










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.29 no.3 Bogotá Sept./Dec. 2011
PROPAGATION AND TISSUE CULTURE
Propagation of Chusquea tessellata (Munro) culms at different naphthaleneacetic acid concentrations and in different substrates
Propagación de culmos de Chusquea tessellata (Munro) a diferentes concentraciones de ácido naftalen-acético y en distintos sustratos
Jennyfer Insuasty-Torres1,3, Oscar Rojas-Zamora1, Orlando Vargas-Ríos1 and Camilo de los Ángeles Cárdenas2
1Grupo de Restauración Ecológica, Department of Biology, Faculty of Science, Universidad Nacional de Colombia. Bogota (Colombia).2Grupo de Restauración Ecológica, Universidad Antonio Nariño. Bogota (Colombia).
3Corresponding author. jinsuastyt@unal.edu.co
Received for publication: 22 February, 2011. Accepted for publication: 2 November, 2011.
ABSTRACT
Chusquea tessellata is native specie of Colombian wet páramos. Despite the ecological importance of this ecosystem, production activities have altered the structure of the C. tessellata patches. The aim of this study was to evaluate vegetative propagation from sections of culm stimulating their rooting, to have a lot of material for transplantation and with high survival. We tested 56 treatments with three combined factors: phytohormone NAA concentration, type of substrate and type of culm section. After three months we assessed the frequency and number of roots and vegetative shoots. The results suggest that the frequency and quantity of roots and shoots are affected mainly by the type of culm section, the best results is the basal sections, because these sections of culm have the morphological characteristics that allow successful propagation. Shoot production is also affected by interactions between other factors, substrate and NAA concentration.
Key words: bamboos, páramos, tropics, rooting, phytohormones.
RESUMEN
Chusquea tessellata es una especie nativa de los páramos húmedos de Colombia. A pesar de la importancia ecológica de este ecosistema, las actividades productivas como la ganadería han alterado la estructura de los parches de C. tessellata. El objetivo de este estudio fue evaluar la propagación vegetativa a partir de secciones de culmo estimulando su enraizamiento, con el fin de tener gran cantidad de material para trasplantar y con altos niveles de supervivencia. Se probaron 56 tratamientos con tres factores combinados: concentración de fitohormona ANA, tipo de sustrato y tipo de sección de culmo. Después de tres meses se evaluó la frecuencia y número de raíces y brotes vegetativos. Los resultados sugieren que la frecuencia y cantidad de raíces y brotes están afectadas principalmente por el tipo de sección de culmo, los mejores resultados se presentaron con las secciones basales, puesto que estas secciones de culmo tienen las características morfológicas que permiten una exitosa propagación. La producción de brotes está afectada también por las interacciones entre los otros factores, sustrato y concentración de ANA.
Palabras clave: bambúes, páramos, trópicos, enraizamiento, fitohormonas.
Introduction
Colombia has about 21 species of the genus Chusquea Kunth, distributed from lowland to high altitude regions. In particular, the following species have been reported above 3,000 m a.s.l.; Chusquea tessellata, Chusquea angustifolia, Chusquea lehmannii, Chusquea scandens, Chusquea serrulata, Chusquea spadicea and Chusquea spencei (Instituto de Ciencias Naturales, 2008). These species are typical from understory high mountain tropical forests, although they are also common in areas of open vegetation as the páramos of Colombia, Venezuela, Ecuador, Peru and Costa Rica (Clark, 1992; Judziewicz et al., 1999).
In Colombian páramos, atmospherically wet and poorly drained soils is common to find large patches or populations of C. tessellata (Munro) as an important floristic element; that is why Cleef (1981) classified the Andean bamboo associations as bamboo páramos referring to areas with abundant rainfall and high relative humidity where it is common to find C. tessellata. These areas are also characterized by the presence of lakes and water reservoirs that regulate the hydrological cycle and are a permanent source of drinking water for cities, which is related to their ecological, economic and social importance. Despite this fact, many of the wet páramo areas are used primarily for livestock, which causes changes in the structure of the patches of C. tessellata due to biomass removal and the fragmentation of clump (Vargas et al., 2002; Cárdenas- Arévalo and Vargas, 2008).
