










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.29 n.3 Bogotá set./dez. 2011
SCIENTIFIC NOTES
In vitro morphogenic responses of Vasconcellea chilensis Planch. ex A. DC (Caricaceae)
Respuestas morfogenéticas in vitro de Vasconcellea chilensis Planch. ex A. DC (Caricaceae)
Miguel Jordan Z.1
1Institute of Biotechnology, Universidad Mayor. Santiago (Chile). miguel.jordan@umayor.cl Received for publication: 30 August, 2011. Accepted for publication: 2 November, 2011.ABSTRACT
Multiple shoot formation was induced in vitro on nodal sections excised from adult "palo gordo" plants of Vasconcellea chilensis ex Carica chilensis, in the presence of relatively high levels of thidiazuron (TDZ), indole-3- acetic acid (IAA) and organic addenda included in the WPM formulation including casein hydrolysate, adenine sulfate and cystein. Multiple shoots appeared simultaneously in strands or clusters on the surface of the explants; usually two or more clusters with new shoots were initiated, all showing synchronous growth. In subculture, in the presence of indole-3-butyric acid (IBA) as single hormone, root formation did not take place; the shoots turned green instead, evidenced elongation and development of new shoots at the base of the explants. Other explants, such as petioles and leaf sections exhibited callus initiation upon the vascular system after 4-5 weeks; however, this tissue turned brown later on and died.
Key words: papaya silvestre, palo gordo, monte gordo, endemic, endangered, in vitro, multiple shoots.
RESUMEN
La formación de brotes múltiples bajo condiciones in vitro pudo ser inducida en secciones nodales de plantas adultas de "palo gordo" Vasconcellea chilensis ex Carica chilensis, mediante el uso de altos niveles de tidiazuron (TDZ), ácido indolacético (AIA) y en presencia de addenda orgánica (hidrolizado enzimático de caseína, sulfato de adenina y cisteína) incluidas en el medio nutritivo "WPM". Dichos brotes se iniciaron sincrónicamente, en forma de agregados compactos y/o en hileras, sobre la superficie del explante. En subcultivo, los brotes evidenciaron elongación tornando a color verde, con iniciación de nuevos brotes desde la base y, a pesar de la presencia de ácido indolbutírico (AIB) como única hormona, no desarrollaron raíces. Otros explantes ensayados, tales como pecíolos, no evidenciaron respuesta. Solo secciones de hojas iniciaron la formación de una estructura compacta o callo a nivel de la nervadura principal transcurridas 4-5 semanas, tornándose pardo luego y con posterior muerte tisular.
Palabras clave: papaya silvestre, palo gordo, monte gordo, endémicas, en peligro de extinción, in vitro, brotes múltiples.
Introduction
The endemic to Chile Carica chilensis (Planch. ex A.B.C) Solms-Laub, at present Vasconcellea chilensis (Badillo, 2000, 2001) "palo gordo", (Fig.1a) has been classified as an endangered species (Benoit, 1989; Squeo et al., 2001). V. chilensis grows restricted to coastal semiarid rocky locations in low-density natural stands within South Latitude 30°-33°. In its habitat, the plant shows very slow growth, poor biomass production, small-sized fruit (Fig.1b) and germination problems. As it is not fit for human consumption its main use is as forage (Hechenleitner et al., 2005), but it is not cultivated as a forage crop due its various limitations, it is just gathered from the matorral and therefore has only little economic importance. As a result, there is scant information available regarding its growth conditions, phenology, and possible alternative uses of this species; i.e., search of valuable natural compounds in the latex (Kyndt et al., 2007). The germplasm potential and/ or specific traits of this species are yet to be researched. For example, the National Research Council (1989) indicated that several traits of a series of Vasconcellea species, although divergent between genera (Ocampo et al., 2006), are important due to their possible use in papaya breeding programs, especially to introduce cold adaptability and use of pathogen resistance genes present in Vasconcellea sp. highland papayas. The scant reports available for V. chilensis are mainly taxonomical studies; no information or studies on selection, morphogenic responses, grafting, nor asexual multiplication by cuttings or sexual multiplication by seeds have been reported (Hechenleitner et al., 2005, Jordan et al., 2009) up to now as far as information is available. The aim of this work was to explore the morphogenic potential existing in some V. chilensis explant-types as an alternative to achieve vegetative propagation of this endangered species.

