with electron microscopy analysis and pulp properties, except in mutant granule hole, in which no abnormalities of the granule were correlated with significant variations in the paste. However, Pearson correlations associated the hot paste viscosity with consistency and reorganization. Variability was found in the granule characteristics and properties of the paste between years, suggesting significant influence of genotype-environment interaction. Similarly, there were differences in paste properties associated with the type of radiation. In addition, promising mutants were identified with post-harvest physiological deterioration tolerance associated mainly to gamma.
Key words: inbreeding, optical microscopy, viscosity of the starch, root quality, morphological, mutagenesis.
RESUMEN
En busca de mutantes deseables del almidón y otros caracteres de interés en yuca, se evaluó la morfología, físico química de raíz y la pasta, en poblaciones de seis familias endocriadas M2, derivadas de semillas irradiadas. Se establecieron 1.097 plantas, 829 del tratamiento Gamma y 268 de neutrones. En la descripción morfológica, se identificaron, la mayoría de las quimeras registradas en M1, excepto flores hermafroditas y corteza de diferente color en la misma planta; se encontraron nuevas anormalidades en M2, como raíces sin suberificación, ausencia de apertura floral, floración anticipada, alteraciones en meristemos y relación fuente demanda. Sin embargo, la mortalidad, reducción del vigor, rendimiento y pérdida de la apertura floral, también se han identificado como efectos de endocría. Técnicas sencillas como microscopía óptica y tinción por yodo fueron efectivas para la identificación de mutantes del gránulo y los resultados se ratificaron con análisis de microscopía electrónica y propiedades de la pasta, excepto en los mutantes de gránulo hueco, en los que no se correlacionaron anormalidades del gránulo con variaciones significativas en la pasta. Sin embargo, correlaciones de Pearson asociaron viscosidad de pasta caliente con reorganización y consistencia. Se encontró variabilidad en características del gránulo y propiedades de la pasta entre años, lo cual sugiere influencia significativa de la interacción genotipo ambiente. De igual forma, se presentaron diferencias en las propiedades de la pasta asociadas con el tipo de radiación. Además, se identificaron mutantes promisorios con tolerancia al deterioro fisiológico poscosecha asociados principalmente, a gamma.
Palabras clave: endogamia, microscopía óptica, viscosidad del almidón, calidad de raíces, caracterización morfológica, mutagénesis
Introduction
Cassava (Manihot esculenta Crantz), is a key component in the diet of one billion people, thanks to its high content of starch in the roots, with values up to 85% (FAO and IFAD, 2000). To consolidate the industrial expansion of cassava, one is required to find value-added features with high yields. Other challenges in crop improvement include: decreasing post-harvest physiological deterioration (PPD), tolerance to major biotic and abiotic constraints endemic in every area of cultivation, manipulation of numbers and floral synchrony between cultivars. To achieve these objectives, we have modified the conventional breeding methods and biotechnology tools are being implemented (Tofiño et al., 2007).
Mutagenesis is a strategy to create variability in agronomically important characters and quality of agricultural products (Liu et al., 2004). We have identified mutant populations, large numbers of morphological flowering and fruiting distortions (Masuda et al., 2004; McManus et al., 2006); morphological and physiological alterations in vegetative states (Wu et al., 2005; Xin et al., 2009); and increased tolerance to biotic and abiotic factors (Anderson et al., 1996; Kowalski and Cassels, 1999; Nagata et al., 1999; Rai, 2001; Dita et al., 2006).
Specifically, starchy species have been obtained by mutation- induced changes in the characteristics of starch such as complete deletions in the Waxy locus (Kawakami et al., 2007), decreased size of rice granule accumulation and modification in the amylopectin structure (Satoh et al., 2008), differences in the rate of starch digestibility (Groth et al., 2008) and series of point mutations in the Waxy gene in potatoes with high amylopectin content (Muth et al., 2008).
Additionally, the number of mutations that occur in a mutant population depends on the interaction of multiple factors and the likelihood that the mutation produces an altered phenotype (Wu et al., 2005; Nag and Smerdon, 2009). CIAT, the research group of the cassava breeding improvement program, reported the discovery of the mutation of small granule in the M2 population (Ceballos et al., 2008). However, other atypical characteristics in the plants or quality of roots have not been reported in these populations. According to the above, this article proposes to confirm M2 inbred populations, the occurrence of atypical morphological features of interest, previously identified in generating chimeric M1 (Tofiño et al., 2008) and advance the understanding of the effect of ionizing radiation on cassava germplasm, especially on some morphological characteristics and quality of the plant roots.