Taking into account the consequences of disturbances associated with livestock and ecological and social importance of páramos, the ecological restoration is a process and an opportunity to restore some areas altered by this profitable activity. Ecological restoration processes are intended to assist the recovery of an ecosystem that has been degraded, damaged or destroyed (SER, 2004). To carry this out, it is essential to obtain and handle vegetable material propagated by seed or vegetatively.
Vegetative propagation in bamboos is a life history traits and reproductive and expansion strategy of great importance; the growth form of these plants is given by the branching of the rhizomes and the formation of new shoots, which grow and become culms or branches, which as well form clump called ramets if they are vegetative units or genets if they are units with different genetic identity (Mc- Clure, 1966; Sodestrom and Calderón, 1978; Clark, 1989; Briske and Derner, 1998; Makita et al., 1998; Judziewicz, 1999). In the case of C. tessellata it is known that vegetative propagation by rhizome sprouts allowed it to be highly competitive, expand its coverage as well as expand quickly, which favors its dominance in the Colombian wet páramos. However a permanent soil seed bank, seedlings or young plants have not been found; these characteristics can lead into local extinction in areas with a high degree of alteration. In addition to this, burning and grazing affect these species by removal of biomass and fragmentation due to livestock stomping, and can be displace by Lachemilla orbiculata, a dominant ground herb of wet páramos that has been transformed to support livestock (Vargas et al., 2002; Cárdenas-Arévalo and Vargas, 2008).
In preliminary tests, one result was that the culms extracted from a group of plants with 1 m or above and relocated in places with similar conditions to the original did not survive or produce new roots or shoots (unpublished data). It confirmed the importance of carrying out research on stimulation of rooting culms, to propagate the species vegetatively, gather lots of plants that can be transplanted, and that have the ability to survive in areas altered by human activities in which C. tessellata naturally would be. To date there have been no studies on vegetative propagation of this species of bamboo, one reason is that their thin culms do not satisfy the needs for handcrafting or industrial use, fact that also decreases its commercial value. For other species of bamboo in the american tropic, protocols or guidelines have been developed for the vegetative propagation through methods as "chusquines", culm sections and in vitro culture; an example is the guadua (Guadua angustifolia), whose uses are primarily related to construction, reforestation and protection of riversides and lake shores (Corpocaldas, 1983; Hidalgo et al., 1992; Londoño, 1998; Giraldo and Sabogal, 1999; Marulanda et al., 2002; Rodríguez, 2003), and a high mountain tropical bamboo typical from forests (C. scandens) (DAMA, 2000).
However, further research is needed focusing on each specie of interest, since the morphological and physiological characteristics of each bamboo may generate different responses in the survival, production of shoots and roots. The phytohormone of the auxin group, has been used widely to promote rooting of cut stems or other parts of plants (Hartmann et al., 2002), these phytohormones control a series of processes in the growth and development of the plants, such as embryogenesis, apical dominance, stem elongation, root development and tropism (Berleth and Sachs, 2001).
The goal of this study is to assess vegetative propagation from sections of culms, stimulating rooting and shoot production of C. tessellata by using naphthalene acetic acid (NAA), a synthetic auxin that is available in the market. Additionally, two maintenance substrates, watery substrate and earth were tested, to propose protocols that make it possible to have enough material for transplantation and with high rates of survival, which will increase the success of ecological restoration programs of disturbed páramo areas.
