










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. vol.30 no.1 Bogotá jan./abr. 2012
Loyla Rodríguez1, 2 and Viviana Andrea Beltrán1
1 Plant Biotechnology Unit, Faculty of Sciences, Pontificia Universidad Javeriana. Bogota (Colombia).
2 Corresponding author. loyla.rodriguez@javeriana.edu.co
Received for publication: 9 March, 2011. Accepted for publication: 1 March, 2012.
ABSTRACT
Cordia alliodora (coffee-shading walnut) is a species of economic importance to Colombia because of its multiple uses for farmers and for reforestation as a timber and industrial resources, mainly in the central coffee region of the country. The aim of this research was to study parameters of C. alliodora shoot and root system architecture over the first year of development. Plants were established in Pacho municipality (Cundinamarca) at an 2,150 m a.s.l. A split plot design with a randomized complete block and three replicates was applied. Within the main plot, we evaluated the substrates (soil, husk and compost 2:1:1; soil and husk 3:2; soil and husk 3:1), and withing the subplot, the mycorrhizal fungus (without fungus, Kuklospora colombiana, Glomus manihotis and Acaulospora lacunosa). Substrate type affected C. alliodora root and shoot architectural parameters. Plants transplanted into soil, husk and compost 2:1:1 had the best responses in architectural parameters: diameter of the base of the root (7.82 mm), number of secondary roots (48.4), root dry weight (5.38 g), number of leaves (47.33), dry weight of shoots (7.71 g), shoot length (72.64 g) and leaf dry weight (6.28 g) at 384 days after transplant. Since no fertilizer was applied to coffee-shading walnut plants, we conclude that the mycorrhizal fungi facilitated a better use and development of mineral elements present in the substrates.
Key words: growth dynamic, development, substrate, mycorrhizal, forest species.
RESUMEN
La especie Cordia alliodora (Nogal cafetero) representa importancia económica para Colombia debido a los múltiples usos que tiene para agricultores y reforestadores, como recurso maderero e industrial principalmente en la región cafetera central del país. El objetivo de esta investigación fue estudiar parámetros de la arquitectura del sistema radical y de vástago de C. alliodora durante el primer año de desarrollo. Las plantas fueron establecidas en el municipio de Pacho (Cundinamarca) a 2.150 msnm. Bajo el diseño de parcelas divididas con distribución de bloques completos al azar con tres repeticiones, se evaluó en la parcela principal el sustrato (suelo, cascarilla y compost 2:1:1; suelo y cascarilla 3:2; suelo y cascarilla 3:1), y en la subparcela, el hongo de micorriza (sin hongo de micorriza, Kuklospora colombiana, Glomus manihotis y Acaulospora lacunosa). El tipo de sustrato afectó parámetros arquitecturales de raíz y vástago en C. alliodora, las plantas trasplantadas en suelo:cascarilla:compost 2:1:1 presentaron las mejores respuestas en los parámetros arquitecturales: diámetros de la base de la raíz (7,82 mm), número de raíces secundarias (48,4), peso seco de la raíz (5,38 g), número de hojas (47,33), peso seco de tallos (7,71g), longitud del vástago (72,64 g) y peso seco foliar (6,28 g) a los 384 días después del transplante. Considerando que no se aplicó fertilización a las plantas de nogal cafetero, los hongos de micorriza evaluados favorecieron una mejor utilización y aprovechamiento de los elementos minerales presentes en los sustratos.
Palabras clave: dinámica de crecimiento, desarrollo, sustrato, micorriza, especie forestal.
Introduction
Globally, environmental policies and actions attempt to reduce key pressures on ecosystems, the environment and biodiversity, by incorporating strategies based on sustainable management of environmental goods and services and, the efficient use of natural resources. The Colombian forestry sector should strive to minimize the negative impact of farming activities on forests and tree farms, and comply with the CONPES (2010) reforestation objectives to reduce the deforestation rate (15% annually). Therefore, the development of the forestry chain will ensure the sustainable production of forest trees and high quality products that meet the domestic demand and reduce the need to import these products (CONPES, 2010).
In the case of forest species used as timber such as Cordia alliodora (Ruiz et Pav.) Oken, farmers and forestry companies have difficulties to obtain permanent production of high quality trees to ensure their commercialization, because the environmental component is not included in the planning and development of production systems. Likewise, there are few studies about the plant growth from a dynamic and comprehensive aspect with a morphological and ecological approach (Hallé et al., 1978; Perreta and Vegetti, 2005). These situations can often be- come limiting factors for the establishment of plantations, whereas plant architecture is the result of the interaction between endogenous growth processes and the constraints exerted by the environment (Barthelemy and Caraglio, 2007; Sussex and Kerk, 2001).
