










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.29 n.3 Bogotá set./dez. 2011
PROPAGATION AND TISSUE CULTURE
Morphological and anatomical analyses of the seed coats
Análisis morfológico y anatómico de las cubiertas de semillas de granadilla (Passiflora ligularis Juss.)
Julián Cárdenas-Hernández1,2, Diego Miranda L.1, Stanislav Magnitskiyand1 and Carlos Carranza1
1Department of Agronomy, Faculty of Agronomy, Universidad Nacional de Colombia. Bogota (Colombia).2Corresponding author. julianentero@gmail.com Received for publication: 15 June, 2011. Accepted for publication: 2 November, 2011.
ABSTRACT
The study of histology and morphology of seeds of genus Passiflora has been of high utility for the classification of species. In seeds of sweet granadilla, the histological characteristics and methodologies for their study are unknown. This study was aimed to know the tissue and morphological characteristics of the seed coats of seeds of sweet granadilla and to be able to determine its value in the differentiation of accessions. Five accessions collected in producing zones of the Province of Huila, Colombia, were analyzed. In morphological analysis, all accessions presented falsifoveate ornamentation and entire margin. The seeds presented high change in weight and size for every accession; there stood out the seeds of the accession PmN for presenting major size (7.42 mm long), weight (35.62 mg), homogeneity in these variables and a typical orange color. For the histological analysis, a protocol was adapted to realize sections of seed coats in parafin, by means of which one managed to obtain sections (7 µ) that in the optical microscope show clearly three well differentiated layers, belonging, possibly, to exotegmen (internal layer), mesotesta (medium layer) and exotesta (external layer). Every layer presented differences in the form of the cells, color and thickness, between the sections of the basal and medium parts of the seeds, but the differences between the analyzed accessions were not observed. The majority of seeds had a thickness of seed coats that varied among 235 and 475 µ.
Key words: exotegmen, mesotesta, exotesta, macrosclereids, taniferous cells.
RESUMEN
El estudio de la histología y morfología de las semillas del género Passiflora ha sido de gran utilidad para la clasificación de especies. Sin embargo, las semillas de granadilla no han sido investigadas. Con este estudio se buscó conocer las características tisulares y morfológicas de la cubierta de las semillas de granadilla y así poder determinar su valor en la diferenciación de accesiones. Se analizaron cinco accesiones colectadas en zonas productoras del departamento del Huila (Colombia). En el análisis morfológico, todas las accesiones presentaron ornamentación falsifoveada y borde entero. Las semillas presentaron gran variación en peso y tamaño dentro de cada accesión; se destacaron las semillas de la accesión PmN por presentar mayor tamaño (7,42 mm de longitud), peso (35,62 mg), homogeneidad en estas variables y un color naranja característico. Para el análisis histológico se adaptó un protocolo para realizar cortes de la cubierta de las semillas en parafina, mediante el cual se logró obtener cortes (7 µ) que al microscopio óptico muestran claramente tres capas bien diferenciadas, pertenecientes posiblemente a la exotesta (externa), mesotesta (media) y exotegmen (interna). Cada capa presentó diferencias en la forma de sus células, color y espesor, entre los cortes basales y los cortes medios de las semillas, sin diferencias claras entre las accesiones analizadas. La mayoría de las semillas mostraron un grosor de la cubierta entre 235 µ y 475 µ.
Palabras clave: exotegmen, mesotesta, exotesta, macroesclereids, células taníferas
Introduction
Morphology and anatomy seed research contributes to knowledge of taxonomy, evolution, and ecology of Angiospermae species (Cortez and Carmello-Guerreir, 2008). Passiflora is the most important genus of the family Passifloraceae and is represented by approximately 500 species. This taxon presents a wide natural distribution, with plants grown in the wild in the Americas, Western India, Galapagos Islands, Australia, Southeast Asia, Malaysia, Polynesia, and some islands of the Pacific Ocean (Vanderplank, 1990). Fruits of Passiflora sp. possess multiply seeds adhered to the funiculus on the ovary wall and surrounded by an aril that covers the seed and constitutes the eatable part of the fruit (Werker, 1997). The seeds of the family Passifloraceae present seed coats with generally lignified cells that not only affect water absorption but also offer a resistance to the embryo growth (Cardozo, 1988).