Material and methods
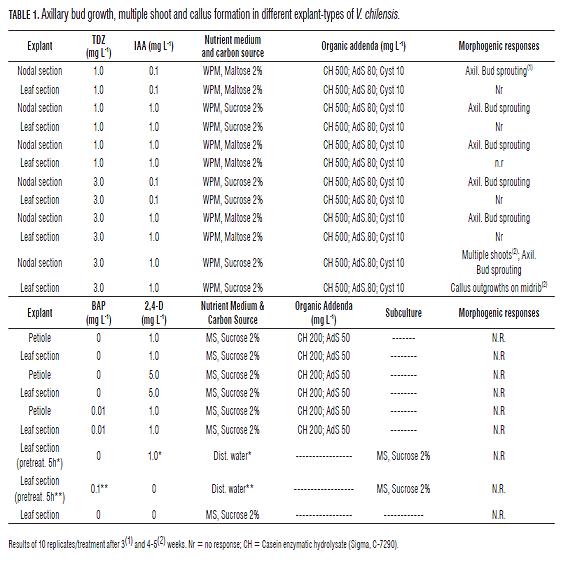
In a preliminary work (results not shown here) V. chilensis showed similar developing patterns as Vasconcellea x heilbornii cv. 'babaco' and similar plant growth regulator and/or organic addenda demands for establishment, shoot formation (Cohen and Cooper, 1982; Jordan and Piwanski, 1997) as well as for somatic embryogenesis (Jordan and Piwanski, 1999). Consequently, casein enzymatic hydrolysate, adenine sulphate and L-cysteine, were included in the media, and tested at levels of 500 mg L-1; 80 mg L-1 and 10 mg L-1, respectively; besides other concentrations (Tab.1). Among plant growth regulators, thidiazuron (TDZ), 6-benzylaminopurine (BAP), dichlorophenoxyacetic acid (2,4-D) and indole -3-acetic acid (IAA), alone or in combinations were tested. As maltose was shown to be critical for shoot initiation in babaco (Jordan and Piwanski, 1997); it was also tested besides sucrose at the level of 2% added to the WPM medium (Lloyd and McCown, 1980) as well as to the MS-medium (Murashige and Skoog, 1962). After surface disinfection for 2 h in a fungicide suspension of 1.0 g L-1 triadimefon and 5.0 g L-1 captan (80% WP), under agitation, the explants were immersed for 5 min in a 0.25% (v/v) sodium hypochlorite solution and finally rinsed twice in sterile distilled water. The explants, node and petiole sections (excised 1 cm long), as well as 1 cm2 leaf sections including the midrib, were transferred to Pyrex tubes containing 13 mL solidified media (0.4% Midesa® Agar). The nodal explants were sectioned from the uppermost region of the branches (nodes from the apex to 10 cm below). Leaf sections were cultured with their abaxial side in contact with the media (Jordan and Velozo, 1996). Explants (in culture or subculture) were maintained for 2 months under an 18-h photoperiod at mol m-2s-1 provided by fluorescent (General Electric F40T10-SS) light sources. Plant growth regulators, nutrients, vitamins and organic sources were all provided by Sigma and were added to media previous sterilization at 121°C during 15 min.
Results and discussion
Morphogenic responses concerning initiation of multiple shoots and/or somatic embryogenesis have been reported earlier in Vasconcellea x heilbornii cv. 'babaco' in cultivated node leaf explants, either from leaf tissue (Cohen and Cooper, 1982) or from preexisting vascular outgrowths connected to the leaf midrib (Vega de Rojas and Kitto, 1991; Vega de Rojas et al., 1993; Jordan and Piwanski, 1997, 1999). Similarly, V. chilensis explants (this report) showed several morphogenic responses in culture, but of a much lesser extent and restricted, regarding hormonal conditions in comparable hormonal conditions. In the case of node segments, the most noticeable first response was the initiation of multiple shoots. Multiple-shoot formation started with a very noticeable swelling of the node segment; this was more evident in treatments with the higher TDZ and IAA levels, and was completed after 10 d (Figs. 1c and d).
