










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. v.29 n.3 Bogotá set./dez. 2011
SCIENTIFIC NOTES
Diaphorina citri (Kuwayama, 1907) and Tamarixia radiata (Waterson, 1922) in citrus crops of Cundinamarca, Colombia
Diaphorina citri (Kuwayama, 1907) y Tamarixia radiata (Waterson, 1922) en cítricos en el departamento de Cundinamarca, Colombia
Everth Emilio Ebratt-Ravelo1,6, Leidy Tatiana Rubio-González 2, Valmir Antonio Costa3, Ángela Patricia Castro-Ávila4,Edna Milena Zambrano-Gómez 5,and Jorge Evelio Ángel-Díaz1
1Laboratorio Nacional de Diagnóstico Fitosanitario, Tibaitatá Research Center, Instituto Colombiano Agropecuario (ICA). Mosquera (Colombia).2Universidad Francisco de Paula Santander; Laboratorio Nacional de Diagnóstico Fitosanitario, Tibaitatá Research Center, Instituto Colombiano Agropecuario (ICA). Mosquera (Colombia).
3Centro Experimental do Instituto Biológico Rodovia Heitor Penteado. Campinas, SP (Brasil).
4Faculty of Agronomy, Universidad Nacional de Colombia. Bogota (Colombia)
5Fruit Project, Laboratorio Nacional de Diagnóstico Fitosanitario, Tibaitatá Research Center, Instituto Colombiano Agropecuario (ICA). Mosquera (Colombia).
6Corresponding author. everth.ebratt.ravelo@gmail.com Received for publication: 8 February, 2011. Accepted for publication: 2 November, 2011.
ABSTRACT
The presence of Diaphorina citri Kuwayama (Hemiptera: Psyllidae) and Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae) was recorded in citrus crops of Cundinamarca, Colombia. This work is to know the geographic distribution of D. citri and an initial record of the parasitoid T. radiata in citrus producing areas of this department.
Key words: biological control, Psyllidae, Eulophidae, citrus, Murraya sp.
RESUMEN
La presencia de Diaphorina citri Kuwayama (Hemiptera: Psyllidae) y Tamarixia radiata (Waterson) (Hymenoptera: Eulophidae) se registró en cultivos de cítricos en el departamento de Cundinamarca, Colombia. Igualmente se da a conocer la distribución geográfica de D. citri y se realiza un primer reporte del parasitoide T. radiata en regiones productoras de cítricos en este mismo departamento.
Palabras clave: control biológico, Psyllidae, Eulophidae, cítricos, Murraya sp.
Introduction
Diaphorina citri Kuwayama was reported in Colombia associated with citrus seedlings in various nurseries in the departments of Valle del Cauca and Tolima in 2007 by the Colombian Agricultural Institute (ICA). Afterwards in 2008 it was found in the town of Girardot in Cundinamarca by the same entity. In Colombia, there are approximately 60,000 ha planted in citrus of which 10,633 are in Cundinamarca (Plan Nacional Hortifrutícola (MADR, 2006).
According to Halbert and Núñez (2004), D. citri has 59 species of the family Rutaceae as hosts, especially of the genera Citrus sp. and Murraya sp. The importance of D. citri is its ability to transmit the disease known as Huanglongbing or HLB (Da Graca, 1991; Van der Merwe and Andersen, 1937; Broadbent et al., 1977; Aubert, 1987; Liu and Tsai, 2000; Tsai and Liu, 2000; Tsai et al., 2002), whose etiologic agent are the bacteria Candidatus Liberibacter asiaticus and Candidatus Liberibacter americanus in America (Halbert and Núñez, 2004).
This work aimed to acquire knowledge of the life cycle and the geographic distribution of Asian psyllid D. citri and reported the parasitoide Tamarixia radiata (Waterston) in citrus-producing regions in the department of Cundinamarca in Colombia.