Materials and methods
Irradiation was done with ? rays 200 Gy (Cobalt 60) and fast neutrons 0.2 MeV (235 U), on 1,400 seeds of six families with full and half brothers (CM9331, GM155, SM3015, SM3045, C4 and C127). We performed self-pollination on M1 plants, to obtain seeds and M2 plants.
Evaluation -M2 plants
Phenotypic evaluations were performed according to the descriptors of stem, leaves and roots of Fukuda and Guevara (1998), to screen potential mutants, identified in M2, M1 registered chimeras (Micke and Donini, 1993). Within populations, survival was evaluated at 180, 240 and 300 days after seeding. Vigor and flowering were also evaluated. Physico-chemical, microscopic and viscosity analyses were carried out to identify individuals with unusual characteristics in the roots, with respect to limit values recorded in the cassava germplasm bank, including non-irradiated parents (Ceballos et al., 2007). Analyses were conducted in two crops (sexual and clonal seed), amylase, dry matter, cyanide and PPD. The PPD was evaluated by measuring the level of damage to perimeter roots at 8 and 15 days after harvest (dah).
Determination of the morphology of starch granules by light and electron microscopy. Trans-sectional root cuttings were rubbed on slides with a drop of 0.2% iodine subsequently added, and observed by light microscopy (Olympus CX41) at 40X. Individuals with abnormalities in the granule were evaluated by electron microscopy. Starch granules were dusted on double-sided tape mounted on circular aluminum, covered with 35 nm of goldaluminum, photographed and measured in a Scanning Electron Microscope (Jeol JSM 820, Tokyo, Japan) with an accelerating voltage of 20 kV.
Amylase content by iodine staining
The harvested roots were cut crosswise and sprayed with 2% iodine (2 g KI and 0.2 g I2 in 100 cm3 of distilled water). Determination of dry matter content of cassava roots. Five g of grated cassava was placed in a crucible of known weight and placed in an oven at 60®C. The crucible was removed and placed in a drying hood to reach room temperature. It was weighed and the difference settled with the fresh weight and taken as a percentage.
Determination of cyanide content of cassava roots
Forty g cassava had 50 mL of extraction buffer added and liquefied. The liquefied sample was centrifuged at 6,000 rpm. 0.4 mL of 0.1 M phosphate buffer pH 7.0 was added and a 0.1 mL sample was taken and placed in a water bath at 30®C. 0.1 mL of linamarase was taken, centrifuged, bathed at 30®C. 0.6 mL of 0.2 N NaOH was added as well as 2.8 mL of 0.1 M buffer pH 6.0 and 0.1 mL of chloramine T 1%. 0.6 mL of color reagent was added as well as 0.1 mL of chloramine T, and 0.6 mL of color reagent, and read by spectrophotometry at 605 nm (Aristizabal et al., 2007).
Determination of amylase
For 0.1 g of starch was gathered in 100 mL balls and 1 mL of absolute ethanol at 95% was added and homogenized. Nine mL of 1N sodium hydroxide was added. A volume of 100 mL was reached with distilled water. One mL of acetic acid and 2 mL of 2% iodine were added. A volume of 100 mL was reached with distilled water, reacted in the dark and read at 620 nm (Aristizabal et al., 2007).
Analysis of the viscosity of the paste by RVA The profiles of hot paste viscosity were obtained by RVA4 Series model RVA (Newport Scientific, Australia). 5% suspension (w/v) dry basis of each sample was added to aluminum viscoamilografic containers and subjected to the temperature program (Ceballos et al., 2007).
Data analysis
Statistical analysis of the variables evaluated in the field and laboratory was performed using the SAS® statistical package version 9.1 and frequency tables were made with the qualitative variables. Analysis of variance was performed with untransformed data, using the Tukey test to determine differences between families, treatments and years. Similarly, we applied the Pearson correlation test between the variables measured. The coefficient of variation and the coefficient of model determination were 12% and 0.9, respectively.
Results and discussion
The 1,097 M2 plants were established, of which 829 received Gamma treatment and 268 neutron treatment. Ionizing radiation has largely contributed in the production of varieties in crops of economic importance (Tofiño et al., 2007; Xin et al., 2009).