Next, a sprouting of the preformed axillary bud of the node segment, bud branching and proliferation of new shoots took place and was completed after 3 weeks. (Fig.1g, arrow). Finally and after a culture period of 4-5 weeks, a profuse initiation of multiple shoots all over the node surface was observed (Figs. 1c, d and e). Although axillary bud sprouting occurred in several TDZ/IAA combinations, multiple shoot formation from the node segment was only observed in the presence of the highest TDZ/IAA levels tested (3.0 and 1.0 mg L-1, respectively). (Tab.1). However, in the case of V. chilensis and diverging from babaco requirements, multiple shoot formation occurred in the presence of sucrose and not necessarily maltose. The new shoots appeared in clusters or rows on different portions of the node segments. Their color becomes visible initially white or yellowish, similar to the basal tissue; with some browning at the top (Fig 1c). Afterward, the shoots enlarged and turned green, developing synchronously (Fig.1e). Although at early stages the new shoots had a tightly closed shape, not unlike somatic embryos, closer examination by histological studies did not evidence a bipolar structure nor signs of root primordia; the shoots appeared as attached to the basal mother tissue (not in figures). Consequently, single shoot-tips, whole shoot clusters as well as axillary buds with a proliferation of new shoots were subcultured in conditions to promote rhizogenesis; that is, conditions similar to those that promoted root formation from de novo shoot-tips of babaco (Jordan and Piwanski, 1997). Results showed however, that root formation was not achieved in the presence of 2.0 mg L-1 IBA, used in various combinations alone or with organic addenda (not in Table); instead, the sub-cultured shoots stopped growing, turned brown and died. Similarly, attempts to induce rooting using other growth regulator combinations, alike to those used for babaco (Jordan and Piwanski, 1997) adding 100 mg L-1 phloroglucinol, as a root promoter (James and Thurbon, 1981), were not successful.
Scarce results were similarly observed for leaf explants. After a 4-week culture period, several pale outgrowths, looking like compact callus and/or nodules, size 2 mm developed upon the midrib vein in the presence of TDZ and IAA (3.0 and 1.0 mg L-1, respectively). TDZ has been reported as a potent growth regulator that induces shoot formation in many species (Huetteman and Preece, 1993); however contrary to babaco and in spite of the relatively high concentration employed, no further development of these outgrowths could be obtained. As indicated, under a similar concentration multiple shoot initiation took place on the pre-existing outgrowths attached to the vascular system of leaf sections in babaco (Jordan and Piwanski, 1997). These outgrowths, exhibited by babaco, located at the proximal region of the leaf explants, initiated new shoots also in higher TDZ concentrations (up to 7 mg L-1) and in other hormonal combinations, permitting plant regeneration by means root induction of shoot in subculture (Jordan and Piwanski, 1997). Finally, the petiole explants of V. chilensis did not evidence any morphogenic response and turned brown in the different treatments assayed. As a conclusion, although it was possible to induce some morphogenic responses in V. chilensis more work, regarding plant metabolism, as well as its control by specific growth regulators, must be invested to reach a better definition of the requirements and overcome the constraints limiting the regeneration of this species, as well as work to pinpoint the natural compounds present in the latex, besides papain.
Acknowledgment
The author wants to thank Mrs. Andrée Goreux for revision of the manuscript and Mr. Carlos Roveraro for technical assistance.