Materials and methods
The monitoring began in November 2008 and ended in November 2010 and included the following regions: Gualiva, Alto Magdalena, Centro Magdalena, Bajo Magdalena, Rionegro, Sumapaz, Medina, Tequendama, and the east in the department of Cundinamarca; a classification method based on zones proposed by L.R. Holdridge (1967, 1982) and the IGAC (1997) were implemented for this purpose. Seven routes were established for the collection of samples, corresponding to the most important departmental roads: Fusagasugá-Bogota-Girardot, Bogota-Girardot-La Mesa, Bogota-San Francisco-Villeta-Guaduas, Bogota-Facatativá- Sasaima-Villeta, Bogota Zipaquirá-Pacho-La Palma, Bogota-Choachi-Caqueza-Bogota and Bogota-Viani-San Juan de Rioseco.
To determine the presence of D. citri and its natural enemies in crops, some samples were taken from valencia orange (Citrus sinensis Osbeck), mineola tangelo (Citrus reticulata x C. grandis), tangerine (Citrus reticulata Blanco), tahiti lime (Citrus aurantifolia Swingle), myrtle plantas (Murraya paniculata), and swinglia (Swinglea glutinosa), the last, used as living fences in citrus producing farms, cattle ranchs and residential condominiums.
D. citri adults were captured, the host plant was first surrounded with entomological jama, moving up and down, smoothly touching the plant leaves; the content was completely deposited in a sealable plastic bag, then it was observed and verified to assure the presence of adults inside and then were passed to vials with 70% of ethanol using a 00 caliber moistened brush. To capture immature was necessary a 20x loupe to identify infested shoots with either eggs or mass colonies of nymphs, were introduced in paper bags and then into sealable plastic bags. In each site ten plants were sampled for a total of 770 plants during the whole study. The samples were labeled with the basic data of the capture (town, village, farm, plant material, harvest date, geographic location) and to send to the National Laboratory of Plant Pathology Diagnostic– ICA Tibaitatá, located in Mosquera (Cundinamarca, Colombia, 4°41'46,57" N, 74°12'13" W and 2,544 m a.s.l.), where they were maintained at an average temperature of 24°C until the adult emergence of D. citri or its parasitoides from D. citri nymphs.
With the insect material of D. citri obtained in confinement, 10 seedlings of mandarin were infested, with a pair of D. citri per plant and kept in cages of 3 x 3 x 3 m with a swiss veil on top, at an average temperature of 22.27 ± 2.35°C. From the appearance of the first mass of eggs in each plant, the adults were removed; two daily readings were done (08:00 h and 16:00 h) according to the methodology proposed by Liu and Tsai (2000), Tsai and Liu, (2000), Tsai et al. (2002), Gómez and Postali (2009), for a period of 60 d. This also allowed determining the developmental stages of D. citri, their morphological, morphometrica, and the time between the different instants of development. In order to identify and characterize the states of development of D. citri and to obtain parasitoids emerged from pupae, the proposed methodology was the one implemented by Gómez y Postali (2009). The entomological material for adults of D. citri and T. radiata, were confirmed to species by Dr. Daniel Burckhardt (Entomologie Naturhistorisches Museum Basel Switzerland) and Dr. Valmir A. Costa (Centro Experimental do Instituto Biológico Campinas, Brasil), respectively.
Results and discussion
Geographical distribution and hosts
The presence of D. citri Kuwayama was detected in 16 of 20 local town producers of citrus crops established in orange, tangelo, tangerine, lime Tahiti, swinglia myrtle plants in altitude and temperature ranges average between 250 to 1,900 m and 35°C to 18°C respectively, with bimodal rainfall (650 to 3000 mm per year) in the inter-Andean region of the Magdalena River and unimodal in the eastern region (700 to 3,000 mm year-1).
In the department of Cundinamarca, D. citri is present in 60% of the citrus-producing regions and involved three life zones for tropical dry forest (bs-T), premontane dry forest (bs-PM) and premontane wet forest (bh-PM). It is not present in the region of Sasaima which is a very humid forest (bmh-PM), eventhough this zone is characterized for having the presence of host plants with the exception of myrtle, however, it is important to note that the conditions of high rainfall and low temperature, which occurred during this study could be generated adverse environmental conditions to D. citri, in contrast to those reported in other regions. Additionally, it is not present in the municipalities of Fómeque, Choachí, and the premontane wet forest (bh-PM), at the department eastern slope, possibly due to the temperature which is below 20°C and the low or no presence of host plants (Tab.1).