Materials and methods
Location and plant material
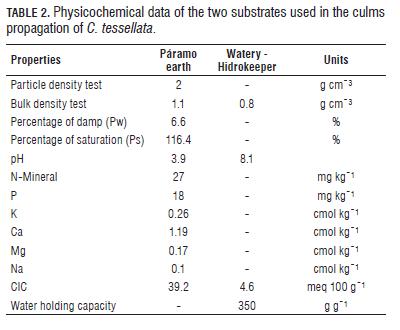
The research was performed under greenhouse conditions in the sector of Lagunas de Siecha, northwest Chingaza National Natural Park (PNN) - Colombia, located at 4°46 '10'' N, 73°51'53.9'' W and 3,424 m a.s.l. Because of high solar radiation in the páramo and that at noon the temperature can reach 30°C, this greenhouse was covered with UV plastic on the roof and in the lower half of the sides, while the top half of the sides were covered with fine mesh veil so that the temperature conditions did not differ much from those present in the páramo, taking into account that the temperature in the greenhouses in which this parameter is not controlled, the temperature is higher than in the outside. Inside the greenhouse a datalogger was installed, through which the temperature and relative humidity during the day were recorded every two hours, from 8:00 am to 6:00 pm, and 13.64°C average (SE 0.16 ) and 74.33% RH; and during the night from 8:00 pm to 6:00 am, with 6.16°C average temperature (SE 0.08) and 99.55% RH.
The data used in this research was collected between January and April 2009. We collected 500 culms of C. tessellata from a community characterized by C. tessellata and Espeletia killipii as dominant species. The culms were extracted with a piece of rhizome; these were approximately 1.5 m long and 1 to 2 cm in diameter. Subsequently the culms were taken to the greenhouse and cut straight and so obtain seven types of culm section, these culm types are; two of the basal or root (B1, B3), two of the middle (M1, M3), two of the distal (D1, D3) and one with the whole culm (E). None of the whole culms or culm sections were exposed to any kind of fungicide during the experiments.
The experiment consisted in evaluating the effect of three factors and their interaction: the culm sections, NAA phytohormone concentration and substrate type (Tab.1), giving us the application of 56 treatments; each treatment was applied in 15 culm sections or replicates. NAA concentration and time of exposure to this phytohormone were made based on the method used by Murillo and Montiel (1998a).
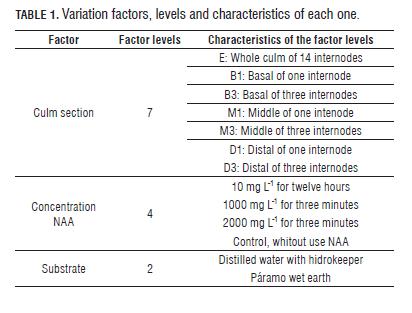
Culm sections that were planted in the páramo earth kept their moisture by dripping irrigation (Tab.2). For sections of culm that remained in the watery substrate, distilled water was used with hidrokeeper, a copolymer of acrylamide and potassium acrylate (> 85%), in order to simulate flooded soil conditions where C. tessellata grows, as the edges of lakes or areas of lower slope in the valleys.
Data collection and statistical analysis
Three months after we applied the treatment, we counted the number of roots and the number of shoots in each culm section, and assessed the survival of these by verifying the presence of active photosynthetic tissues. For each of the treatments, we estimated the rooting frequency and the number of culm sections with at least a new root; also we estimated the shoots frequency in each treatment and the number of culm sections with at least one new shoot. We analyzed the frequency of rooting and shoot production using generalized linear models (GLM) with a binomial probability model (rooted/non-rooted - shoot/non shoot).
As factors of variation we used NAA concentration, the type of culm section and the substrate. We used Wald chi-squared statistic, through the maximum likelihood method, to test the effect of the factors and their interaction on the production of shoots and rooting. The number of roots and shoots produced was analyzed only with culm sections that had at least one root or shoot, that is, we excluded all events that did not produce roots or shoots. We used a GLM (Poisson regression with log-link), and Wald chi-squared statistic through the maximum likelihood method, to test the effect of the factors and their interaction on the number of roots or shoots produced. The differences between the levels of the main factors were analyzed using Bonferroni pairwise comparisons. All analysis were carried out using SPSS 19 for Windows (SPSS, Inc. Chicago, IL).
