




![Biological nitrogen fixation by Rhizobium sp. native gliricidia (Gliricidia sepium [Jacq.] Kunth ex Walp.) under greenhouse conditions](/img/en/next.gif)





Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.29 no.3 Bogotá Sept./Dec. 2011
SOILS, FERTILIZATION AND MANAGEMENT OF WATER
Effect of land use on the density of nitrifying and denitrifying bacteria in the Colombian Coffee Region
Efecto del uso de suelo sobre la densidad de bacterias nitrificantes y desnitrificantes en la Ecorregión Cafetera Colombiana
Victoria E. Vallejo1,María M. Gómez1,Ana M. Cubillos1 and Fabio Roldán1,2
1Sanitation Unit and Environmental Biotechnology (USBA), Department of Biology, Faculty of Science, Pontificia Universidad Javeriana. Bogota (Colombia). 2Corresponding author. fabio.roldan@javeriana.edu.co Received for publication: 24 August, 2010. Accepted for publication: 2 November, 2011.ABSTRACT
Soil microbial communities involved in the cycling of nitrogen (N) are essential to maintaining and improving soil fertility, productivity and functionality of natural and agricultural ecosystems. However, some compounds generated during the metabolic processes performed by nitrifying (NB) and denitrifying (DB) bacteria are associated with the production of greenhouse gases, groundwater pollution and acidification. Therefore, the study of these bacteria is essential for economic and environmental sustainability. This study evaluated the effect of different land uses in two river basins (La Vieja and Otún) on NB and DB densities. Two sampling events (SE) were conducted by selecting the most representative land uses. Physicochemical (T °, pH, moisture and nitrate) and microbiological properties (NB and DB densities) were evaluated. In both SEs, significantly higher densities of NB and DB were observed in the land uses: pasture, guadua (DB only) and unshaded coffee (La Vieja) and onion (Otún). These land uses, excluding guadua, are dependent on nitrogen fertilizers, which together with the activities of grazing livestock on pastures may lead to greater availability of substrates for the NB. The use of agricultural machinery and overgrazing in pasture and onion uses generate compacted soil and other physical disturbances, encouraging the growth of DB. Forests had the lowest densities of NB and DB possibly due to a reduced availability of N and the releasing of allelopathic compounds from certain plants. Finally, the densities of ammonium-oxidizing bacteria had the greatest differences between the land uses evaluated, demonstrating its high sensitivity to agricultural management practices and livestock. We suggest that changes in the abundance of this community could serve as a relevant and cost-effective bioindicator for soil monitoring.
Key words: nitrifying bacteria, denitrifying bacteria, land use, Colombian coffee region, ammonia oxidizing bacteria.
RESUMEN
Las comunidades microbianas edáficas involucradas en el ciclaje de nitrógeno (N) son fundamentales para el mantenimiento y mejoramiento de la fertilidad, productividad y funcionalidad de los ecosistemas naturales y agrícolas. Sin embargo, algunos compuestos generados durante los procesos realizados por bacterias nitrificantes (BN) y desnitrificantes (BD), se asocian con la producción de gases efecto invernadero, contaminación de aguas subterráneas y acidificación. Por lo tanto, el estudio de estas bacterias resulta esencial para la sostenibilidad económica y ambiental. El presente estudio evaluó el efecto de diferentes usos de suelo en dos cuencas hidrográficas (la Vieja y Otún) sobre la densidad de BN y BD. Se realizaron dos eventos de muestreo (EM) seleccionando los usos de suelos más representativos. Se evaluaron propiedades fisicoquímicas (T°, pH, porcentaje de humedad y nitratos) y microbiológicas (densidad de BN y BD-Número más probable). En los dos EM se observaron densidades significativamente mayores de BN y BD en los usos de pastizal, guadual (solo BD) y cafetal sin sombrío (la Vieja), así como en cebolla (Otún). Estos usos de suelo, excluyendo guadual, son dependientes de fertilizantes nitrogenados, lo cual en conjunto con las actividades de pastoreo del ganado en los pastizales, podrían generar una mayor disponibilidad de sustratos para las BN. El uso de maquinaria agrícola y sobrepastoreo en cebolla y pastizal generan disturbios físicos en el suelo y lo compactan favoreciendo el crecimiento de BD. Los bosques presentaron las densidades más bajas de BN y BD posiblemente por una menor disponibilidad de N y la liberación de compuestos alelopáticos. Finalmente, las densidades de bacterias oxidadoras de amonio fueron quienes presentaron mayores diferencias entre los diferentes usos de suelo evaluados, demostrando su alta sensibilidad frente a prácticas de manejo agrícola y pecuario.
Palabras clave: bacterias nitrificantes, bacterias desnitrificantes, usos de suelo, ecorregión cafetera colombiana, bacterias oxidadoras de amonio.
Introduction
In recent years, there have been increasing concerns about the consequences of land cover change and its effects on the earth's ecosystems. For example, conventional farming systems share many characteristics, such as: large capital investments with rapid technological innovation, largescale farms with monoculture agricultural practices, and extensive use of external chemical inputs (e.g., pesticides and fertilizers) to increase agricultural productivity (Matson et al., 1997). However, high demand and dependence of chemical inputs has caused economic and environmental problems (Hurni et al., 2008), which have reduce fertility and productive capacity in soil as a result of physical (erosion and compaction), chemical (acidification, nutrient loss and pollution) and biological degradation (biodiversity loss) (Hunter, 1996; Girvan et al., 2003).