Seed coats develop from the integuments that surround the ovule prior to fertilization. Before fertilization, cells of the integuments are relatively undifferentiated. However, development after fertilization can include extensive differentiation of the cell layers into specialized cell types. In addition, some cell layers in the seed coats may accumulate large quantities of certain substances, such as mucilage or pigments that can also contribute to overall seed morphology (Moï sa et al., 2005).
The layers of the seed coats in seeds of Passiflora sp. are developed from bitegmical ovary, in which a radial unequal elongation of cells could be observed (Werker, 1997). The aril begins of a meristematic margin continued around the distal part of the raphe and includes the exostomal region. The pigments of the aril are located in the chromoplasts (Werker, 1997).
MacDougal (1994) realized a taxonomical review of the section Pseudodysosmia of subgenus Decaloba of genus Passiflora, and included the ornamentation of seeds among the characters that he used in its descriptions. This allowed considering the species of genus Passiflora to be an excellent object of study to characterize the seed morphology and, therefore, illustrate the taxonomical potential that the above mentioned structure presents for this genus (Pérez- Cortéz et al., 2005).
On this matter, Pérez-Cortéz et al. (2005) realized an anatomical description of seed coats of eight species of Passiflora sp., found eight anatomically different patterns and observed a layer of macrosclereid cells and a layer of obliterate cells in all the studied species. Nevertheless, P. ligularis was not among the analyzed species. The seeds of P. edulis, in the external tissues, present cells of palisade parenchyma, prismatic and lignified with thick reticulate walls, which form conical projections inside the endosperm. The endotesta has thick cell walls, prismatic elongated cells in the depressions of the tegmen and cuboids in the channels (Werker, 1997).
The seeds of P. ligularis are black, compressed, relatively small, and hard. The seeds and the aril contain about 17.6% carbohydrates (Trujillo, 1983; Werker, 1997). Pérez-Cortéz et al. (2002) describe the seeds of P. ligularis with transverse average section of elliptical form, basal of sharp form or sharply truncated, having ornamentation foveated or falsifoveated and entire margin.
The objective of the present research was to characterize the anatomy of seed coats of seeds of sweet granadilla using five accessions collected in producing zones corresponding to the South of Province of Huila, Colombia.
Materials and methods
The collection of five accessions was realized in the producing zone of sweet granadilla (Passiflora ligularis Juss.) in the South of Huila in the municipalities La Argentina, San Agustín and Palestina, the last one being the largest producer of sweet granadilla at national level.
The fruits were taken to the Laboratory of Crop Physiology of Faculty of Agronomy, Universidad Nacional de Colombia (Bogota), for their physico-chemical characterization. For the analysis of seeds, five accessions found in two commercial cultures were collected, one in the district La Mensura (2,012 m a.s.l.) and the other one in the district El Roble (1,807 m a.s.l.) of the municipality Palestina. The chosen accessions were named PrJ1, PrJ2, PrJJ, PmFD, and PmN; the first three were from the district El Roble and two last ones were from the district La Mensura.
Fifty seeds of every accession were taken at random; in each of these, the mass, length, width, thickness, and number of fosets were measured, the uniformity of distribution of fosets was described, the relation lenght-width (L/W) was calculated, and the presence of appendices in the base and the apex of the seeds was measured. In order to do this, scales of precision and a digital gauge were employed. The presence of appendices was described in the base and the apex of the seeds, enumerating them according to their presence. For this, some descriptors proposed by Pérez- Cortéz et al. (1995, 2002) were applied.
Finally, a photographic record of the seeds of every accession was realized with help of a microscope Nikon SMZ800 with integrated camera DSZMV using the software elements, in the Laboratory of Entomology of the Faculty of Agronomy, Universidad Nacional de Colombia, Bogota. In order to undertake histological sectioning, firstly, the protocol for sectioning in microtome in the Laboratory of Microbiology of Faculty of Agronomy of the Universidad Nacional de Colombia (Bogota) was adapted, taking into account the methodology used by Pérez-Cortéz et al. (2005, 2009). The principal methodological adaptations occurred in the processes of tissue softening with hydrochloric acid (18.5%) and fixation of tissues with Mayer Albumen.