Results and discussion
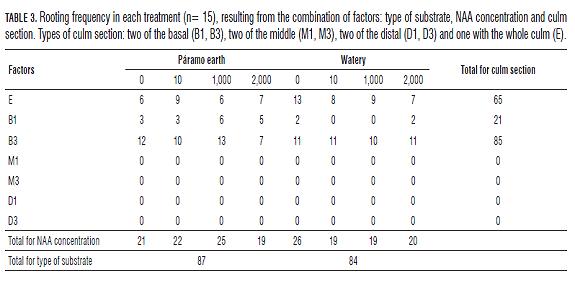
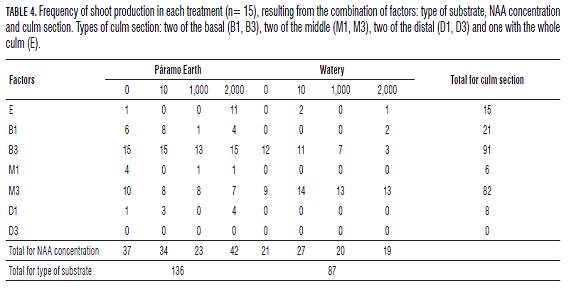
The middle and distal culm sections (D1, D3, M1 and M3) had not rooting events (Tab.3), neither the section D3 had shoot production (Tab.4), that is why we excluded these sections from the respective analysis.
Rooting
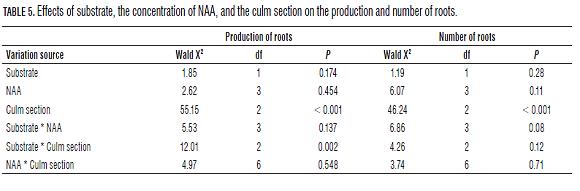
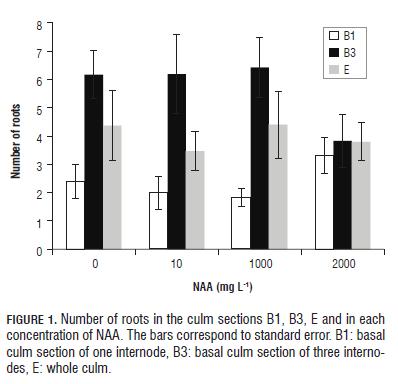
The culms rooting C. tessellata is affected by the type of culm section and the interaction between this factor and the type of substrate, but is not affected by the concentration of NAA, nor the substrate in which the sections are kept or other interactions between factors. However, the amount of roots produced is affected only by the type culm section (Tab.5). The number of roots produced using whole culm sections (E) differs from the number of roots produced using basal culm sections of one internode (B1) (Bonferroni P=0.001) and from the basal culm sections of three internodes (B3) (Bonferroni P=0.001); always being higher the amount of roots produced using the B3 section, and the B1 culm section producing the lowest amount of roots. In the Fig.1, we can see that despite there are no differences between the concentrations of NAA, the 2,000 mg L-1 concentration reduced the number of roots in the basal sections (B1 and B3); other concentrations have a similar effect on this variable.
Shoot production
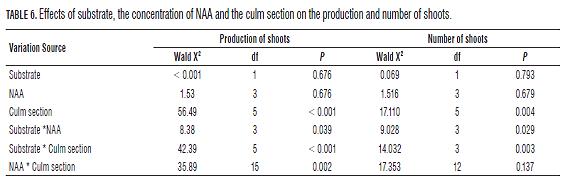
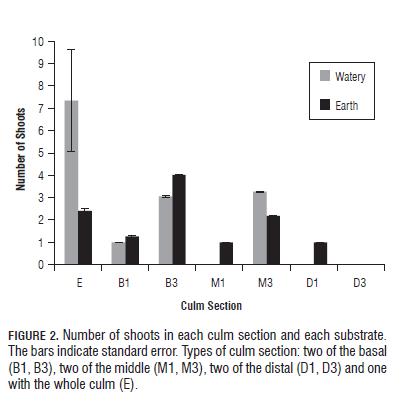
The production of shoots in culms of C. tessellata is affected by the type of culm section used, and the different interactions between pairs of main factors. In the culms that showed shoots production, the variation in the number of shoots produced depended on the section type used, the interaction between the substrate and the culm section type, and the interaction between the substrate and the concentration of NAA (Tab.6). The sections that showed the highest number of shoots corresponded to B3, M3 and E (Fig.2), but only B1 and B3 differed significantly from each other (Bonferroni p= 0.032).