Morphological characterization of mutants, individual seedling is commonly performed on cereals in later generations, M3 and M4, which has been identified as alterations in the color of the organs, leaf characteristics and reproductive abnormalities (Wu et al., 2005; Xin et al., 2009). In the case of rice and Lotus japonicus, the assessment of an average of 40,000 M2 plants was required to detect mutant genotypes in genes involved in a wide range of metabolic and developmental processes (Wu et al., 2005).
The verification of the identified abnormalities in M1, was performed in M2 without observing hermaphrodite flowers or stem bark of different color in the same plant. In contrast, apomictic plants are identified with weak form, state of descriptor form of leaflets and atypical leaf area, not recorded in cassava. Similarly, new abnormalities were identified such as root without suberification, no anthesis, early flowering, changes in demand and supply ratio, and meristems distortion. In the populations tested, at least one atypical feature was identified in 62.2% of individuals (Fig.1, Tab.1). Overall, the heterogeneity of radiation sensitivity was evident in the population, the SM3045 family with gamma treatment showed 13.5% of individuals with at least one abnormality while the same family under neutron treatment presented 100% atypical individuals. In contrast, the C4 family had more than 80% atypical individuals under both ionizing radiations (Tab.1). Also, 17% of plants were weak, smaller than 30 cm, (Tab.1). However, this feature, or the decrease in root production, is associated with inbreeding depression. Therefore, these criteria should not be interpreted as indicators of the effect of the mutation in M2 (Rojas et al., 2009). Evolutionary studies of cassava and population genetic models indicate that the size of the plants is correlated with multilocus heterozygosity (Pujol and McKey, 2006) and possible causes of inbreeding depression on the characteristics of agronomic importance in open or mixed pollinated plants, associated mainly to the expression of deleterious recessive alleles (Charlesworth and Charlesworth, 1999).
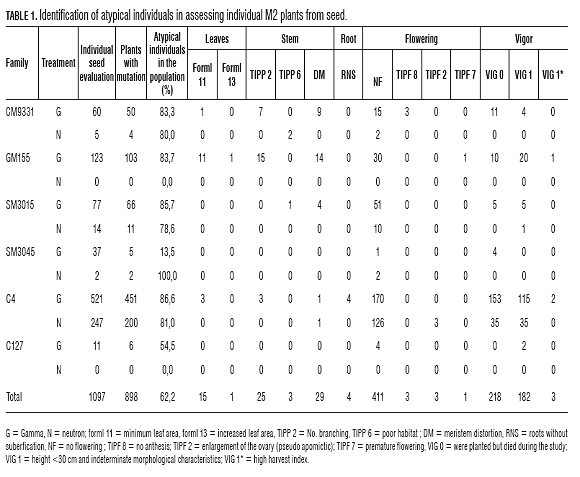

High rates of mortality were identified in M2 populations, 20%, as well as overall decline in the production of roots, less than 5 kg per plant, (Tab.1). The above features cannot be associated exclusively with the effects of radiation because inbreeding depression effects mortality in the field and in surviving plants. Studies on cassava suggest that the rate is not uniform among inbred families (Rojas et al., 2009), which may become apparent between the mutant inbred populations, especially in the SM3015 and C4 families which showed mortality and presence of atypical individuals as highly variable between sources of radiation.
On the other hand, despite the inclusion of at least one inbred generation in new models for improving cassava, the effect of inbreeding on granule size and other characteristics of the starch has not yet been registered. The decrease in root size and performance observed among related genotypes, non-irradiated grandfather, M1 parent and the M2 mutant with small granule is accompanied by changes in size, shape and density of starch granules, detectable by microscopy (Fig.2).
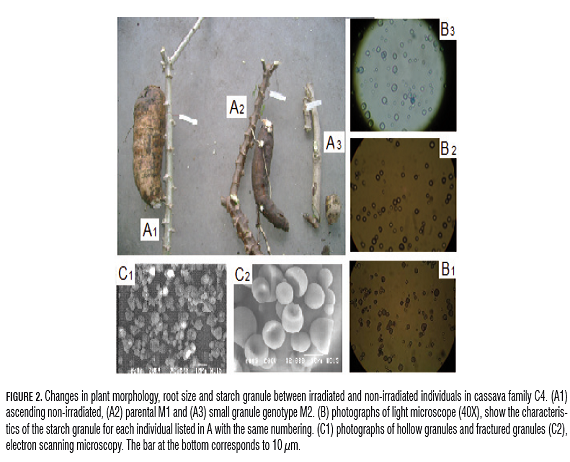
However, the combination of microscopic techniques, root staining with iodine (Carvalho et al., 2004) and morphological description (Singh et al., 2006), have been used successfully in the characterization of variability and identification of mutant cassava starch and different crops. Iodine staining has been useful in identifying natural mutants in cassava such as Waxy and Sugary; in both studies the preliminary observation of the results of this simple, low cost technique was confirmed with biochemical and molecular analysis (Ceballos et al., 2007).