From 770 plants sampled, 525 were infested with D. citri. The largest occurrence corresponded to the myrtle (M. paniculata) with 100% findings (190/190 plants), followed by mandarin (C. reticulata) with 85,71% findings (120/140 plants), orange (C. sinensis) with 60% findings (120/200 plants), tangelo (C. reticulata x C. grandis) with 55,55% findings (50/90 plants), Tahiti lime (C. aurantifolia) with 50% findings (35/70 plants), and swinglia (S. glutinosa) with 50% findings (40/80 plants), confirming the results obtained by Liu and Tsai, (2000); Tsai and Liu, (2000), and Tsai et al. (2002); however, but under field conditions, the preference may be favored by the presence of hosts, the range of feeding sites and oviposition sites for the psyllid, by way of a greater number of shoots; above as a result of specific conditions like water stress, rainfall, and temperature en each zone involved. These conditions make allow to understand how difficult it is to provide regional management of D. citri, if the presence, distribution and host plant preference within the management plans of their populations is not taken into account
Life cycle of D. citri
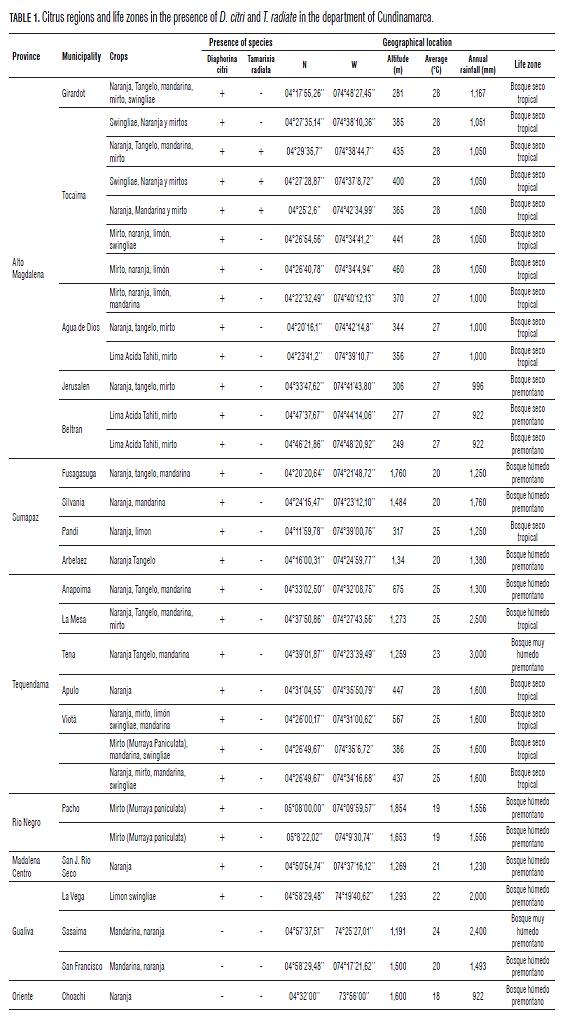
The insect showed the following life cycle stages: it was first in its egg, then five nymph stages and a final adult stage which took place over a period of 26.88±8.23 d to 22.27±2.35°C (Tmax= 27.37°C and Tmin= 18.04°C) ( Fig.1, Fig.2), with are differential characteristics in morphology and morphometry, according to Gómez and Postali (2009), that enable the characterization process of each of these stages of development (Tab.2, Tab.3), as observed by Liu and Tsai (2000), Tsai and Liu (2000), and Tsai et al. (2002), who determined the life cycle of D. citri is full filled in 28.6 ± 4.5 d at optimum temperatures of 25 and 28°C, and a significant reduction in oviposition, at temperatures outside these ranges.