Vegetative propagation of C. tessellata culms Many restoration programs have approached in the use of trees and do not take into account life forms as epiphytes, lianas and shrubs, although they play an important role in natural regeneration (Rodrigues et al., 2009). However, one reason for not using these species is the lack of knowledge about the propagation of other life forms, in this way we intend to contribute to the knowledge of the propagation of C. tessellata.
The different NAA concentrations tested do not affect the rooting frequency of the culms neither the amount of roots produced. Taking into account that 2,000 mg L-1 NAA concentration produced the lowest amount of roots, that is to say the concentration has a negative effect on root production; we suggest that there is some inhibition in cell division for the root formation. It was observed that high auxin concentrations act as inhibitors of cell division, whether phytohormone is from endogenous and exogenous origin or only exogenous. The first case occurs because the plant parts after cutting keep the property of synthesizing auxins, that is to say they have an endogenous concentration of auxines, and therefore when exogenous auxins are added, results in a supra-optimal concentration of this phytohormone in plant tissues, causing a negative effect on rooting. The second case occurs in some plant species when they decrease the production of roots due to the addition of high concentrations of exogenous auxin (Moura-Costa and Lundoh, 1994; Blakesley et al., 1991).
Additionally, the culm section type is a variable that affects rooting capacity and number of roots produced by increasing them, because the parental plant material influences directly these capabilities. It is important to note that the basal parts of the culms (B1 and B3) are composed mainly of rhizome, that is to say modified stems that grow horizontally and emit roots and shoots. McClure (1967) and Murillo and Montiel (1998b) carried out tests of propagation in the specie of bamboo Gigantochloa apus, and emphasized the fact that the number of root primordia decreases differentially from the base to the distal part, and there is a natural latency in the middle of the culms; that is why basal culm sections produced roots, unlike middle and distal sections of the culms. For the production of shoots and number of shoots it is important to consider the type of culm section, and the interactions between factors: section type and substrate, substrate and concentration of NAA.
Based on the results it is not recommended the use of NAA as a stimulant for rooting or shoot production. The inverse relationship between the phytohormone concentration with the number of roots is similar to results found in studies of vegetative propagation in species of tropical bamboos such as Bambusa vulgaris, and temperate bamboos such as Dendrocalamus sp. and Phyllostachys sp.; in which high concentrations of phytohormone show the lowest scores for the production of roots, although these studies are also associated with decreased production of shoots (e.g. McClure, 1973; Hasan, 1980; Banik, 1980; Murillo and Montiel, 1998a). This result is important for the propagation protocol approach of C. tessellata, taking into account that the implementation of restoration projects, the use of simple and inexpensive treatments show positive results, meaning restoration processes that gain time and money.
Despite there are not any studies or other information about vegetative propagation from other species of the Chusquea genus, they share two features in the analysis for the successful propagation of tropical bamboo species. The first is the direct relationship between parental material and shoot production, emphasizing that some parts of the plant have more meristematic ability and nutrient storage; this is why different types of culm section of various ages, produce different amounts of shoots. Hartmann et al. (2002), explain this feature by setting two fundamental aspects for a cutting to propagate vegetatively and grow into a complete plant: the totipotency and cell differentiation. In the propagation of C. tessellata is noticeable the effect that different types of culm section has on production and number of roots and shoots, particularly highlighting the basal sections of three internodes (B3) and the whole culm with rhizome (E), since they are the sections that increased frequency and quantity of roots and shoots production.