C. alliodora is a forest species used in agroforestry be- cause it produces high quality wood, and is also used to provide shade for coffee plantations and pastures and for reforestation programs (Palomeque, 2009). In Colombia, Cordia alliodora is distributed in altitudes from 0 to 2,000 m on the Pacific slope, and from 0 to 700 m on the Atlantic slope, with 422 ha planted with this species in the departments of Tolima, Santander, Cundinamarca, Risaralda, Magdalena, Cauca, Valle del Cauca, Meta and Nariño (Salazar et al., 2000).
In the context of plant architecture, aspects of plant growth and development related to the shape of the plant shoot and root system are considered. This is a dynamic, global approach to growth and is the result of the operation of the plant meristems in response to environmental conditions (Perreta and Vegetti, 2005; Barthelemy and Caraglio, 2007, Sussex and Kerk, 2001; Vester, 2002). The body of a plant is formed from repetitive morphogenetic processes that create fundamental architectural units: the cell, with its ability for division allows the establishment of the meristem. Architectural plant studies are related to fundamental concepts such as architectural modeling, architectural unity and repetition. The architectural model expresses the overall growth strategy and is the set of axes that make up the body of the plant during its ontogeny (Perreta and Vegetti, 2005). The basic architectural unit structure, which allows the prescription of the plant's body shape, is metamere, formed by the insertion node, the associated leaf (or leaves), the axillary bud, the internode and in many cases the roots (Perreta and Vegetti, 2005; Sussex and Kerk, 2001). Repetition is the process that allows architectural units repeated partially or completely during ontogeny, in response to environmental conditions, to build the plant body (Perreta and Vegetti, 2005; Barthelemy and Caraglio, 2007).
The aim of this work was to study parameters of shoot and root system architecture during the first year of development of Cordia alliodora plants grown in three substrates and inoculated with three mycorrhizal fungi from three isolates of previously reported Glomeromycetes for soil in the Colombian coffee zone: Kuklospora colombiana (Instituto Venezolano de Investigaciones Científica - IVIC), Glomus manihotis (IVIC), Acaulospora lacunosa (IVIC) and a control. This architectural study on Cordia alliodora is a detailed and comprehensive approach to the development of the plant useful to propose management strategies for reforestation, forest production greenhouses, nurseries for the production of trees with excellent quality shafts, because it points to the best propagation technique, from among those compared in this study, to improve yield and mass production of high quality trees, considering that from the perspective of forest products, size and growth dynamics of the shoot and root system are determinants of the quality of the wood and the trees (Di Lucca, 1995; Garber et al., 2008; Moglia and Giménez, 2006).
Materials and methods
The present study on the root and shoot architecture of Cordia alliodora (Ruiz et Pav.) Oken was carried out during the first year of the vegetative stage. The plants were established in the town of Pacho (Cundinamarca), Bambusa station, owned by Geoambiente, located at 2,150 m a.s.l., in the dry forest foothills, with an average annual rainfall of 1,500 mm, temperature of 17.5°C and 75% relative humidity.
The experimental design was a split plot with distribution using a randomized complete block with three replications. Within the main plot, the substrate evaluated was: a mixture S1 soil: husk: compost in proportion 2:1:1 v/v/v, mixture S2 soil: husk in proportion 3:2 v/v mixture S3 soil: husk in proportion 3:1 v/v; within the subplot, we assessed mycorrhizal fungi: HM0 without mycorrhizal fungus, HM1 Kuklospora colombiana, HM2 Glomus manihotis and HM3 Acaulospora lacunosa. Each experimental unit consisted of 90 plants. Tab. 1 presents the physico-chemical properties of the tested substrates.
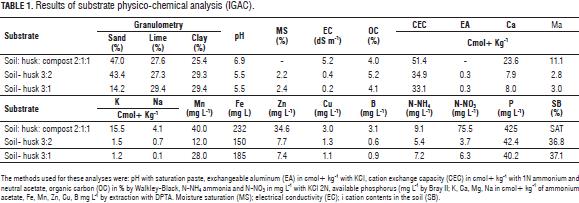
To establish the field test, C. alliodora seeds were sowed in peat. When the plants reached 8 cm in length they were transplanted to the substrates, with the application of 3 g of mycorrhizal fungi (per bag of homogenized substrate) according to the experimental design. Mycorrhizal fungi selected for the study came from three Glomeromycetes isolates previously which have been reported in soils of the Colombian coffee zone: Kuklospora colombiana (IVIC), Glomus manihotis (IVIC), Acaulospora lacunosa (IVIC), from a germplasm bank where they were propagated in Brachiaria decumbens at the Laboratory of Soil-Plant Interactions of the Plant Biotechnology Group at Javeriana University. At 169 days after transplant (dat), the roots of the plants were pruned because the evaluated substrates promoted root growth and elongation pruning was done by taking the root from the substrate and cutting the third portion of the main root apex to the neck.