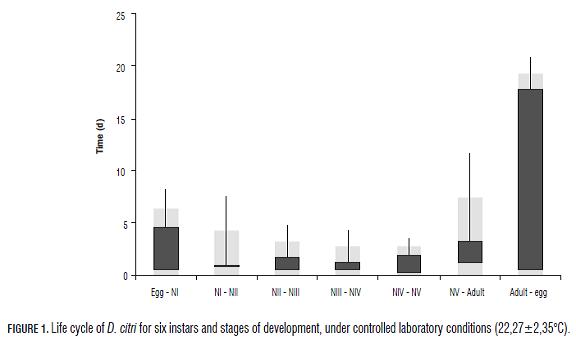

Worldwide, Tamarixia radiata (Waterston) is among the most potentially beneficial organisms for biological control of D. citri. T. radiata (Waterston) is a microhimenophtero of 1.43 mm (n=4; Ds= 0.3513) and 1.20 mm (n=4; Ds= 0.2008) in length in females and males respectively, which acts as a ectoparasitoide idiobionte in nymph III, nymph IV and nymph V of D. citri (Gómez and Postali, 2009; Chien et al., 2001). According to Chien et al. (2001), the female lays egg on the ventral part of the nymph III, between the thorax and the abdomen, the newly hatched larva sucks hemolymph from the outer zone which is attached to host integument, then the parasitoid is dragged to the ventral region of the chest and still feeding. When T. radiata fulfills all its development and eats the whole hemolymph of its host, the latter gets mummified and inside the mature parasitoid larva transforms into a prepupa and finnaly into a pupa. According to Gómez and Postali (2009), the life cycle of T. radiata from egg to adult emergence is accomplished in 12 d at 25°C or in 8 d at 30°C, and has longevity of 60 d at 20°C (Quilici and Fauvergue, 1990; Fauvergue and Quilici, 1991).
The nymphs of D. citri parasitized by T. radiata ( Fig.3), were obtained from vegetable material of mandarin and orange from Tocaima (04°29'35,7'' N, 74°38'44,7'' W y 435 msnm; 04°27'28,87'' N, 74°37'8,72'' W y 400 msnm; 04°25'2,6'' N, 74°42'34,99'' W y 365 msnm), located in the zone of tropical dry forest (bs-T), characterized by temperatures of 28°C and 1,050 mm mean annual precipitation, and a 15% of natural level of parasitoidism. These thermal-hygrometric conditions correspond to the optimal considered for T. radiata reported by Gómez and Postali (2009) in the state of Sao Paulo (Brazil), suggesting the dependence of the parasitoid to these environmental conditions for dispersion, colonization, and establishment in areas with D. citri presence.
Also, Aubert (1987) reported these conditions and he found levels of 80% parasitoidism in D. citri by inoculation of parasitoid of citrus crops in India, the Arabian Peninsula, Philippines, Thailand and Indonesia from entomological material that grown massively. According to Chien et al. (1991a, 1991b, 1995), the mass breeding of T. radiate permits inundative introductions of crops infested by D.citri, with more effective parasitoidism levels to those exercised by the natural parasitoidism.
Conclusions
D. citri is present in 60% of the citrus-producing areas of the Andean region in the department of Cundinamarca and has become the main phytosanitary problem of the citrus production in the department, being the main transmitter of the disease known as HLB.
The presence of T. radiata was reported as a natural enemy of D. citri colonizing citrus producing regions located in andean areas of tropical dry forest of the department of Cundinamarca and its presence is an alternative biological control management for the psyllid vector of HLB to the citriculture in Colombia.
Acknowledgments
To Instituto Colombiano Agropecuario – ICA, Cundinamarca seccional, local offices Girardot, Pacho, Caqueza, Fusagasuga, La Meza, Dr. Jorge Evelio Angel Díaz National Laboratory of Plant Pathology Diagnostic, Dr. Daniel Burckhardt for diagnostic confirmation the asian psyllid, and Dr. Valmir A. Costa (Centro Experimental do Instituto Biológico, Campinas Brasil) to confirm the T. radiata diagnostic.