However, staining with iodine only allows discrimination because extreme genotypes with thick, reddish-brown coloration is typical of amylase-free starches, while low starch or high sugar content does not cause visible changes in the initial coloring of the solution. The starch content of 16.6 to 23.6% of amylase stains dark blue root (Ceballos et al., 2007). A scope with a finer discrimination is required for quantification of amylase by spectrophotometer or Differential scanning calorimeter (DSC) (Ceballos et al., 2008). After iodine staining, an individual was identified without appreciable changes in the initial coloration of the solution that could be associated with real sugar (GM155N35-1). Similarly, optical microscopy allowed the identification of small granules of M2 genotypes, with a predominance of 5-8 µm diameter (C4G160-13, C4G160-16, C4G160- 18, SM3015-1), granule holes (C4N83-9, and C4G28 GM155G28-9-10) and fractured granules (GM155G75-1) (Fig.2). The combination of both techniques allowed the identification of individuals with little starch (C4G190- 11, C4N14-5, C4N14-14, CM9331, G67-36), no change in the color of the solution and a low number of granules. In rice, induced mutants were identified with features that correspond to defects in the gene Pho1 such as variability in wrinkling, mature seed size and starch content (Wu et al., 2005).
One of the main objectives of the establishment of a clonal test with sexual M2 seed, was to verify the persistence of the detected abnormalities and thus rule out a possible masking effect in the interaction between the environment genotype and the M2 mutant population. Especially in the expression of granule abnormalities, rheological properties of starch, amylase (Am), dry matter (DM) and cyanide (HCN). With respect to behavior in the two assessments, analysis of variance and Tukey's test suggest that C4 differed statistically from the other families in the accumulation of MS and HCN; gamma treatment showed a general trend toward the accumulation of DM and HCN, and the neutron treatment showed a significant effect between years in the production of Am, MS, and HCN. The C4 family presented greater variability in the distribution of data for the gamma treatment and the lowest and highest value of MS and Am with 14.8, 71.6 and 17.2, 30.67, respectively.
Similarly, in the C4 neutron treatment, the highest value of HCN was found while the lowest value was recorded in GM155, 1,643 and 45 ppm, respectively. In studies of starches from different sources: cereals, legumes, roots and tubers, the correlation between thermal, rheological, Am and granule size has been proven (Li and Yeh, 2001). In this vein, it is suggested that the characterization of starch mutants must be made from the integrated analysis of these characteristics over time and for greater reliability, in various locations.
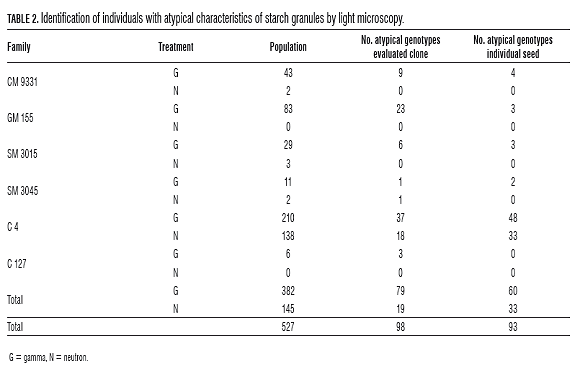
The evaluation of the morphology of starch granules by light microscopy showed the effect of genotype-environment interaction because there were differences in the number of individuals with abnormalities between years, in every family of gamma treatment. In the treatment with neutrons, they found abnormalities in the morphology of the granule in only the families SM3045 and C4 in both evaluations (Tab.2).