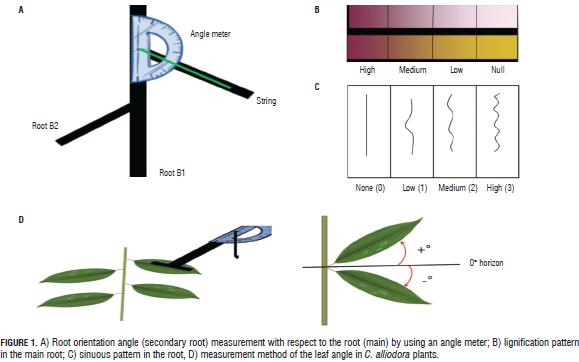
In the vegetative stage of C. alliodora (between 55 and 384 dat) destructive sampling was applied, taking two random plants per experimental unit. In the Laboratory of Plant Physiology of the PUJ, we evaluated the parameters of the root and shoot architecture: diameter of the base of the root (BD), root length (RL), number of secondary roots (B2) (Arias, 2004), orientation angle of the B2 ramification with respect to B1 (ROA) (Fig. 1a), root dry weight (RDW), leaf dry weight (LDW) and shoots dry weight (SDW), placing samples of organs in a drying oven for 48 h at 35°C, and then recording the dry weight (Hunt et al., 2002), root growth (RG) using the methodology proposed by Arias (2004), root form (RF) (Becerra et al., 2002), sinuosity (S) by means of observation and comparison with a predefined pattern: high (3), middle (2), low (1) or none (0) (Fig. 1c ) (Arias, 2004), lignification (L) in the tissues of the root with the previously established pattern of lignification (Fig. 1b) (Becerra et al., 2002), the root orientation (RO) (Arias, 2004), shoot length (SL), leaf orientation angle (LOA) with leaf angle meter (Fig. 1d), leaf length (LL), number of leaves (NL), ramification (R) according to the spatial organization of secondary axes (branches) (Sussex and Kerk, 2001; Perreta and Vegetti, 2005; Barthélémy and Caraglio, 2007), phyllotaxy (P) (Valla, 2005; Barthélémy and Caraglio 2007), growth type (GT) according to the pattern of activity of the shoot apical meristem (Sussex and Kerk, 2001; Vester, 2002; Barthélémy and Caraglio, 2007).
The statistical analyses were conducted in SAS software, the assumptions of normality and homogeneity of variances of the data were checked; because the variables NL, LOA, RL, ROA and RDW did not meet the assumption of normality, the following changes were made: NL0.7, LOA1.5 , vRL, ROA0.4 and vRDW. Analysis of variance was carried out on the parameters BD, RL, B2, ROA, RDW, LOA, SL and LDW, LL, SDW, NL. For those parameters that showed significant differences (P≤0.05) due to the effects of the substrates or mycorrhizal fungi, Duncan's test was applied for comparison of means. For variables RG, RF, S, L, RO, R, P and GT, a qualitative description was made of the obtained results.
Results and discussion
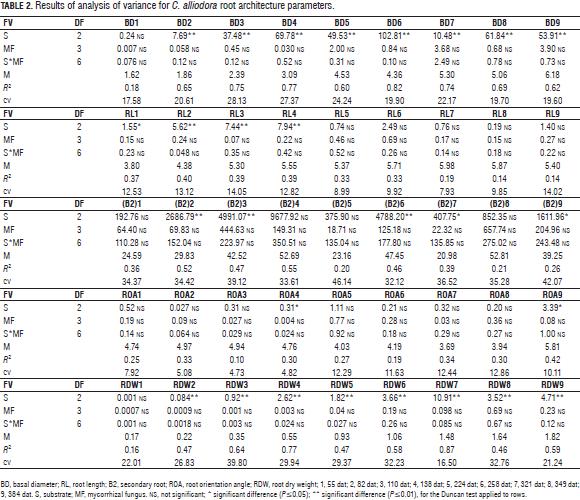
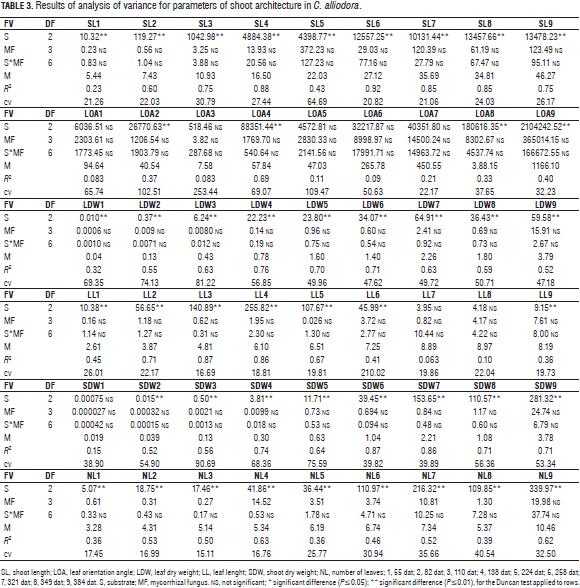
Significant differences were recorded for BD, RL, B2, ROA, RDW, SL and LDW substrate, while the interaction between mycorrhizal fungi and the substrate: mycorrhizal fungus did not show an effect on the evaluated parameters (Tabs. 2 and 3).