Agricultural management practices (e.g., tillage, organic amendment addition, chemical fertilization, overgrazing, cropping rotation, and irrigation) can strongly influence the size, composition, and activity of the microbial community in soils. Moreover, a modification of the soil microbiota may in turn affect soil processes, providing a positive or negative feedback on plant productivity (Feng et al., 2003; Parfitt et al., 2010). Previous studies have established that changes in soil microbial communities are caused by alterations in the physicochemical characteristics of the soil (e.g., organic C, nutrients, bulk density, pH and moisture), and in the quantity, quality and distribution of crop residues as a result of implemented agricultural and livestock management practices (Hayden et al., 2010; Kong et al., 2010).
To assess the impact of land use on soil microorganisms, two approaches can be used. In the first approach, total microbial community is studied, but diversity changes on this community may not generate effects on ecosystem processes due to a high functional redundancy (Allison and Martiny, 2008; Nyberg et al., 2006). The second approach evaluate microbial functional groups which are involved in N cycling, where functional redundancy is lower, and therefore, any diversity change would have a significant impact on soil processes (e.g., nitrification, denitrification and N fixation) (Kowalchuk and Stephen, 2001; Malchair et al., 2010).
Historically, the coffee region in Colombia has undergone a transformation and fragmentation of natural ecosystems for the establishment of different crops such as bananas, onions, and citrus, as well as pastures for intensive livestock grazing (IGAC, 2007; Yanine, 2010). These farming practices have increased N inputs in soil through the application of organic and inorganic nitrogen fertilizer (e.g., manure, urea, NPK), generating significant environmental pollution and loss of soil fertility, through the alteration of processes such as nitrification and denitrification (Hurni et al., 2008). The first stage of nitrification (conversion of ammonium to nitrite) is carried out by ammoniumoxidizing archaea (AOA) and bacteria (AOB) (Avrahami and Bohannan, 2007). Subsequently, nitrite is oxidized to nitrate by nitrite-oxidizing bacteria (NOB), which can leach and contaminate ground and surface water (Chu et al., 2007; Junier et al., 2009; Kong et al., 2010). AOB can use nitrite as an electron acceptor in environments with a low availability of oxygen to generate NO, via reduction of NO2- and produced N2O (Bock and Wagner, 2006), another problem caused by nitrification is the acidification of the soils, which generates negative effects on the health and productivity of agricultural ecosystems.
Denitrification is a part of the global nitrogen cycle in which fixed nitrogen in the biosphere is returned to the atmosphere, and is mediated by diverse communities of microorganisms. This process is conducted by bacteria that use NO3- as an electron acceptor (Paul, 2007). Additionally, the production of N2O (by AOB or DB) is a major source of air pollution, contributing to the greenhouse effect (Bock and Wagner, 2006), which together with nitrification produces N losses, lower efficiency of fertilizers on crops, and affects the economy of farming (Pacheco et al., 2002).
Previous studies have shown that AOB and nitrogen fixing bacteria can be used as indicators of soil quality and health, due to their high sensitivity and low physiological tolerance to natural and anthropogenic disturbances (Nielsen and Winding, 2002; Horz et al., 2004; Nyberg et al., 2006; Roldán et al., 2008). Microorganisms possess the ability to give an integrated measure of soil quality, an aspect that cannot be obtained with physicochemical measures and analyses of diversity of higher organisms. Microorganisms respond quickly to changes so they rapidly adapt to environmental conditions. This adaptation potentially allows microbial analyses to be used in soil quality assessment, and changes in functional groups and activities may therefore function as an excellent indicator of change in soil quality (Nielsen and Winding, 2002).
Despite the biotechnological and environmental interests of N bacteria, few studies have been conducted in Colombia on the effect of different agricultural management practices and land uses in the density of functional groups associated with N cycling has been evaluated (Torres and Lizarazo, 2006; Roldán et al., 2008; Cardona et al., 2009). This can be related with the difficulties encountered during traditional laboratory culture techniques caused by factors such as slow growth, inadequate selection of the culture medium, ease of contamination by heterotrophic and possible activity loss after cultivation and preservation (Avrahami and Bohannan, 2007; Chu et al., 2007; Gómez, 2008).
The main goal of this study was to evaluate the effect of different land uses on the density of NB and DB. For this, it was necessary to standardize the optimal conditions for cultivation and enumeration of AOB, NOB and DB through the technique of most probable number (MPN). Finally, the isolated bacteria were taxonomically identified by amplification and sequencing of the 16S rRNA gene.
Materials and methods
Study area
The study area was located in the basins of the La Vieja and Otún rivers (Colombia Coffee Region), in the departments of Quindio, Valle del Cauca and Risaralda. These sites were selected based on criteria of spatial heterogeneity, altitudinal gradient and landscape structures by means of a canonical correspondence analysis of biotic and abiotic characteristics as well as the presence and representation of different land uses (coverage) of the region (Yanine, 2010). For each basin, the most representative land uses were selected: unshaded coffee (USCF), guadua and cattle pastures in the basin of La Vieja and onion monocultures, guadua and pine plantations (Pino patula) for Otún (Camargo, 2006). In each basin, systems with low human disturbance were selected (secondary forests) which were used as a reference site. For each land use, two different farms were selected and sampled (n=3).
Soil sampling
In each farm, three quadrants (2.5 x 2.5 m) were randomly established and samples were taken at the vertices and center. NB samples were taken using an auger (20 x 5 cm) which were mixed and homogenized to form a composite sample. For DB, a core sampler with cylinders (15 cm) was used to ensure low oxygen conditions. The samples were refrigerated (4-6°C) until processing in the laboratory. There were two sampling events (SE) in June and October, 2006.