Literature cited
Badillo, V.M. 2000. Carica L. vs. Vasconcella St. Hil. (Caricaceae) con la rehabilitación de este último. Ernstia 10, 74-79. [ Links ]
Badillo, V.M. 2001. Nota correctiva Vasconcellea St. Hil. y no Vasconcella (Caricaceae). Ernstia 11, 75-76. [ Links ]
Benoit, I.L. 1989. Red book of chilean terrestrial fauna (Part One). CONAF, Santiago. [ Links ]
Cohen, D. and P.A. Cooper. 1982. Micropropagation of babaco. A Carica hybrid from Ecuador. pp. 743-744. In: Fujirawa A. (ed.) Proc. 5th International Cong. Plant Tissue and Cell Culture. Maruzen, Tokyo. [ Links ]
Hechenleitner, P., M. Gardner, P. Thomas, C. Echeverria, B. Escobar, P. Brownless, and C. Martínez. 2005. Plantas amenazadas del centro-sur de Chile. Universidad Austral de Chile; Real Jardín Botánico de Edimburgo, Santiago. [ Links ]
Huetteman, C. and J. Preece. 1993. Thidiazuron, a potent cytokinin for woody plant tissue culture. Plant Cell Tissue & Organ. Cult. 33, 105-119. [ Links ]
James, D.J. and I. Thurbon. 1981. Shoot and root initiation in vitro in apple rootstock M.9 and the promotive effects of phloroglucinol. J. Hort. Sci. 56, 15-20. [ Links ]
Jordan, M., I. Cortés, and G. Montenegro. 1983. Regeneration of plantlets by embryogenesis from callus cultures of Carica candamarcensis. Plant Sci. Lett. 28, 321-326. [ Links ]
Jordan, M. and J. Velozo. 1996. Improvement of somatic embryogenesis in highland-papaya cell suspensions. Plant Cell Tissue & Organ. Cult. 44,189-194. [ Links ]
Jordan, M. and D. Piwanski. 1997. Regeneration of babaco [Carica pentagona (Heilborn) Badillo] using leaf explants and shoot-tip culture. Fyton 61,109-115. [ Links ]
Jordan, M. and D. Piwanski. 1999. Regeneration of babaco (Carica pentagona) through somatic embryogenesis. Fyton 64, 101- 105. [ Links ]
Jordan, M., D. Vélez, and R. Armijos. 2009. Biotecnologías aplicables al desarrollo de algunas especies de Caricáceas cultivadas en la región Andina: avances y problemas. Rev. Colomb. Cienc. Hortíc. 3, 9-17. [ Links ]
Kyndt, T., E.J.M. Van Damme, J. Van Beeumen, and G. Gheysen. 2007. Purification and characterization of the cysteine proteinases in the latex of Vasconcellea spp. FEBS J. 274, 451-462. [ Links ]
Lloyd, G. and B. McCown. 1980. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia by use shoot-tip culture. Int. Plant Prop. Soc. Proc. 30, 421-427. [ Links ]
Murashige, T. and F. Skoog. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15, 473-497. [ Links ]
National Research Council. 1989. Lost crops of the Incas. Littleknown plants of the Andes with promise for worldwide cultivation. National Academy Press, Washington DC. [ Links ]
Ocampo, J.A., D. Dambier, P. Ollitrault, G. Coppens d'Eeckenbrugge, P. Brotier, and A.M. Risterucci. 2006. Microsatellite DNA in Carica papaya L.: isolation, characterization and transferability to Vasconcellea species. Mol. Ecol. Notes 6, 212-217. [ Links ]
Squeo, F.A., G. Arancio, and J.R. Gutiérrez. 2001. Libro rojo de la flora nativa y de los sitios prioritarios para su conservación: región de Coquimbo. Ediciones Universidad de La Serena, La Serena, Chile. [ Links ]
Van Droogenbroeck, B., P. Breyne, P. Goetghebeur, E. Romeijn- Peeters, T. Kyndt, and G. Gheysen. 2002. AFLP analysis of genetic relationships among papaya and its wild relatives (Caricaceae) from Ecuador. Theor. Appl. Genet. 105, 289-297. [ Links ]
Vega de Rojas, R. and S.L. Kitto. 1991. Regeneration of babaco (Carica pentagona) from ovular callus. J. Amer. Soc. Hort. Sci. 116, 747-752. [ Links ]
Vega de Rojas, R., C. Barberan, and S.L. Kitto. 1993. Shoot regeneration from peduncles and shoot-like regeneration from leaves of babaco, Carica pentagona. Turrialba 43, 93-99. [ Links ]