Ruíz and Montiel (1998), assessed vegetative propagation taking into account the type of culm sections and the age of culm groups of G. chacoensis in Costa Rica; their results showed that there is less production of shoots in basal sections, unlike what happens to middle and distal sections. Additionally they reported that one year-old culms, produced more shoots than the three year-old ones. In the current research, the age of culms of C. tessellata was not a factor of variation, since it sought to control the effect of age by choosing similar culm length and diameter. Although one cannot infer the age of the culms, is a clear relationship between the different parts of the culm and the production of shoots and roots. In C. tessellata the important elements are the morphological characteristics of culms; the basal sections display a dense set of internodes and root meristem lots, which are the primordia of the rhizome with the capacity of forming adventitious roots and shoots, unlike what happens in the middle and distal sections whose meristematic differentiation is mainly into leaves and branches. Along with the lack of effect by the different concentrations of NAA, and the good results associated with the non-use of this phytohormone, we can suggest that cells of root meristem contain adequate levels of endogenous auxins of plant growth regulators, able to stimulate the rooting of culm and promote cell division and elongation, among others.
The second feature to consider is the most suitable type of substrate for rooting and shoot production. Particularly it is to be highlighted the importance of providing a substrate with appropriate physicochemical conditions for the development of roots and shoots; the substrate must contain essential nutrients as P and Ca for root growth (Hartmann et al., 2002), it should also keep moisture and promote aeration.
Several studies on vegetative propagation have tested different types of substrates with variations in the proportion of earth, rice husks and sand, however it is not common to use an aqueous substrate for this type of propagation by cuttings, but as mentioned above, in the case of propagation in C. tessellata was intended to simulate the condition of poorly drained soils, characteristic of the places where it lives.
In the results of this research, the type of substrate affects the frequency of rooting and production and number of shoots when interacting with the section type and concentration of NAA. It should be highlighted that in logistics terms, the aqueous substrate is more difficult to manage and control, since it is an environment that requires constant attention to avoid the total evaporation of water, and growth of algae that contribute to the rottening of the culms. Additionally this substrate lacks of essential nutrients for plant growth, with limited ventilation and poor physical stability. By contrast, the physicochemical conditions of moist soil may be more suitable for the development of roots and shoots: it has some nutrients (N, P, K, Ca, Mg), a porous structure that allows anchoring and stability for culms, as well as allowing enough aeration and moisture. These characteristics when interacting with culm section type and NAA concentration could have generated adequate conditions to develop roots and shoots, enhancing the turgor in the plant and healthier tissues and since no signs of rottening or degradation by fungi or bacteria were observed.
Some guides have been prepared for the vegetative propagation of high Andean bamboo species as C. scandens, specie that is used for basin protection and soil retention. These protocols describe the cutting of culms and direct seeding in wet areas (DAMA, 2000), suggesting the easy propagation of these species. However, in this study was evident the ineffectiveness for rooting and shoot production of C. tessellata, aspects that may be associated with life history traits typical of the species and the morphological and physiological characteristics that determine their growth and development in areas of wet páramo.
Conclusions
The culm propagation of C. tessellata, in terms of production and number of roots and shoots, was mainly influenced by the type of culm section, taking into account this factor is directly related to the morphological characteristics of the culms of C. tessellata and therefore with the intrinsic capacity to produce roots and shoots.
The results show that concentrations of NAA, 10; 1,000 and 2,000 mg L-1 are not effective for inducing roots in C. tessellata. Instead, culms or sections from these that were not treated with NAA and that were kept in moist soil had higher amount of roots and shoots. The maintenance of culms in moist soil is a suitable method for the development of root and shoot production, as well as allowing greater control in the process of vegetative propagation of this species of bamboo. The physicochemical conditions of this substrate enhance this process by keeping nutrients available in the environment, stabilizing the plant material and indirectly avoid contamination by fungi and bacteria.
We recommend to use the basal culm section with three or more internodes and the whole culms, it is essential to extract them carefully so that they can keep the rhizome part; since it is the part of the plant that contains meristems whose cells can differentiate into roots and shoots. In order to expand knowledge about the requirements for the propagation of C. tessellata it is important to carry out experiments that test other concentrations and mode of application of NAA, as well as testing other auxins such as indole acetic acid (IAA), phenyl-acetic acid (PAA) or indole-butyric acid (IBA).