Physicochemical analysis
We determined the soil in situ temperature, pH by the 9045C method (EPA, 1995), nitrate percentage by method No. 366 (HACH, 1994) and humidity (IGAC, 1994)
Microbiological analyzes
For optimal growth of AOB, NOB and DB, different carbon and energy sources reported in the literature were evaluated with the MPN technique (Stienstra et al., 1993; Aakra et al., 1999; Bigelow et al., 2002). NB were evaluated for the following carbon sources at different concentrations (g L-1): NaHCO3 (0.125) and CaCO3 (5.0 and 1.0). As an energy source (g L-1): (NH4)2SO4 (0.5 and 0.13) for AOB and NaNO2 (2.0 and 0.035) for NOB. For DB, KNO3 (2.0) was used as a terminal electron acceptor and as carbon sources: ethanol (1.0), glutamic acid (1.7) and sodium acetate (1.0) were evaluated.
Determining of NB density and isolation
We used the MPN technique in 96-well plates (Rowe et al., 1977) for determining the density of NB, using previously standardized mineral salts media. In each well, 100 µL of culture media for AOB and NOB and 50 mL of each dilution of the samples (10-2-10-7) were added with 5 replicates per dilution. The plates were incubated at 24±2°C for four weeks in darkness. AOB and NOB growth were verified by the disappearance of nitrite production, using diphenylamine and Griess-llosvay reagents, respectively (Rowe et al., 1977; Phillips et al., 2000). The combination of positive and negative wells was analyzed in the Most Probable Number program (version 4.04) and the results were expressed as MPN of NB/g dry weight (gdw). For the isolation of these bacteria, the dilution to extinction technique was used (Aakra et al., 1999).
Determining of DB density and isolation The MPN technique using the Hungate methodology was employed for DB (Pachón and Posada, 2003), with incubation for 15 d at 28±2°C. Growth and isolation was performed following the protocol described by Gómez (2008) and Pachón and Posada (2003).
Extraction of DNA from pure cultures of bacteria Two methods were used for extraction of DNA: the first by boiling at 95°C by suspending a colony in 50 µL of TE buffer (10 mM Tris-HCl pH 7.5, 1 mM EDTA, pH 8.0). The suspension was subjected to an initial heating (100°C for 15 min), followed by cooling on ice (5 min) and subsequent centrifugation at 13,000 rpm for 2 min. In the second method, lysozyme (10 µg mL-1) was used. The two methods followed the protocol described by Sambrook and Russell (2001).
PCR amplification of the gene 16S rRNA
To amplify the V3-V5 regions of the 16S ribosomal subunit, the universal bacterial primers 341F and 907R were used (Casamayor et al., 1999). All reactions were performed in a final volume of 25 µL containing 0.2 µM of each primer, 0.25 mM deoxynucleotide triphosphates mixture, 1X buffer, 2.5 mM MgCl2 and 0.025 U µL-1 of Taq polymerase (Taq Polymerase Kit, Promega) and 2 µL as DNA template. The program used consisted of 35 cycles with an initial denaturation phase at 94°C for 1 min, an annealing step at 54°C for 1 min and a final extension phase of 72°C for 10 min (Muyzer et al., 1993). The PCR product was verified by electrophoresis in agarose 1% (w/v) (120 V for 40 min) (Sambrook and Russell, 2001).
Sequence analysis and determination of the taxonomic affiliation
Sequences were analyzed using the BLAST program StandAlone version, using the Smith-Waterman algorithm (V. 2.2.9) from the National Center of Biological Information (NCBI) page and alignments were compared against the RDP database (Ribosomal Database Project) (v. 10).
Statistical analysis
The data that did not meet the requirements of normality and homogeneity of variances were transformed using logarithm base 10. To evaluate the effect of different land uses on the density of NB and DB, analysis of variance and Tukey-Kramer test (P=0.05), using the JMP statistical program (V.9.0), were used. To determine the relationship between soil physicochemical factors on the density of NB and DB, Pearson correlation analysis was performed.
Results and discussion
Physicochemical analysis
In general, soils of the study were characterized by a low pH (4.9-6.3) (Tables 1 and 2). This could be related mainly to the type of fertilizer used and the low buffering capacity of soil (Donahue et al., 1981). During the oxidation process of NH4+ by microorganisms, soil acidification occurs as a result of the continuous release of H+ (Kemmitt et al., 2006). Additionally, the metabolism of microorganisms and roots generates CO2 and soluble organic acids, decreasing the pH and behave as free acids of the soil (Inselsbacher et al., 2010). Our data showed a significant effect of land use on the pH, where forests had significantly lower pH values in the two river basins (P=0.05) (Tables 1 and 2). These land uses had higher organic matter content, which when is decomposed, release active carboxylic and phenolic groups, behaving as weak acids, acidifying the soil (Ritchie and Dolling, 1985).
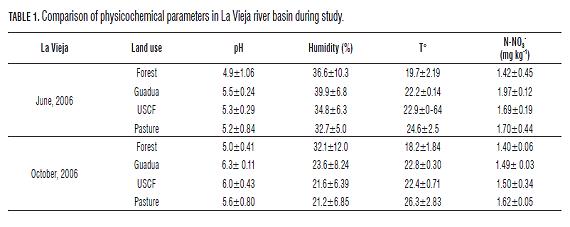
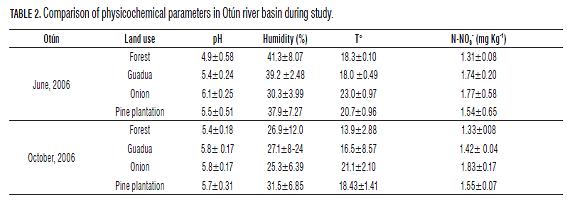
Forest and guadua values were significantly higher for humidity and lower for temperature (P =0.05) (Tabs. 1 and 2). This may be associated with the presence of a large tree cover in the forests which generates more leaf loss input, less solar radiation and decreased wind speed, factors that contribute to lower temperatures and evaporation processes (Sylvia et al., 2005). On the other hand, higher temperatures and lower humidity was observed in pasture and onion uses (Tables 1 and 2), possibly caused by the absence of tree cover and the use of conventional agricultural management practices (tillage and overgrazing), which cause soil compaction affecting water infiltration processes (Murgueitio, 2003; Yanine, 2010). Finally, in forests and some guadua sites, the observed lowest values of nitrates could be due to the slow processes of mineralization that could limit the growth of DB (Azam et al., 1995).