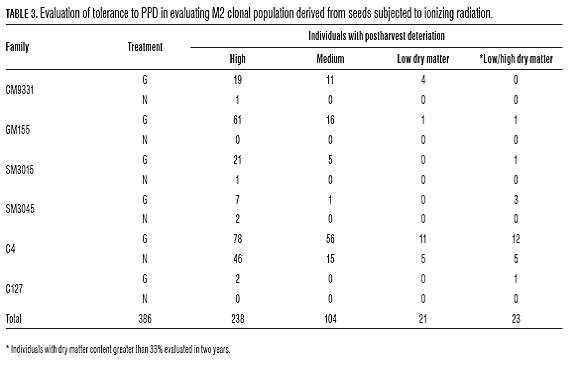
For the evaluation of physiological deterioration (PPD), we performed two readings from 8 to 15 dah. The analysis of variance and Tukey's test indicate that family SM3045 was more prone to deterioration and the treatment presented significant effect on the intensity of damage, neutron showed higher vascular obstruction while gamma higher rotting. Materials with low damage after 15 d, were associated with dry matter content as there is an undesirable inverse relationship between PPD and MS in clones with moderate content of carotenoids (El-Sharkawy and Cadavid, 2002; Sánchez et al., 2006). We identified 23 individuals in families C4, SM3045, SM3015 and GM155 of gamma, in descending order according to the number of individuals identified. In the neutron treatment promising individuals were identified only in C4 and SM3045 (Tab.3). It is recommended that individuals with high promise in PPD and dry matter be evaluated in areas with contrasting rainfall since differences in the expression of PPD have been recorded due to sowing environment (El-Sharkawy and Cadavid, 2002).
Several studies of wild starches (Blazek and Copeland, 2008; Kong et al., 2009) and mutants have correlated differences in the characteristics of the starch granule, chemical and physical properties, and content of amylase and amylopectin, with changes in the pattern viscosity. This is true in the case of rice, a mutant resistant starch exhibited reduced viscosity of the paste, low peak temperature and final temperature, and enthalpy of gelatinization (Shu et al., 2006). However, in hollow granule mutants C4N83-9, GM155G28-9-10 and C4G28, there is no correlation between granule abnormalities and significant variations in the properties of the paste. Contrary to what was observed in the small granule mutant, in which granule analysis, quality of roots and critical values of RVA show changes between the generations: non-irradiated grandfather, M1 parent and M2 mutant with respect to gradual decline in performance, granule size, instability and clarity of the gel, and dry matter content of Am (Tab.4, Fig.2 and Fig.5).
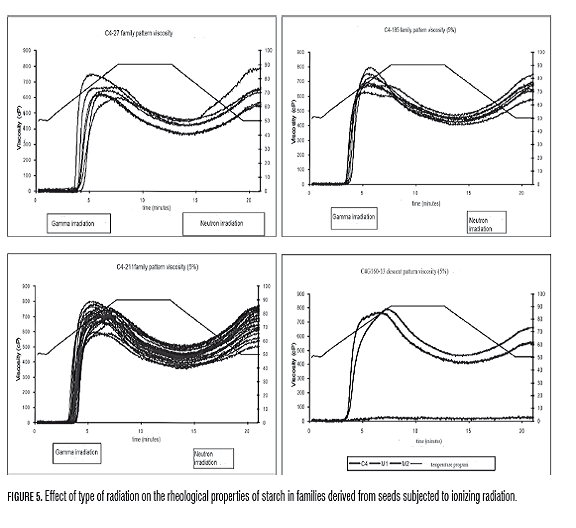
The RVA analysis has been useful in the identification and characterization of yucca mutants such as Waxy and small granule, in which the values of peak viscosity, breakdown and reorganization of the gel are drastically different from normal starch (Ceballos et al., 2007, Ceballos et al., 2008). Quality variables in roots and RVA, analysis of variance and Tukey test indicate significant differences between GM155 and the other families in hot paste viscosity and consistency. The hot paste viscosity is also affected differentially by the treatment, and the neutron treatment which showed the highest values for paste temperature, while gamma showed the highest values in gel consistency. On the other hand, the viscosity was very sensitive to the effect of seed type as assessed for two years, there were statistical differences in the variables: maximum viscosity (Vm), hot paste viscosity (VPC), cold paste viscosity (Vpf), instability of the gel (Ig), reorganization (Rg) and consistency (Cg). Differences in the evaluation for two years indicate influence of genotype-environment interaction in the expression of these characters, cassava M2. In this sense, there was more variability in the rates of RVA in the clonal evaluation and a strong tendency to produce more viscous starches (Fig.4). Concerning the effect of radiation on the pulp properties in C4 family, the neutron treatment produced more viscous starches than gamma due to higher values in Vm and Ig while gamma produced starches more prone to retrogradation due to elevated Rg and Cg (Fig.5).