Using the adapted protocol, sections of 7 µ of seed coats were done of the seeds of five chosen accessions. With help of the program Scopephoto 3.0® and optical microscope Micro Bm2000 with built-in camera DCMC510, the photographic record of sections was realized. The entire thickness of seed coat was measured as well as the thickness of each of three layers identified with help of the program Imagepro Express 6.3® de Medicybernetic Inc. Both morphological and histological analyses were descriptive ones.
Results
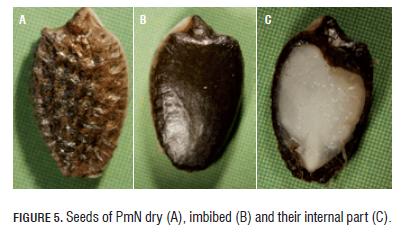
Morphological description of seeds of sweet granadilla Among the accessions no valuable differences in the type of ornamentation were recorded, since all accessions had falsifoveated type that is to say with asymmetrical fosets or foveas that were deeper and better delimited towards the central part of the seeds. All seeds presented entire margin and apical central horn (Fig.5)
When counted the number of appendices present (left and right) in the apical and basal zones of the seeds, it was found that these could possess from none up to two appendices for each zone; nevertheless, the presence or exact number of appendices was not considered an exclusive feature of every accession.
The dry seeds (9-11% moisture) presented the following colors: pale yellow, black, orange, and the most common coffee color; it was more common to find seeds with combination of these colors than monochrome seeds (Fig.1,2,3,4,5A). In all accessions, imbibed seeds (30-34% moisture) presented black color (Fig.1,2,3,4,5B). After the seed swallowed with the imbibition, the fosets were lost to produce a smooth surface. The membranous layer (exotesta) that surrounded the seeds could have been lost partially or completely (Fig.5,6). Internally, the seeds did not present apparent differences among the accessions. In seeds of all accessions, the embryo was straight and located in the central part of the base; two cotyledons that occupied the highest proportion of the internal space and a layer of endosperm surrounding these organs were observed (Fig.1,2,3,4,5C).
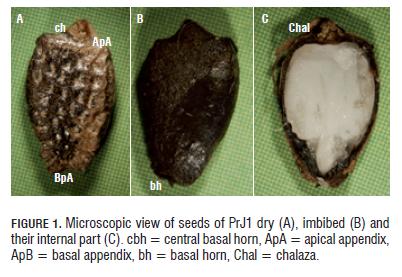



In dry state, the seeds of PrJ1 presented, in all cases, combined coloration: clear and black coffee (Fig.1A), while in imbibed state they were of black color (Fig.1B). In these seeds, the presence of apical and basal appendices was common on both sides (right and left) of the surface (Fig.1A). The presence of fosets was much desuniform in these seeds (Fig.1A). In the Fig. 1C, it is possible to appreciate the chalaza positioning towards the apical part of the seeds, which for this case was of coffee color. In some seeds, the presence of basal horn was observed (Fig.1B); this one was much smaller than the central basal horn (Fig.1A).
Some seeds of PrJ2 presented monochrome seed coats of black color (Fig.2A), also seeds were observed with partially or completely yellow colorations. The distribution of fosets in these seeds was highly uniform (Fig.2A). In these seeds, the presence of apical or basal appendices was not common. In no case, basal horn was observed.
All dry seeds of PrJJ presented combined coloration: coffee and black (Fig.3A). The fosets were distributed uniformly in the majority of seeds (Fig.3A). In Fig.3B, the mucilaginous layer (exotesta) could be appreciated that surrounds the whole seeds; it was observed that this layer was removed in imbibed seeds, entirely or partially, by superficial contact. In these seeds, the presence of apical appendices was common, while the basal ones were present in a minor proportion.