Optimal culture conditions for the growth of NB and DB For all the land uses evaluated, there were significantly higher densities of AOB and NOB using NaHCO3 (0.125 g L-1) as a carbon source (P=0.05) (data not shown). Additionally, the use of this compound facilitated the reading of the positive wells, reducing turbidity and minimizing evaporation of the culture medium. We selected the concentration of 0.5 g L-1 (NH4)2SO4 for AOB, which has been previously reported (Aakra et al., 1999; Bruns et al., 1999) and 0.035 g L-1 of NaNO2 was selected for NOB as an energy source (Stienstra et al., 1993). For DB a mixture of ethanol and sodium acetate as a carbon source were selected (Gómez, 2008).
NB and DB densities in the different land uses AOB densities for the first SE did not differ significantly between the different land uses (P= 0.05), although pastures and USCF had higher densities (Fig.1). For the second SE, there was a significant decrease in AOB counts in USCF (La Vieja) and forest (Otún) (P=0.05) (Fig.1). Additionally, onion and forest (Otún) had AOB densities significantly higher and lower, respectively (P=0.05), which may be associated with pH values observed in these land uses. It is well known that in agricultural soils, the density and activity of AOB decreases when the pH is below 5.5 or above 9.0, the optimum pH for nitrification is between 7.6-8.0 (Atlas and Bartha, 2002; Nugroho et al., 2007), therefore, significantly lower values of pH present in the forest and higher in onion (Table 2) may have affected AOB density.
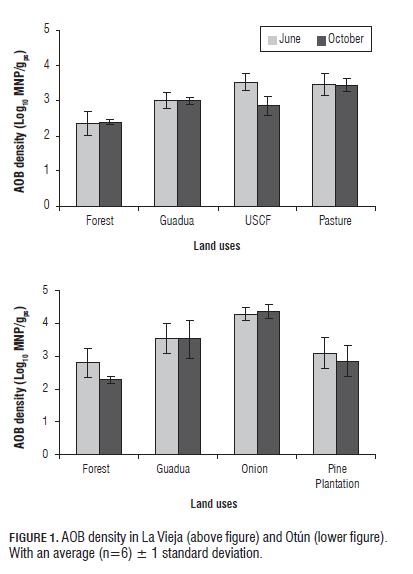
For all land uses assessed, AOB showed positive correlations (P=0.05) with pH, nitrates and temperature (R2 = 0.95, 0.85, 0.70, respectively) and negatively with moisture (R2 = -0.84). This could be related to increased moisture in the soil that could reduce the availability and transfer of oxygen, this being the final electron acceptor needed for the oxidation of ammonia.
NOB densities in La Vieja were significantly lower (P=0.05) for the second SE (Fig.2). Unlike AOB, NOB did not differ significantly between land uses for any SE, but the forest and onion counts were lower and higher, respectively, for Otún (P=0.05) (Fig.2). In general, higher counts of NB and mainly AOB in the land uses pasture, USCF and onion can be associated with implemented agricultural management practices, which involve the addition of nitrogen fertilizers (NPK-15: 15: 15, urea and organic fertilizers: cattle and poultry manure) that produce an increased availability of ammonium for NB (Hayden et al., 2010; Inselsbacher et al., 2010). The results are in agreement with previous studies which shown that agricultural practices, such as mineral N fertilizer application, significantly alter AOB populations, which subsequently impact nitrification and production rates of nitrous oxide (N2O), a potent greenhouse gas, in crop production systems (Kowalchuk and Stephen, 2001). Additionally, the grazing and tillage activities cause changes in physicochemical properties of soil (e.g., porosity, bulk density, penetration resistance, availability of oxygen and nutrients) affecting the density of NB (McNaughton et al., 1997; Busso et al., 2001; Patra et al., 2005; Siavosh et al., 2000; Murgueitio, 2003). On the other hand, pastures showed a positive correlation (P=0.05) between the density of AOB and NOB and nitrates (R2 = 0.942 and R2 = 0.907, respectively), which is the end product of nitrification, reflecting the active role of these two microbial groups in N cycling.
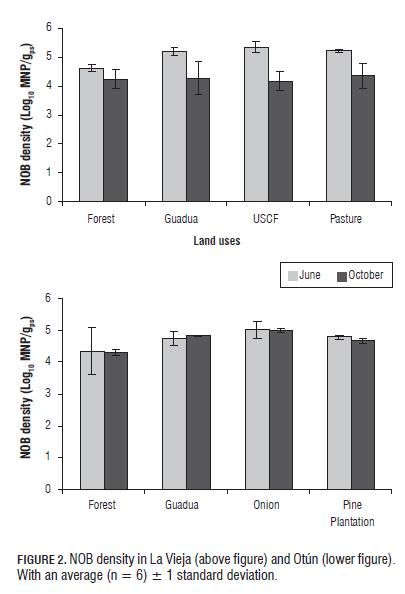
The low densities of NB in forests (Figs. 1 and 2) could be associated with low pH values, but also with low N availability and quality of organic matter. In forests, trees are the predominant vegetation, which produce woody plant residues with high C: N ratios and high concentrations of polyphenolic compounds (Kowalchuk and Stephen, 2001), which have proven to be toxic and inhibitory to AOB, mainly Nitrosomonas (Blanco, 2007; Kowalchuk and Stephen, 2001). The residues that have a high C:N ratio produce strong competition between NB and heterotrophs for available N in the soil, with heterotrophs leading the competition for available NH4+, decreasing its density (Verhagen et al., 1995).