Literature cited
Aubert, B. 1987. Trioza erytreae Del Guercio and Diaphorina citri Kuwayama (Homoptera: Psylloidae), the two vectors of citrus greening disease: Biological aspects and possible control strategies. Fruits 42, 149-162. [ Links ]
Broadbent, P., L.R. Fraser, A. Beattie, N. Grylls, and J. Duncan. 1977. Australian citrus dieback problem. Proc. Int. Soc. Citricult. 3, 894-896. [ Links ]
Chien, C.C., C. Yi and S.C. Ku. 1991a. Biological control of citrus psyllid, Diaphorina citri, in Taiwan. II. evaluation of Tamarixia radiata and Diaphorencyrtus diaphorinae for the control of Diaphorina citri. Chin. J. Entomol. 11, 25-38. [ Links ]
Chien, C.C., C. YI, and S.C. Ku. 1991b. Parasite strategy, morphology and life history of Tamarixia radiata (Hymenoptera, Eulophidae). Chin. J. Entomol. 11, 264-281. [ Links ]
Chien, C.C. 1995. The role of parasitoids in the pest management of citrus psyllid. pp. 245-261. In: Proceedings of the Symposium on Research and Development of Citrus in Taiwan, Taichung, Taiwan. [ Links ]
Chien, C.C., S. Chiu, and S.C. Ku. 2001. Mass rearing and field release of an eulophid wasp, Tamarixia radiata (Waterson). In: FFTC, Technical Notes No. 2001-5, http://www.agnet.org/ library/tn/2001005/; consulted: August, 2011. [ Links ]
Da Graca, J.V. 1991. Citrus greening disease. Annu. Rev. Phytopathol. 29, 109-136. [ Links ]
Fauvergue, X. and S. Quilici. 1991. Etude de certains parametres de la biologie de Tamarixia radiata (Waterston, 1922) (Hymenoptera: Eulophidae), ectoparasitoide primaire de Diaphorina citri Kuwayama (Hemiptera: Psyllidae), vecteur asiatique du greening des agrumes. Fruits 46(2), 179-185. [ Links ]
Gómez, M. and J.R. Postali. 2009. Estudos bioecologicos de Tamarixia radiata (Waterston, 1922) (Hymenoptera: Eulophidae) para o controle de D. citri Kuwayama, 1907 (Hemiptera: Psyllidae). Ph.D. thesis. ESALQ, Piracicaba, Brasil. [ Links ]
Halbert, S.E. and C.A. Núñez. 2004. Distribution of the Asian citrus psyllid, Diaphorina citri Kuwayama (Sternorrhyncha: Psyllidae) in the Caribbean basin. Fla. Entomol. 87, 401-402. [ Links ]
Holdridge, L.R. 1967. Life zone ecology. Tropical Science Center, San José. [ Links ]
Holdridge, L.R. 1982. Ecología basada en zonas de vida. IICA, San José. [ Links ]
IGAC, Instituto Geográfico Agustín Codazzi. 1997. Bases conceptuales y guía metodológica para la formulación del plan de ordenamiento territorial departamental. IGAC, Ministerio de Hacienda y Crédito Público, Bogota. [ Links ]
Liu, Y.H. and J.H. Tsai. 2000. Effects of temperature on biology and life table parameters of the Asian citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae). Ann. Appl. Biol. 137, 210-216. [ Links ]
MADR, Ministerio de Agricultura y Desarrollo Rural. 2006. Documento técnico. Plan Nacional Hortofruticola (PNH). Bogota. [ Links ]
Quilici, S. and X. Fauvergue. 1990. Studies on the biology of Tamarixia radiata Waterston (Hymenoptera: Eulophidae). pp. 251-256. In: Aubert, B., S. Tontyaporn, and D. Buangsuwon (eds.). Proceedings of the International Conference on Rehabilitation of Citrus Industry in the Asia Pacific Region. FAO; UNDP, Chiang Mai, Thailand. [ Links ]
Tsai, J.H. and Y.H. Liu. 2000. Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. J. Econ. Entomol. 93, 1721-1725. [ Links ]
Tsai, J.H., J.J. Wang, and Y.H. Liu. 2002. Seasonal abundance of the Asian citrus psyllid, Diaphorina citri (Homoptera: Psyllidae) in southern Florida. Fla. Entomol. 87, 446-451. [ Links ]
Van der Merwe, A.J. and F.G. Andersen. 1937. Chromium and manganese toxicity. Is it important in Transvaal citrus greening? Fmg. S. Afr. 12, 439-440. [ Links ]