It was very common to find seeds of PmFD with different tonalities of coffee color and without black color (Fig.4A). Towards the basal and apical parts the presence of fosets was not common in these seeds (Fig.4A). The presence of appendices was not typical for this accession being the one that had a minor number of appendices in seeds. In no case, basal horn was observed.
PmN was the only accession, which seeds, in dry state, presented orange color (Fig.5A); this coloration was present in all seeds of this accession, alone or combined with coffee and/or black color. In these seeds, the presence of appendices was more common for apical parts than for basal ones (Fig.5A). The fosets presented high uniformity on the seed surface (Fig.5A). The basal horn observed previously in PrJ1 and PrJJ was not observed in this accession. The accession with the highest average length of seeds (7.42 mm) was PmN (Tab.1) that presented values between 7.11 and 7.88 mm. Also, the latter accession had the highest values of width and thickness, presenting values between 4.12 and 4.7 mm, and 1.73 and 2.07 mm, respectively. The rest of the accessions had width of seeds between 3.38 and 4.38 mm; both extreme values were registered in the accession PmFD. The minimal thickness was reported for the seeds of accession PrJJ with 1.42 mm and the maximum one was reported for the seeds of accession PrJ2 with 1.86 mm. The accession PrJJ had very variable L/W values, very rounded (1.01) and also lengthened (1.86) seeds, with major tendency to be lengthened, since it presented the highest average value (1.73); in the rest of the accessions no value was minor than 1.56 (Tab.1). It is interesting that the seeds from the district El Roble (PrJ1, PrJ2, and PrJJ) had average L/W values higher than those in the seeds from the district La Mensura (PmFD and PmN) located at major altitude (Tab.1).
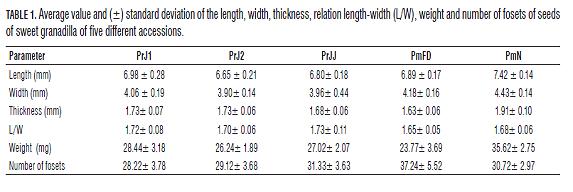
Meanwhile the mass of every seed in other accessions was between 12.6 (PmFD) and 33.0 mg (PrJ1), PmN presented seed mass that varied from 30.1 up to 39.6 mg, with an average of 35.62 mg (Tab.1). The number of fosets was very variable between the seeds of every accession, presenting a minimum of 21 fosets (PrJ1) and a maximum of 51 fosets (PmFD) (Tab.1). The number of fosets was least variable in the accession PmN (Tab.1).
Histological sections
One of the most importantant achievements of this research was the adaptation of the protocol which ollowed to obtain the quality of images observed in the histological cuts. The principal modification of the protocol used by Pérez-Cortéz et al. (2005) was to keep the blocks of parafine, with the samples, in a solution of 18,5% (half of the commercial concentration) of HCl by three days. After that, the blocks were washed with current water by 10 min to eliminate completely the acid, thus preventing the damage of the blades. Other important modification, which let a beter fijation of the cuts in the porta, was the application of the fijator Mayer albumen to the porta prior to set the cut on the porta. The application of the albumen was made at least 5 min before seting the cut, aming it culd dry. An important tip to maintain the tissue continuity was to use solid steel blades, because with thin disposable blades the results are not satisfactory by the vibration produced at contact with the hard seed coats.
With the adjustment of the protocol, the images were taken that are presented in Fig. 6; in this case, three layers of cells in the seed coats could be clearly differentiated.Seed coat layers result are originated from the integument development, it could be found the testa and tegmen divide in three well differenced layers: exo, meso and endo, each one, but is more comon to found a reduced number. In the sweet granadilla seed coat two most external layers did part of testa and represented the mesotesta and exotesta (Fig.6A, B and D). The most inner layer corresponds to the exotegmen.