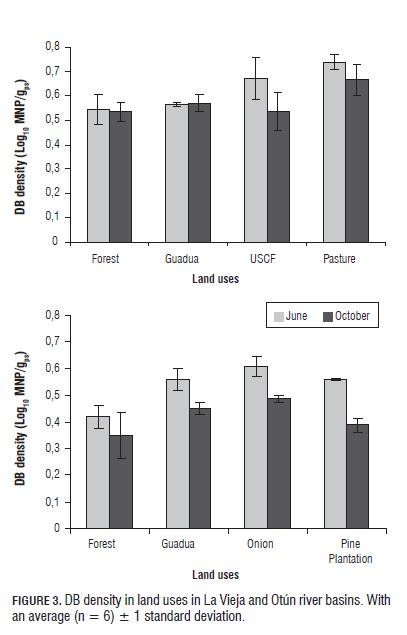
Finally, DB was significantly higher in USCF and pasture (La Vieja) for the first SE, and for the second SE was significantly higher only in pasture (P=0.05). Additionally, there was a significant decrease (P=0.05) in counts in the second SE in USCF and pasture (Fig.3). Furthermore, onion (Otún) presented the highest densities of DB for both SEs (Fig.3), with the forest and pine plantation land uses having significantly lower densities (P =0.05). DB are favored by the products generated during nitrification by NB, which also are stimulated by organic and inorganic fertilizers (Ellis and Pennington, 1989; Hill and Cardaci, 2004). Thus, in pasture and onion, positive correlations were found between the densities of AOB and DB (R2 = 0.90 and R2 = 0.92, respectively for pasture) and NB and DB (R2 = 0.84 and R2 = 0.98, respectively for onion). On the other hand, pastures showed a positive correlation between DB density and nitrates (R2 = 0.981, P=0.05), indicating that the high densities seen in this land use were favored by nitrates. These results agree with those reported by Kong et al. (2010), who showed a greater abundance of nosZ genes as a result of a higher input of organic C from fertilizations, which contain easily degradable C compounds, which stimulate denitrification. Conversely, the low DB densities in forest are possibly due to the low NB densities, which is consistent with that reported by Rich et al. (2003).
Taxonomic identification by the 16S ribosomal subunit of AOB, NOB and DB
Only 13 AOB were isolated due to the difficulty in obtaining pure cultures because the heterotrophic bacterial contamination. Of the strains isolated, 11 showed high similarity (98-99%) with the genus Hyphomicrobium. While this genus has been widely recognized as denitrifying, it has been demonstrated as being capable of using ammonium as an energy source (Brook et al., 1987). The 2 others showed similarity with the genera Mesorhizobium (95%) and Bradyrhizobium (98%) (Supplementary Table S1), which have not been reported as AOB, but as N-fixing (Teske et al., 1994; Compton et al., 2004).
For NOB, 7 isolates were found; 6 of them had high similarity (97-100%) with the species Oligotropha carboxidovorans and the remaining one presented similarity (100%) with Rhodopseudomonas palustris (Table supplementary S2), which are phylogenetically related with each other and to genera reported as nitrite oxidizers (Meyer, 2005; Aamand et al., 1996).
As for DB, 30 of the 39 isolates were identified, and 9 of them showed similarity (92-99%) with partial 16S rRNA sequences for non-cultivable bacteria. The genera found in La Vieja corresponded to Pseudomonas, Klebsiella, Citrobacter, and Microvirgula, while in Otún the genera found corresponded to Klebsiella, Bacillus, Enterobacter, Microvirgula, and Pantoea., These genera correspond to the ß-proteobacteria, ?-proteobacteria and Bacillales subdivisions (Supplementary Table S3), that have been previously reported by other studies (Cofman and Levine, 1986; Patureau et al., 2000; Rösch et al., 2002; Braker and Tiedje, 2003; Dandie et al., 2007; Morozkina and Zvyagilskaya, 2007). On the other hand, the genus Bacillus was only found in USCF and onions, and has been reported as one of the most representative microorganisms in altered soils (Dandie et al., 2007).
Conclusions
Land use had a significant effect on NB and DB densities. In addition, the higher densities of these microorganisms were observed in pastures, USCF and onion. Land use and land management practices (e.g., use of nitrogen fertilizers, tillage and grazing) had an effect on the densities of these bacteria associated with the N cycle. On the other hand, AOB densities were significantly different in the evaluated land uses, suggesting its use as a potential indicator of soil quality. On the other hand, forests had the lowest AOB densities, possibly due to reduced availability of N and release of allelopathic compounds. Moreover, although not taxonomically identified, NB bacterial genera and species were found involved in N cycling and closely related to the genera Nitrosomonas and Nitrosospira, which are reported as abundant in agricultural soils. As for DB, all sequences related to genera and species previously reported in agricultural soils were found.
Acknowledgement
Thanks to the Centro de Investigación en Biodiversidad y Recursos Genéticos (Research Center for Biodiversity and Genetic Resources) (Ciebreg)-COLCIENCIAS and the Vicerrectoría Académica de la Pontificia Universidad Javeriana for funding for this study.