Acknowledgments
We would like to thank to Department of Administrative Science, Technology and Innovation – COLCIENCIAS, for financing the project "Strategies for the Ecological Restoration of the Andean páramos" which was developed within this research. We also thank to the National Natural Parks Unit, the PNN Chingaza for giving us the research permit, especially to Carlos Lora, to park officials Fredy Avellaneda, Alirio Rodriguez and Fredy Garcia; the members of the Foundation Suasie and the rural community of the village Paso Hondo, Guasca for their valuable collaboration in the field; to David Escobar for their assistance in reviewing and translating the text. A special thank to the Biology Department of the Universidad Nacional de Colombia and the "Grupo de Restauración Ecológica" especially to Pilar Gómez for her support in various tasks during the project. To Maria Argenis Gomez Bonilla and the Research Group "Biología de Organismos Tropicales» for their academic support.
Literature cited
Banik, R.L. 1980. Propagation of bamboos by clonal methods and by seeds. pp. 139-150. In: Lessard, G. and A. Chouinard (eds.). Bamboo research in Asia. International Development Research Center, Otawa. [ Links ]
Blakesley, D., G.D. Weston, and J.F. Hall. 1991. The role of endogenous auxin in root initiation. Plant Growth Regulat. 10(4), 341-353. [ Links ]
Briske, D.D. and J.D. Derner. 1998. Clonal biology of caespitose grasses. pp. 106-135. In: Cheplick, G.P. (ed.). Populations biology of grasses. Cambridge University Press, Cambridge, UK. [ Links ]
Berleth, T. and T. Sachs. 2001. Plant morphogenesis: long-distance coordination and local patterning. Curr. Opin. Plant Biol. 4, 57-62. [ Links ]
Cárdenas-Arévalo, G. and O. Vargas. 2008. Rasgos de historia de vida de especies en una comunidad vegetal alterada en un páramo húmedo (Parque Nacional Natural Chingaza). Caldasia 30(2), 245-264. [ Links ]
Clark, L.G. 1989. Systematics of Chusquea section Swallenochloa, section Verticillatae, section Serpentes and section Longifoliae (Poaceae: Bambusoideae). Syst. Bot. Monogr. 27, 1-127. [ Links ]
Clark, L.G. 1992. Chusquea sect. Swallenochloa (Poaceae: Bambusoideae) and allies in Brazil. Brittonia 44(4), 387-422. [ Links ]
Cleef, A.M. 1981. The vegetation of the Paramos of the Colombian Cordillera Oriental. Ph.D. thesis, Utrecht University, Utrecht, The Netherlands. [ Links ]
Corpocaldas, Corporación Autónoma Regional del Caldas. 1983. La guadua, planta emblema de Caldas: propagación, manejo y utilización. Caldas, Colombia. [ Links ]
DAMA. 2000. Protocolo distrital de restauración ecológica. Departamento Técnico Administrativo de Medio Ambiente, Bogotá. [ Links ]
Giraldo, E. and A. Sabogal. 1999. Una alternativa sostenible: La guadua, técnicas de cultivo y manejo. Corporación Autónoma Regional del Quindío, Quindío, Colombia. [ Links ]
Instituto de Ciencias Naturales. 2008. Colecciones científicas en línea del Instituto de Ciencias Naturales – Herbario Nacional Colombiano COL., http://www.biovirtual.unal.edu.co/ colecciones/datos/herbario/consultasHerbario.jsp; consulted: july, 2011. [ Links ]
Hartmann, H.T., D.E. Kester, F.T. Davies, Jr., and R.L. Geneve. 2002. Plant propagation, principles and practices. 7th ed. Prentice Hall, Saddle River, NJ. [ Links ]
Hasan, S.M. 1980. Lesson from partial studies on the propagation of bamboos. pp. 131-138. En: Lessard G. and A. Chouinard (eds.). Bamboo research in Asia. International Development Research Center, Otawa. [ Links ]