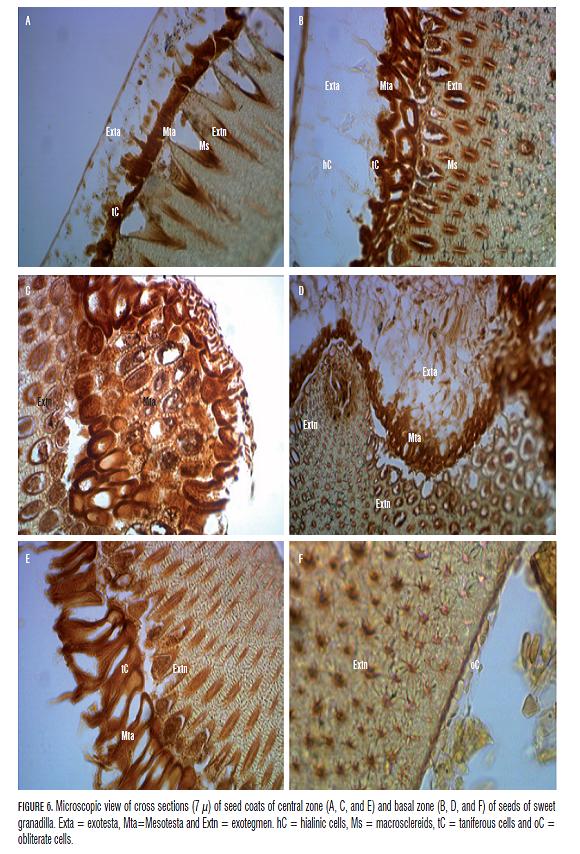
Fig.6A, B an C shows that the most external layer, exotesta, is composed by hialinic cells of mucilaginous type (Fig.6A, B and D) that shaped the brilliant and thin layer that could be observed in seeds of sweet granadilla at simple sight. This layer had been already reported in seeds of Passiflora sp. by Corner (1976). The thickness of this layer changed widely, and even, in some sections, it was observed that this layer was not continued in the medium part of the seeds (Fig.6 C and E). Towards the basal part this layer had a very uniform thickness (Fig. 6B). The only accession that presented this layer in all sections in continues form was PrJJ (Fig.6A). This layer had maximum thickness of 248 mkm in the medium zone of seeds and 610 mkm in the base, which indicates the highest development of this layer towards the part the tissues nearest to the embryo, as observed in Fig.6D.
The intermediate layer, mesotesta, was characterized with high fluctuation in size, since in the zone corresponding to fosets it had high thickness (Fig.6 C and E), whereas in the rest of the surface, this layer was narrower than the layer of macrosclereids (Fig.6A, B and D). The cells that composed this medium layer had changes in color, form and thickness of cell walls as well as variation in the number of cell layers. The color changed from clear coffee (Fig.6E) to very dark tones (Fig. 6B). This pigmentation could be related to the presence of tannins. The change in the thickness of cell walls could indicate the lignification of this layer in some zones (fosets) (Fig.6C), whereas different cells seemed to be taniferous, with thin cell walls and live citosol. The darkest colors and cellular walls of major thickness appeared towards the basal part (Fig.6B and D), such as a unique pattern for every accession was not observed.
In the part of the appendices, the internal layer could be increased up to 751 mkm, although in the zone of fosets, where its thickness diminished more, it could have up to 152 mkm. The opposite happened with the medium layer that thickened towards the fosets, where it could reach up to 378 mkm only in tissues, as observed in the Fig.6C, since this layer commonly did not overcome 152 mkm, with minimal thickness of 16 mkm, such as observed in Fig. 6E. The images show a layer of macrosclereids (Ms), in accordance with descriptions of exotegmen, which constitutes the most internal layer of cells. This one was characterized with most uniform thickness of three layers in all accessions, in the sections of the basal part as well as in the medium part of the seeds (Fig.6 A, B and E). This layer presented conical very elongated cells arranged in anticlinal form, with cell walls thickened towards the internal part of cytosol, which was arranged towards the most external layers that formed large spaces with the following layer.
These cells presented characteristics of high grade of lignification that leaved little space to the lumen (Fig.6 A). This characteristic was more observable in the medium part of the seeds than in the basal one, since, in the last one, the thickness was more uniform and the cytosol was more reduced in volume (Fig.6 B and D). The differences among the form of cells of the internal layer between the figures 2A, 2B and C were owed to the angle at which the seeds were sectioned. The multiple circles observed in B and C corresponded to the same layer, and these were reducing their size because there were several cross sections of cells with a conical form.