Bibliography
Aakra, A., J. Utaker, I. Nes, and L. Bakken. 1999. An evaluated improvement of the extinction dilution method for isolation of ammonia-oxidizing bacteria. J. Microbiol. Meth. 39, 23-31. [ Links ]
Aamand, J., T. Ahl, and E. Spieck. 1996. Monoclonal antibodies recognizing nitrite oxidoreductase of Nitrobacter hamburgensis, N. winogradskyi, and N. vulgaris. Appl. Environ. Microbiol. 62(7), 2352-2355. [ Links ]
Allison, S.D. and J.B.H. Martiny. 2008. Resistance, resilience, and redundancy in microbial communities. Proc. Natl. Acad. Sci. USA 105, 11512-11519. [ Links ]
Atlas, R.M. and R. Bartha. 2002. Ecología microbiana y microbiología ambiental. Addison Wesley, Madrid. [ Links ]
Avrahami, S. and B. Bohannan. 2007. Response of Nitrosospira sp. strain AF-Like ammonia oxidizers to changes in temperature, soil moisture content, and fertilizer concentration. Appl. Environ. Microbiol. 73(4), 1166-1173. [ Links ]
Azam, F., R. Mulvaney, and E. Simmons. 1995. Effects of ammonium and nitrate on mineralization of nitrogen from leguminous residues. Biol. Fertil. Soils. 20, 49-52. [ Links ]
Bigelow, C., D. Bowman, and A. Wollum. 2002. Characterization of soil microbial population dynamics in newly constructed sand-based root zones. Crop Sci. 42(5), 1611-1614. [ Links ]
Blanco, J. 2007. The representation of allelopathy in ecosystem-level forest models. Ecol. Modell. 209, 65-77. [ Links ]
Bock, E., and Wager, M. 2006 Oxidation of inorganic nitrogen compounds as an energy source. In: Dworkin, M., Falkow, S., Rosemberg, E., Schleifer K-H., Stackebramdt, E. (Eds.) Prokaryotes, Vol 2, Springer, NY, pp 457-495. [ Links ]
Braker, G. and J. Tiedje. 2003. Nitric oxide reductase (norB) genes from pure cultures and environmental samples. Appl. Environ. Microbiol. 69(6), 3476-3483. [ Links ]
Brook, A., M. Duchars, and M. Attwood. 1987. Nitrogen assimilation in the facultative methylotroph Hyphomicrobium X. FEMS Microbiol. Lett. 41(1), 41-45. [ Links ]
Bruns, M., J. Stephen, G. Kowalchuk, L. Prosser, and P. Eldor. 1999. Comparative diversity of ammonia oxidizer 16S rRNA gene sequences in native, tilled, and successional soils. Appl. Environ. Microbiol. 65(7), 2992-3000. [ Links ]
Busso, C.A., D.D. Briske, and V. Olalde-Portugal. 2001. Root traits associated with nutrient exploitation following defoliation in three coexisting perennial grasses in a semi-arid savanna. Oikos 93, 332-342. [ Links ]
Camargo, J. 2006. Informe de la dirección científica del CIEBREG. Informe electrónico. Pereira, Colombia. [ Links ]
Cardona, G., C. Peña, and M. Ruíz-García. 2009. Comunidades de hongos actinomicetos en tres tipos de vegetación de la Amazonía colombiana: abundancia, morfotipos y el gen 16s ADNr. Rev. Biol. Trop. 57(4), 1119-1139. [ Links ]
Casamayor, E., H. Schafer, L. Bañeras, C. Pedros, and G. Muyzer. 1999. Identification of spatio-temporal differences between microbial assemblages from two neighboring sulfurose lakes: comparison by microscopy and denaturing gradient gel electrophoresis. Appl. Environ. Microbiol. 66(2), 499-508. [ Links ]
Cofman, I. and J. Levine. 1986. Relative rates of nitric oxide and nitrous oxide production by nitrifiers, denitrifiers, and nitrate respirers. Appl. Environ. Microbiol. 51(5), 938-945. [ Links ]
Compton, J., L. Watrud, A. Porteous, and S. DeGrood. 2004. Response of soil microbial biomass and community composition to chronic nitrogen additions at Harvard forest. For. Ecol. Mgt. 196, 143-158. [ Links ]
Chu, H., T. Fujii, S. Morimoto, X. Lin, K. Yagi, J. Hu, and J. Zhang. 2007. Community structure of ammonia-oxidizing bacteria under long-term application of mineral fertilizer and organic manure in a sandy loam soil. Appl. Environ. Microbiol. 73, 485-491. [ Links ]
Dandie, C., D. Burton, B. Zebarth, J. Trevors, and C. Goyer. 2007. Analysis of denitrification genes and comparison of nosZ, cnorB and 16S rDNA from culturable denitrifying bacteria in potato cropping systems. Syst. Appl. Microbiol. 30, 128-138. [ Links ]
Donahue, R.L., R.W. Miller, and J.C. Shickluna. 1981. Introducción a los suelos y al crecimiento de las plantas. Prentice Hall, Englewood Cliffs, NJ. [ Links ]
EPA, United States Environmental Protection Agency. 1995. Methods for evaluating solid wastes. Physical/chemical methods. Method 9045C. Soil and waste pH. SW-846. Chicago, IL. [ Links ]
Ellis, R. and P. Pennington. 1989. Nitrification in soils of secondary vegetational successions from Eucalyptus forest and grassland to cool temperate rainforest in Tasmania. Plant Soil 115(1), 59-73. [ Links ]