Hidalgo, O. 1992. Tipos de Bambúes y métodos de cultivo. pp. 1-11. In: Memorias II Simposio Latinoamericano de Bambú. Universidad Laica Vicente Rocafuerte, Guayaquil. [ Links ]
Judziewicz, E.J., L.G. Clark, X. Londoño, and M.J. Stern. 1999. American bamboos. Smithsonian Institution Press, Washington DC. [ Links ]
Londoño, X. 1998. A decade of observations of a Guadua angustifolia plantation in Colombia. J. Amer. Bamboo Soc. 12(1), 37-43. [ Links ]
Makita, A. 1998. Populations dynamics in the regeneration process of monocarpic dwaft bamboos, Sasa species. pp. 313-332. In: Cheplick, G.P. (ed). Populations biology of grasses. Cambridge University Press, Cambridge, UK. [ Links ]
Marulanda, M.L., M. Carvajalino, C. Vargas, and X. Londoño. 2002. La biotecnología aplicada al estudio y aprovechamiento de la Guadua. pp. 1-5. In: Seminario - Taller Avances en la Investigación sobre Guadua. Pereira, Colombia. [ Links ]
McClure, F.A. 1966. The bamboos: a fresh perspective. Harvard University Press, Cambridge, UK. [ Links ]
McClure, F.A. 1967. The bamboos. Harvard University Press, Cambridge, UK. [ Links ]
McClure, F.A. 1973. Genera of bamboos native to the new world (Gramineae: Bambusoideae). Smithsonian Contributions to Botany No. 9. Smithsonian Institution Press, Washington DC. [ Links ]
Moura-Costa, P.H. and L. Lundoh. 1994. The effects of auxins (IBA, NAA and 2,4-D) on rooting of Dryobalanops lanceolata (Kapur – Dipterocarpaceae) cuttings. J. Trop. For. Sci. 7(2), 338-340. [ Links ]
Murillo, R.L. and M. Montiel. 1998a. Efecto del ácido naftalenacético en el enraizamiento de diferentes secciones del culmo de Bambusa vulgaris. pp. 61-64. In: Montiel-L., M. (ed). Cultivo y uso del bambú en el neotrópico. Universidad de Costa Rica, San José. [ Links ]
Murillo, R.L. and M. Montiel. 1998b. Efecto de la edad y la posición de las yemas en el culmo, en la reproducción vegetativa de Bambusa vulgaris y Gigantochloa apus. pp. 28-35. In: Montiel-L., M. (ed.). Cultivo y uso del bambú en el neotrópico. Universidad de Costa Rica, San José. [ Links ]
Rodrigues, R.R., R.A.F. Lima, S. Gandolfi, and A.G. Nave. 2009. On the restoration of high diversity forests: 30 years of experience in the Brazilian Atlantic Forest. Biol. Conserv. 142(6), 1242-1255. [ Links ]
Rodríguez, N. 2003. Propagación in vitro de Guadua angustifolia Kunth (bambú), importancia y ventajas comparativas. pp. 46. In: Memorias III Seminario Internacional del Bambú. Programa Agricultura Tropical Sostenible; Ministerio del Ambiente y de los Recursos Naturales; Fundación Polar; Gobernación del Estado Yaracuy; Fundación para la Investigación Agrícola (DANAC). San Felipe, Venezuela. [ Links ]
Ruíz, A.I. and M. Montiel. 1998. Efecto de la edad y sección del culmo en la propagación asexual de Guadua chacoensis (Rojas Acosta) Londoño y Peterson y su respuesta a la cobertura con Pennisetumpurpureum var. King Grass (Poaceae). pp. 28-35. In: Montiel-L., M. (ed.). Cultivo y uso del bambú en el neotrópico. Universidad de Costa Rica, San José. [ Links ]
SER, Society for Ecological Restoration International. 2004. The SER primer on ecological restoration. In: http://www.ser.org; consulted: July, 2011. [ Links ]
Sodestrom, T.R. and C.E. Calderón. 1978. The species of Chusquea (Poaceae: Bambusoideae) with Verticillate Buds. Brittonia 30(2), 154-164. [ Links ]
Vargas, O., J. Premauer, and C.dlA. Cárdenas. 2002. Efecto del pastoreo sobre la estructura de la vegetación en un páramo húmedo de Colombia. Ecotropicos 15(1), 35-50. [ Links ]