In the medium part of the seeds, the exotegmen presented gradual undulations; these undulations were those that formed the fosets (Fig.6 C), which is typical for the seed coats of seeds of family Passifloraceae (Corner, 1976). The internal layer presented changes among the central zone and the basal part of the seeds; in the basal part even, seemingly, a variation existed in the number of layers due to a high variation in size and form of cells of this layer, increasing towards the appendices of the seed coats (Fig.6 D).
The entire thickness of the seed coats was taken as the thickness of mesotesta plus the thickness of exotegmen, since the exotesta did not present continuity. The entire thickness fluctuated highly, being more constant in seed coats of seeds of PmN compared with other accessions. In average, the seed coats of seeds of sweet granadilla had a thickness among 235 and 475 mkm that could be increased up to 790 mkm in the region of appendices. The layer of obliterate cells was reported in seeds of other species of Passiflora, however, in this study, it was not observed; nevertheless, a section of basal part of seeds of accession PmFD showed a similar layer (Fig.6 F).
Discussion
The margin (entire) and the precence of central horn, previously reported by Pérez-Cortéz et al. (2002) were observed in all the seeds analized in this study, wich indicates it is a general characteristic for the species. Imbibed seeds of sweet granadilla presented a black color in the entire seed surface, which is in accordance with data reported by Pérez-Cortéz et al. (2002), who conducted a morphological analysis of seeds in 51 species of genus Passiflora and found that, in all cases, the seeds were monochrome. Nevertheless, in our study, every seed in dry state presented between one and three colors, being most typical the orange color for accession PmN that did not appear in seeds of any other accession.
The fosets in seeds of sweet granadilla independently of the accession were distributed uniformly on the central zone of seed surface, but it was common that these were not present in seed apex, base or margins; when presented in these zones, the fosets were of major length. This agrees with the data reported by Pérez-Cortéz et al. (2002) in the description of seeds of P. ligularis. For the species of genus Passiflora, the seminal characteristics could be considered as taxonomical characters, since these were demonstrated being a distinctive feature that allows identifying of species (Pérez-Cortéz et al., 2002).
An advantage of usage of these characteristics in taxonomy of genus Passiflora is a relative facility to study these, taking in account that, in other species, an electronic microscopy for the study of seed surface is needed, whereas, in case of seeds of Passiflora sp., one has managed to describe the seeds and create artificial keys only with the use of an optical microscope (Pérez-Cortéz et al., 2002).
A great part of the advantages that offer these seed characteristics owe to the differences that exist between the species, nevertheless its diagnostically value between accessions of the same species it is not so strict since, the seed coats of seeds is very similar. In spite of these limitations, for studied accessions, were clearly differed the seeds of accession PmN that presented a typical orange color, size and heavier weight than those of other accessions.
Characteristics of seed coats, such as the ornamentation or form of cells of every layer do not undergo high change with changing climatic conditions and, therefore, these present a high potential for identification of the species (Barthlott, 1984). Variables, such as weight and size, could change highly with climatic conditions of seed development (Copeland and McDonald, 2004). In this case, the seeds were coming from two farms with different agricultural practices and climate. In spite of this, the seeds of accession PmN presented major weight and size than the seeds of the rest of the accessions including PmFD that was collected in the same farm. The relation L/W was the highest for the accessions taken from the district El Roble (PrJ1, PrJ2, and PrJJ) that for those of the district La Mensura (PmFD and PmN), being very similar the values of two last accessions in spite of the high differences in size and weight that was observed between them.
The tissues were colored with fast green and safranin; nevertheless those final sections did not color sufficiently to observe the colorations typical of every layer. This allowed determining that pigmentation was darker in the basal tissues them in the tissues of medium zone of the seeds, but clear differences on this matter did not appear among the accessions. Pérez-Cortéz et al. (1995, 2005, 2009), in spite of having studied in depth the anatomy of seeds of Passiflora sp., could not realize the immersion in colorings due to the loss of the tissue. Therefore, results of our study suggest an important advance and contribution not only to seeds of family Passifloraceae, but also to other species with hard seed coats.