Feng, Y., A.C. Motta, D.W. Reeves, C.H. Burmester, E. Van Santen, and J.A. Osborne. 2003. Soil microbial communities under conventional-till and no-till continuous cotton systems. Soil Biol. Biochem. 35(12), 1693-1703. [ Links ]
Girvan, M., J. Bullimore, J. Pretty, M. Osborn, and A. Ball. 2003. Soil type is the primary determinant of the composition of the total and active bacterial communities in arable soils. Appl. Environ. Microbiol. 69(3), 1800-1809. [ Links ]
Gómez, M. 2008. Efecto de usos de suelo en la Ecorregión Cafetera sobre la densidad de bacterias nitrificantes y desnitrificantes. M. Sc. thesis. Pontificia Universidad Javeriana, Bogota. [ Links ]
HACH. 1994. SIW-1 Soil and irrigation manual. DREL/2000. Loveland, CO. [ Links ]
Hayden, H., J. Drake., M. Imhof, A. Oxley, S. Norng, and P. Mele. 2010. The abundance of nitrogen cycle genes amoA and nifH depends on land-uses and soil types in South-Eastern Australia. Soil. Biol. Biochem. 42, 1774-1783. [ Links ]
Hill, A. and M. Cardaci. 2004. Denitrification and organic carbon availability in riparian wetland soils and subsurface sediments. Soil Sci. Soc. Amer. J. 68, 320-325. [ Links ]
Horz, H.P., A. Barbrook, C.B. Field, and B.J.M. Bohannan. 2004. Ammonia-oxidizing bacteria respond to multifactorial global change. Proc. Natl. Acad. Sci. USA 101, 15136-15141. [ Links ]
Hunter, M. 1996. What is biodiversity? pp. 19-31. In: Malcolm L. Hunter and James P. Gibbs (eds.). Fundaments of conservation biology. 2nd ed. Blackwell Sciences, Oxford, UK. [ Links ]
Hurni, H., K. Herweg, B. Portner, and H. Liniger. 2008. Soil erosion and conservation in global agriculture. pp. 41-71. In: Braimoh, A. and P.L.G. Vlek (eds.). Land use and soil resources. Springer, Heidelberg, Germany. [ Links ]
IGAC, Instituto Geográfico Agustín Codazzi. 1994. Métodos analíticos del laboratorio de suelos. Ministerio de Hacienda y Crédito Público, Bogota. [ Links ]
IGAC, Instituto Geográfico Agustín Codazzi. 2007. Definición de usos alternativos y sostenibles para la ocupación de las tierras a nivel nacional (CD). Bogota. [ Links ]
Inselsbacher, E., N. Hinko-Najera Umana, F.C. Stange, M. Gorfer, E. Schüller, K. Ripka, S. Zechmeister-Boltenstern, R. Hood- Novotny, J. Strauss, and W. Wanek. 2010. Short-term competition between crop plants and soil microbes for inorganic N fertilizer. Soil Biol. Biochem. 42(2), 360-372. [ Links ]
Junier, P., M. Carú, and K.P. Witzel. 2009. Effect of common bean (Phaseolus vulgaris L.) on the community composition of ammonia-oxidizing bacteria in soil previously cultivated with Medicago sativa. Eur. J. Soil Biol. 45(3), 252-258. [ Links ]
Kemmitt, S., D. Wright, W. Keith, D. Goulding, and L. Jones. 2006. pH regulation of carbon and agricultural soil. Soil Biol. Biochem. 38(5), 1-14. [ Links ]
Kong, A., K. Hristova, K. Scow, and J. Six. 2010. Impacts of different N management regimes in nitrifier and denitrifier communities and N cycling in soil microenvironments. Soil. Biol. Biochem. 42(9), 1523-1533. [ Links ]
Kowalchuk, G.A. and J. Stephen. 2001. Ammonia-oxidizing bacteria: A model for molecular microbial ecology. Ann. Rev. Microbiol. 55(1), 485-529. [ Links ]
Mcnaughton, S.J., F.F. Banykwa, and M.M. Macnaughton. 1997. Promotion of the cycling of diet-enhancing nutrients by African grazers. Science 278, 1798-1800. [ Links ]
Malchair, S., H.J. De Boeck, C. Lemmens, R. Ceulemans, R. Merckx, I. Nijs, and M. Carnol. 2010. Diversity-function relationship of ammonia-oxidizing bacteria in soils among functional groups of grassland species under climate warming. Appl. Soil Ecol. 44, 15-23. [ Links ]
Matson, P.A., W. Parton, A. Power, and M. Swift. 1997. Agricultural intensification and ecosystem properties. Science 277, 504-508. [ Links ]
Meyer, O. 2005. Genus VII. Oligotropha Meyer, Stackebrandt, and Auling 1994, 182VP (Effective publication: Meyer, Stackebrandt and Auling 1993, 391). pp. 468-471. In: Brenner, D.J., N.R. Krieg, G.M. Garrity, J.T. Staley, D.R. Boone, P. De Vos, M. Goodfellow, F.A. Rainey, and K.-H. Schleifer (eds.). Bergey's Manual® of Systematic Bacteriology. Vol. 2 The Proteobacteria. Class I., Order VI. Springer, New York, NY. [ Links ]
Morozkina, E. and R. Zvyagilskaya. 2007. Nitrate reductases: structure, functions, and effect of stress factors. Biochemistry 72(10), 1151-1160. [ Links ] [ Links ]
Murgueitio, E. 2003. Impacto ambiental de la ganadería de leche en Colombia y alternativas de solución. Livest. Res. Rur. Dev. 5(10), 1-15. [ Links ]
Nielsen, M.N. and A. Winding. 2002. Microorganisms as indicators of soil health. Technical Report No. 388. National Environmental Research Institute, Denmark. [ Links ]