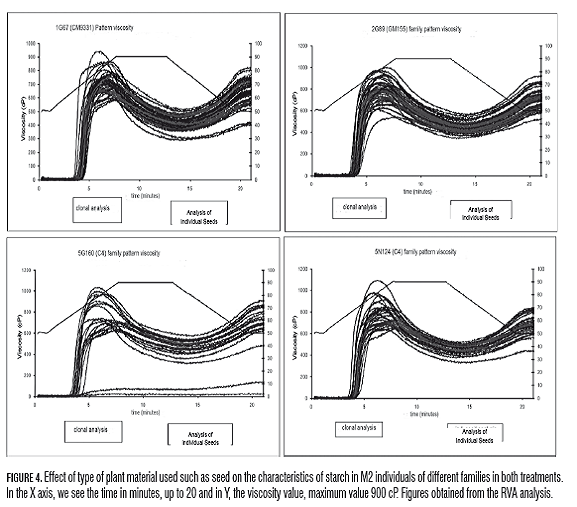
In the potato and Amaranthus, differences have also been identified between varieties in Am, swelling power and solubility, as well as peak temperature, and a tendency for retrogradation Ig (Singh et al., 2006; Kong et al., 2009). Location research in the potato reveals the influence of the genotype-environment interaction on granule size and paste temperature, and temperature-dependent tuber development (Kaur et al., 2007).
The relationships between different properties of starch determined by the Pearson correlation analysis showed positive correlation between Vpc, Rg (r2 = 0.673, P=0.001) and Cg (r2 = 0.311, P=0.001) but negatively correlation with Ig (r2 = -0.446, P=0.001). The Vm showed positive correlation with Vpf (r2 = 0.0.809, P=0.001), Ig (r2 = 0.738, P=0.001) and Cg (r2 = 0.755, P=0.001), while Rg showed negative correlation (r2 = -0.380, P=0.001). These correlations differ from those reported in potato germplasm and localities in which Vpc was positively correlated with Vpf and Te (pasting temperature), while Rg was negatively correlated with Te (Kaur et al., 2007).
Am´s correlation with some variables such as viscosity has been reported in cassava and other starch (Li and Yeh, 2001, Kong et al., 2009). In the integrated analysis of the two M2 assessments, Am and Vpc were negatively correlated (r2 = -0.44, P=0.001), Vm (r2 =-0.25, P=0.010); Vpf (r2 = -0.285, P=0,0038) and R (r2 = -0.30, P=0.002). These correlations are inverse to those recorded in the potato where Am was positively correlated with VPC (Kaur et al., 2007) but are consistent with the behavior of cassava starch type Waxy, enriched with amylopectin, peak viscosity is higher while in the small granule starch with unusually high content of amylase for cassava, the peak viscosity is low (Ceballos et al., 2007, 2008). In addition to Amylase content, variation in the content and characteristics of amylopectin affects the properties of the paste, especially Te and in some cases Vm (Shu et al., 2006).
Conclusions
In M2, 80% of the M1 chimeras and new abnormalities in M2 were identified from radiation similar in mutagenic efficiency (82%). However, fast neutrons, further limits the survival and reproductive capacity of M1 plants, which reduced the number of M2 plants derived from such treatment. However, mortality, reduced vigor, yield and anthesis loss are associated with effects of inbreeding in cassava. Variability in the characteristics of the granule and pulp properties was found in assessments of two years, suggesting significant influence of genotype-environment interaction in these characteristics.
The phenotypic evaluation of inbred M2 populations of mutant cassava germplasm allows the identification of atypical individuals but does not clarify the genetic origin since changes in the starch may be associated with gene expression of an intricate network of genes, where the posttranscriptional relationship is equally significant. Similarly, in M2 populations, effects of inbreeding depression come together, increasing homozygosity of the genetic background and possible fixation of mutations induced by ionizing radiation in sexual seeds. So, it is necessary to apply molecular techniques such as TILLING and Insitu expression to identify the origin of the phenotypic abnormalities identified in the M2 population.
Acknowledgements
The authors are grateful to Colciencias.