Studies on the anatomy of seed coats in species of genus Passiflora point out that these are constituted by one or two external layers of transparent cells, a medium layer of macrosclereids, and an internal layer of obliterate cells (Raju, 1955; Martín and Barkley, 1973; Corner, 1976; Pérez-Cortéz et al., 1995, 2005, 2009). The seed coats in species of genus Passiflora are formed by two teguments; the external tegument originates the most external layer of the seed coats and the multiplicative layer; from the internal tegument originates the layer of macrosclereids and the layer of obliterate cells (Raju, 1955). The layer of macrosclereids and the layer of obliterate cells have been identified in the anatomical profile of seed coats of the species in the literature; for this reason, it could be suggested that these layers are common for all species of this genus and are characteristically of the same one. Nevertheless, the observed images of the seed coat of sweet granadilla could be an exception to this characteristic. As it could be appreciated in the images of histological sections, in seeds of sweet granadilla there appeared a layer of macrosclereids, a layer of taniferous cells, and a layer of transparent cells, but the layer of obliterate cells reported for the seed coats of seeds of all species of Passiflora sp. earlier studied was not common (Pérez-Cortéz et al., 2002, 2009); only a section of the part basal of the seed of PmFD presented a discontinuous layer that could correspond to this layer. Raju (1955), in the anatomical description of the seed coats of Adenia venenata (Passifloraceae), did not report the presence of a layer of macrosclereids. It is necessary to highlight that it should be improved the methodology to obtain a better conservation of the soft tissues of seeds, which could affect the visualization of the most internal layers like oblitate cells.
The seeds of sweet granadilla presented an average thickness of 318 mkm, with values ranging between 218 and 474 mkm, with possibility of extending up to 790 mkm in the part of the appendices. P. alata and P. laurifolia thicker seed coats presented average thickness of 512 and 407 mkm, respectively. P. edulis, P. maliformis, P. quadrangularis, and P. nitida presented low values 123, 188, 267, and 260 mkm, respectively. This suggests that seed coats of sweet granadilla have average thickness in relation to other Passiflora sp. Between accessions of sweet granadilla the anatomical profile did not change a lot between accessions, although the seeds presented differences in weight and form between accessions. Between some related species, the anatomical profile of seed coats is uniform, whereas it varies between others, therefore, this characteristics could be of high utility in the taxonomical interpretation (Netolitzky, 1926; Werker, 1997; Pérez-Cortéz et al., 2009). In genus Passiflora, morpho-anatomical characterization of seed coats in the studied species of subgenus Passiflora, series Lobatae (P. gritensis, P. pallens, P. spectabilis, P. subpeltata), Dysosmia (P. foetida), Incarnatae (P. incarnata) and Imbricatae (P. sidiifolia) (Killip, 1938, cited by Pérez-Cortéz et al., 2009) shows that the anatomical profile of seed coats is specific and ornamentation of seed coats allows forming groups. Additionally to this, other authors emphasized the utility of seeds in the diagnostics of species of this genus (Deginani, 2001; Pérez-Cortéz et al., 1995, 2002, 2005, 2009), which is useful for granadilla too, but not for diferenciation between accessions of the same specie.
Conclusions
Ornamentation of seed coats and margin of seeds did not present differences between the analyzed accessions of sweet granadilla; in all cases, these presented falsifoveated seed coats with entire margin. The seeds of all accessions had from one up to three colors, and only in one accession, PmN, an orange color was observed.
The weight and size were highly variable between the seeds of every accession, except those belonging to PmN that presented major uniformity in these characters and major values of the same ones.
The softening of hard tissues with HCl and the use of fijator in the porta let the improve of the images achived in the cuts of granadilla seed coats.
The seed coats of seeds of sweet granadilla had three layers well differentiated in form and color of their cells, size and undulations.
The seed coats presented differences in thickness and cellular thickness between the medium part and the basal part of the seeds, having denser tissues towards the basal part. The layer of obliterate cells, commonly found in sections realized in seeds of other species of Passifloraceae, was not common for the sections of seeds of sweet granadilla.