Nugroho, R., W. Roling, A. Laverman, and H. Verhoef. 2007. Low nitrification rates in acid scots pine forest soils are due to pHrelated factors. Microb. Ecol. 53(1), 89-97. [ Links ]
Nyberg, K., A. Schnürer, I. Sundh, Å. Jarvis, and S. Hallin. 2006. Ammonia-oxidizing communities in agricultural soil incubated with organic waste residues. Biol. Fert. Soils. 42, 315-323. [ Links ]
Pacheco, J., R. Pat, and A. Cabrera. 2002. Análisis del ciclo del nitrógeno en el medio ambiente con relación al agua subterránea y su efecto en los seres vivos. Ingeniería Revista Académica 6(3), 73-81. [ Links ]
Pachón, A. and Y. Posada. 2003. Cuantificación de poblaciones anaerobias aminoacidoliticas y aminoliticas. Undergraduate thesis. Pontificia Universidad Javeriana, Bogota. [ Links ]
Parfitt, R., G. Yeates, D. Ross, N. Schon, A. Mackay, and D. Wardle. 2010. Effect of fertilizer, herbicide and grazing management of pastures on plant and soil communities. Appl. Environ. Microb. 45, 175-186. [ Links ]
Patra, A.K., L. Abbadie, A. Clays-Josserand, V. Degrange, S.J. Grayston, P. Loiseau, F. Louault, S. Mahmood, S. Nazaret, L. Philippot, F. Poly, J.I. Prosser, A. Richaume, and X. Le Roux. 2005. Effects of grazing on microbial functional groups involved in soil N dynamics. Ecol. Monogr. 75, 65-80. [ Links ]
Patureau, E., J. Zumstein, P. Delgenes, and R. Moletta. 2000. Aerobic denitrifiers isolated from diverse natural and managed ecosystems. Microb. Ecol. 39, 145-152. [ Links ]
Paul, E. 2007. Soil microbiology, ecology and biochemistry. 3th ed. Academic Press, Burlington, VT. [ Links ]
Phillips, C., D. Harris, S. Dollhopf, K. Gross, J. Prosser, and E. Paul. 2000. Effects of agronomic treatments on structure and function of ammonia-oxidizing communities. Appl. Environ. Microbiol. 66(12), 5410-5418. [ Links ]
Rich, J., R. Heichen, P. Bottomley, K. Cromack, and D. Myrold. 2003. Community composition and functioning of denitrifying bacteria from adjacent meadow and forest soils. Appl. Environ. Microbiol. 69(10), 5974-5982. [ Links ]
Ritchie, G.S.P. and P.J. Dolling. 1985. The role of organic matter in soil acidification. Austrasl. J. Soil Res. 23, 569-576. [ Links ]
Roldán, F., A. Varela, E. García, L. Sierra, V. Vallejo, M. Berdugo, M. Aguilera, M. Santos, A. Cubillos, M.M. Gómez, M. Gómez-Sarmiento, N. LaTorre, and A. Vela. 2008. Evaluación del efecto de diferentes usos del suelo sobre grupos funcionales microbianos edáficos en la Ecoregión Cafetera Colombiana. pp. 238. In: Rodríguez, C., J. Niño, A. Pineda, L. Arias, M, Echeverri, C. Miranda (eds.). Valoración de la biodiversidad en la Ecoregión del eje Cafetero. CIEBREG, Pereira, Colombia. [ Links ]
Rösch, C., A. Mergel, and H. Bothe. 2002. Biodiversity of denitrifying and dinitrogen-fixing bacteria in an acid forest soil. Appl. Environ. Microb. 68(8), 3818-3829. [ Links ]
Rowe, R., R. Todd, and J. Waide. 1977. Microtechnique for most probable number analysis. Appl. Environ. Microb. 33, 675-680. [ Links ]
Sambrook, J. and D.W. Russell. 2000. Molecular cloning: a laboratory manual. 3th ed. Cold Spring Harbor Laboratory Press, New York, NY. [ Links ]
Sylvia, D.M., J.J. Fuhrmann, P.G. Hartel, and D.A. Zuberer. 2005. Principles and applications of soil microbiology. 2nd ed. Prentice Hall, Upper Saddle River, NJ. [ Links ]
Siavosh, S., J. Rivera, and M. Gómez. 2000. Impacto de sistemas de ganadería sobre las características físicas, químicas y biológicas de suelos en los Andes de Colombia. pp. 77-95. In: Agroforestería para la Producción Animal en Latinoamérica. FAO-CIPAV, Cali, Colombia. [ Links ]
Stienstra, A., G. Both, S. Gerards, and H. Laanbroek. 1993. Numbers of nitrite-oxidizing bacteria in the root zone of grassland plants. FEMS Microbiol. Ecol. 12, 207-214. [ Links ]
Teske, A., E. Alm, J. Regan, S. Toze, S. Rittman, and A. Stahl. 1994. Evolutionary relationships among ammonia and nitriteoxidizing bacteria. J. Bacteriol. 176(21), 6623-6630. [ Links ]
Torres, M. and L. Lizarazo. 2006. Evaluación de grupos funcionales (ciclo del C, N, P) y actividad de la fosfatasa acida en dos suelos agrícolas del departamento de Boyacá (Colombia). Agron. Colomb. 24, 317-325. [ Links ]
Verhagen, F., H. Laanbroek, and J. Woldendrop. 1995. Competition for ammonium between plant roots and nitrifying and heterotrophic bacteria and the effects of protozoan grazing. Plant Soil 170(2), 241-250. [ Links ]
Yanine, H. 2010. Evaluación de la diversidad de bacterias degradadotas de hidrocarburos aisladas de suelos de las cuencas de los ríos Otún y la Vieja. Tesis de Maestría en Microbiología. Universidad Nacional de Colombia. 122p. [ Links ]