Literature cited
Anderson, P., P. Okubara, R. Arroyo-García, B. Mayers, and R. Michelmore. 1996. Molecular analysis of irradiation-induced and spontaneous deletion mutants at a disease resistant locus in Lactuca sativa. Mol. Gen. Genet. 251(3), 316-325. [ Links ]
Aristizabal, J., T. Sanchez, and D. Mejía. 2007. Technical guidelines for production and analysis of cassava starch. Bulletin FAO Agricultural Services No. 163. Rome. [ Links ]
Blazek, J. and L. Copeland. 2008. Pasting and swelling properties of wheat flour and starch in relation to amylase content. Carb. Polym. 71(3), 380-387. [ Links ]
Charlesworth, B. and D. Charlesworth. 1999. The genetic basis of inbreeding depression. Genet. Res. 74:329-340. [ Links ]
Carvalho L.J.C.B., C.R.B. De Sousa, J.C.M. De Cascardo, C.B. Junior, and L. Campos. 2004. Identification and characterization of a novel cassava Manihot esculenta Crantz clone with high free sugar content and novel starch. Plant Mol. Biol. 56, 643-659. [ Links ]
Ceballos, H., T. Sanchez, N. Morante, M. Fregene, D. Dufour, A. Smith, K. Denyer, J. Perez, F. Street, and C. Mestres. 2007. Discovery of an amylase-free starch mutant in cassava (Manihot esculenta Crantz). J. Agric. Food Chem. 55, 7469-7476. [ Links ]
Ceballos, H., T. Sanchez, K. Denyer, A. Tofino, E. Rosero, D. Dufour, A. Smith, N. Morante, J. Perez, and B. Fahy. 2008. Induction and identification of a small-granule, high-amylase mutant in cassava (Manihot esculenta Crantz). J. Agric. Food Chem. 56(16), 7215-7222. [ Links ]
Dita, M., N. Rispail, E. Prats, D. Rubiales, and K. Singh. 2006. Biotechnology approaches to biotic and abiotic stress overcome constraints in legumes. Euphytica 147(1), 1-24. [ Links ]
El-Sharkawy, M. and L. Cadavid. 2002. Response of cassava to prolonged water stress imposed at different stages of growth. Exp. Agric. 38, 333-350. [ Links ]
FAO, Agriculture Organization of the United Nations and IFAD, International Fund For Agricultural Development Food. 2000. The world cassava economy: facts, trends and prospects. Rome. [ Links ]
Fukuda W. and C. Guevara. 1998. Morphological and agronomic descriptors to characterize cassava (Manihot esculenta Crantz). Doc. 78. Embrapa; Cnpmf, Cruz das Almas BA, Brazil. [ Links ]
Groth, D., J. Santini, B. Hamaker, and Weil, C. 2008. High-throughput screening of ems mutagenized maize for altered starch digestibility. Bioenergy Res. 1(2), 118-135. [ Links ]
Kaur, A., N. Singh, R. Ezekiel, and H. G. Singh. 2007. Physicochemical, thermal and pasting properties of potato starches separated from different cultivars grown at different locations. Food Chem. 101(2), 643-651. [ Links ]
Kawakami, S., K. Kadokawi, R. Morita, and M. Nishimura. 2007. Induction of a large deletion including the waxy locus by thermal neutron irradiation in rice. Breed. Sci. 57(2), 153-157. [ Links ]
Kong, X., J. Bao, and H. Corke. 2009. Physical properties of Amaranthus starch. Food Chem. 113 (2), 371-376. [ Links ]
Kowalski, B. and A. Cassells. 1999. Mutation breeding for yield and Phytophthora infestans (Mont) of Bary foliar resistance in potato (Solanum tuberosum L. cv. Golden Wonder) using computerized image analysis in selection. Potato Res. 42(1), 121-130. [ Links ]
Li, J. and A. Yeh. 2001. Relationships between thermal, rheological characteristics and swelling power for various starches. J. Eng. Food 50(3), 141-148. [ Links ]
Liu, L., L. Van Zanten, Q. Shu, and M. Maluszynski. 2004. Officially released mutant varieties in China. Mutat. Breed. Rev. 14, 1-62. [ Links ]