Acknowledgments
The authors express their gratitude to the National University of Colombia, to Colciencias and to the Corporation Cepass-Huila for the financial and logistic support. Thanks are extended to the Laboratory of Microbiology of the Faculty of Agronomy, UNAL, principally to Mr. Wadith de León for his support in the adaptation of the protocols of microtomy and to Professor Xavier Marquínez for his help in the analysis of sections.
Literature cited
Barthlott, W. 1984. Microstructural features of seed surfaces. pp. 95-105. In: Heywood, V. and D. Moore (eds.). Current concepts in plant taxonomy. Academic Press, London. [ Links ]
Cardozo, H. 1988. Efecto de la escarificación y las dosis de ácido giberélico (GA3) en la germinación de semilla de curuba (Passiflora mollisima). Acta Biol. Colomb. 1(4), 127-132. [ Links ]
Corner, E.J.H. 1976. The seeds of dycotyledons. Vol. I. Cambridge University Press, Cambridge, Uk. [ Links ]
Copeland, L.O. and M.B. McDonald. 2004. Principles of seed science and technology. 4th ed. Kluwer Academic Publishers, Norwell, MA. [ Links ]
Cortéz, P.A. and S.A. Carmello-Guerreiro. 2008. Ontogeny and structure of the pericarp and the seed coat of Miconia albicans (Sw.) Triana (Melastomataceae) from "Cerrado", Brazil. Rev. Bras. Bot. 31(1), 71-79. [ Links ]
Deginani, N.B. 2001. Las especies argentinas del género Passiflora (Passifloraceae). Darwiniana 39, 43-129. [ Links ]
MacDougal, J.M. 1994. Revision of Passiflora subgenus Decaloba, section Pseudodysosmia (Passifloraceae). Syst. Bot. Monogr. 41, 1-146 [ Links ]
Martin, A.C. and W. Barkley. 1973. Seed identification manual. University of California Press, Berkeley, CA. [ Links ]
Moï se, J.A., S. Han, L. Gudynaite-Savitch, D.A. Johnson, and B.L.A. Miki. 2005. Seed coats: structure, development, composition, and biotechnology. In Vitro Cell. Dev. Biol. Plant 41, 620-644. [ Links ]
Netolitzky, F. 1926. Anatomie der Angiospermen Samen. Handbuch der Pfl anzenanatomie. Gebrüder Bornträger, Berlin. [ Links ]
Pérez-Cortéz, S., M. Escala, S. Tillett, and C. Sánchez. 1995. Estudio morfoanatómico de la cubierta seminal de Passiflora quadrangularis L. (Passifloraceae). Anales Bot. Agríc. 2, 25-29. [ Links ]
Pérez-Cortéz, S., S. Tillett, and M. Escala. 2002. Estudio morfológico de la semilla de 51 especies del género Passiflora L. Acta Bot. Venez. 25(1), 67-96. [ Links ]
Pérez-Cortéz, S., M. Escala, and S. Tillett. 2005. Anatomia de la cubierta seminal en ocho especies de Passiflora L. subgénero Passiflora. Acta Bot. Venez. 28(2), 337-348. [ Links ]
Pérez-Cortéz, S., M. Escala, and S. Tillett. 2009. Morfoanatomia de la cubierta seminal en siete especies de Passiflora L., subgénero Passiflora (Passifloraceae). Hoehnea 36(1), 131-137. [ Links ]
Raju, M.V.S. 1955. Embryology of the Passifloraceae. I. Gametogenesis and seed development of Passiflora calcarata Mast. Indian Bot. Soc. 35(1), 126-138. [ Links ]
Trujillo, H. 1983. Algunos aspectos sobre el cultivo de la granadilla. Rev. Esso Agríc. 11(22), 18-22. [ Links ]
Vanderplank, J. 1990. Passion flower and passion fruit. MIT Press, Cambridge, UK. [ Links ]
Werker, E. 1997. Seed anatomy. Gebrüder Bornträger, Berlin. [ Links ]