McManus, L., J. Sasse, C. Blomstedt, and G. Bossinger. 2006. Treatment for mutation induction Pollen in Eucalyptus globules ssp. globules (Myrtaceae). Aust. J. Bot. 54(1), 65-71. [ Links ]
Micke, A. and B. Donini. 1993. Induced mutations pp. 1-21. In Hayward, M.D., N.O. Rosemark, and I. Romagosa (eds.). Plant breeding, principles and prospects. Chapman & Hall, New York, YN. [ Links ]
Muth, J., S. Hartje, R. Twyman, H. Hofferbert, E. Tacke, and D. Prüfer. 2008. Precision breeding for novel starch variants in potato. Plant Biotechnol. J. 6(6), 576-584. [ Links ]
Nag, R. and M. Smerdon. 2009. Altering the chromatin landscape for nucleotide excision repair. Mut. Res. 682(1), 13-20. [ Links ]
Nagata, T., S. Todoriki, T. Hayashi, Y. Shibata, M. Mori, H. Kanegae, and S. Kikuchi. 1999. a-radiation induces leaf trichome formation in Arabidopsis. Plant Physiol. 120, 113-119. [ Links ]
Masuda, M., S. Agong, A. Tanaka, N. Shikazono, and Y. Hase. 2004. Mutation spectrum of tomato seed induced by radiation with helium ion beams and coal. Acta Hort. 637, 257-262. [ Links ]
Pujol, B. and D. McKey. 2006. Asymmetry in intraspecific competition size and the density-dependence of inbreeding depression in a natural plant population: a case study in cassava esculenta Crantz, Euphorbiaceae). J. Evol. Biol. 19(1), 85-96. [ Links ]
Rai, S. 2001. Pleiotropic morphological and abiotic stress resistance phenotypes of the hyper- abscisic acid producing Abo-mutant in the periwinkle Catharanthus roseus. J. Biosci. 26(1), 57-70. [ Links ]
Rojas, M., J. Perez, H. Ceballos, D. Baena, N. Morante, and F. Calle. 2009. Analysis of inbreeding depression in eight S1 cassava families. Crop Sci. 49, 543-548. [ Links ]
Sánchez, T., A. Chavez, H. Ceballos, D. Rodriguez-Amaya, P. Nestel, and M. Ishitani. 2006. Reduction or delay of post-harvest physiological deterioration in cassava roots with higher carotenoid content. J. Food Agric. 86(4), 634-639. [ Links ]
Satoh, H., K. Shibahara, T. Tokunaga, A. Nishi, M. Tasaki, S. Hwang, T. Okita, N. Kaneko, N. Fujita, M. Yoshida, Y. Hosaka, A. Sato, Y. Utsumi, T. Ohdan, and Y. Nakamura. 2008. Mutation of the plastidial-glucan phosphorylase gene in rice affects the synthesis and structure of starch in the endosperm. Plant Cell 20, 1833-1849. [ Links ]
Singh, J., O. McCarthy, and H. Singh. 2006. Physico-chemical and morphological characteristics of New Zealand Taewa (Maori potato) starches. Carb. Polym. 64(4), 569-581. [ Links ]
Shu, X., S. Shen, J. Bao, D. Wu, Y. Nakamura and Q. Shu. 2006. Molecular and biochemical analysis of the gelatinization temperature of rice characteristics (Oryza sativa L.) starch granules. J. Cereal Sci. 44(1), 40-48. [ Links ]
Tofiño, A., M. Romero, M. Fregene, and A. Rosero. 2007. Possibilities and scope of genetic improvement based on induced mutation in cassava (Manihot esculenta Crantz). Fitot. Colomb. 7(1), 1-12. [ Links ]
Tofiño, B.C., D. Cabal, H. Romero, and H. Ceballos. 2008. Morphological characterization of M1 population of cassava (Manihot esculenta Crantz) derived from irradiated seeds. Fitot. Colomb. 8(1), 39-55. [ Links ]
Wu, J., C. Wu, C. Lei, M. Baraoidan, A. Bordeaux, M. Madamba, M. Ramos-Pamplona, R. Mauleon, A. Portugal, V. Ulate, R. Bruskiewich, G. Wang, J. Leach, G. Khush, and H. Leung. 2005. Chemical-and irradiation-induced mutants of rice IR64 for forward states and reverse genetics. Plant Mol. Biol. 59, 85-97. [ Links ]
Xin, Z., M. Wang, G. Burow, and J. Burke. 2009. An induced population sorghum mutant suitable for bioenergy research. Bioen.Res. 2(1-2), 10-16. [ Links ]