Effect of the substrate on the root and shoot architecture parameters in C. alliodora
The Duncan test generally defined two groups for parameters: BD, RL, B2, ROA, RDW, SL, LOA, LDW, LL, SDW and NL; group A for mixture soil: husk: compost and group B for mixtures soil: husk 3:2 and soil: husk 3:1 (Tab. 4).
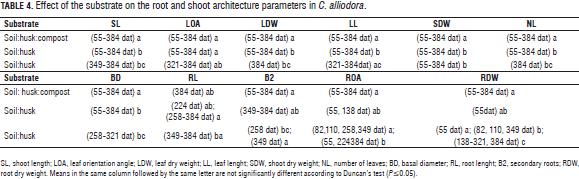
C. alliodora plants planted in soil:husk:compost had higher base diameter, longer roots, higher number of secondary roots, greater root dry weight during the evaluation period in comparison to soil: husk 3:2 and soil: husk 3:1 (Fig. 2); the mixture of soil: husk: compost possibly contributed more to establish favorable conditions for the growth of plants, containing compost, which is an organic product that contains a high content of essential mineral elements (Tab. 1) such as N-NH4 (9.1 mg L-1), N-NO3 (75.5 mg L-1), phosphorus (425 mg L-1), magnesium (11.1 cmol+ kg-1), potassium (15.5 cmol+ kg-1) which have positive effects on growth and development of C. alliodora plants (Shiralipoudre et al., 1992; Saeboa and Ferrini, 2006; Fageria et al., 2008).
Compost is a product used in urban areas for the recovery of degraded soils, containing organic compounds such as humic and fulvic acids that remain in the soil after degradation of organic matter and influence positively biological processes in the soil. The soil structure significantly improves with the application of compost, since it increases porosity and the water retention capacity, which favors development of the root apical meristem, facilitates the penetration of the root and increases the capacity of soil water storage. In addition to root growth, it increases the absorption and transport of nutrients, which leads to improve nutritional status of plants (Saeboa and Ferrini, 2006; Shiralipoudre et al., 1992).
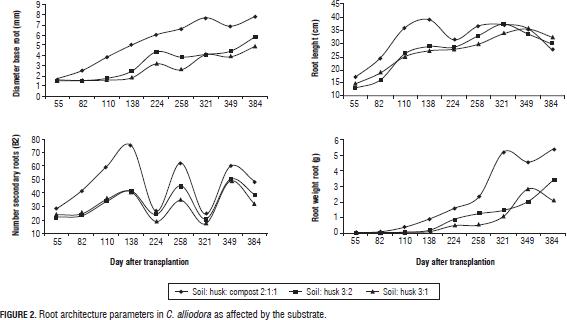
Changes in the RL, B2, RDW, and BD parameters were determined by organogenesis and morphogenetic events which allow the establishment of the root of C. alliodora plants (Sussex and Kerk, 2001; Vester, 2002; Barthélémy and Caraglio, 2007; Perreta and Vegetti, 2005). These changes occurred in two successive stages of root growth (Fig. 2), the logarithmic phase that occurred between 55 and 82 dat and was characterized by an increase in the speed of B1 and B2 root growth and accumulation of root dry weight. With time, at this stage, organogenesis was the result of the operation activity of the undifferentiated cells of the root apical meristem, likewise at this stage, lateral ramification of the root began (Barthélémy and Caraglio, 2007; Lynch, 1995). In the second phase, with exponential growth of the root, which occurred between 82 and 384 dat, processes related to the ramification of the root continued. On average, there were 20 B2 roots, determining the root polytomy (Barthélémy and Caraglio, 2007; Perreta and Vegetti, 2005; Lynch, 1995). At 224 dat (Fig. 2), there was a reduction of the parameters related to the extension and ramification of the root: mainly RL and B2, caused by root pruning, a practice that was necessary considering the excessive root growth of C. alliodora plants.
In the substrate soil: husk: compost 2:1:1, the reduction in RL and B2 was highly significant. This is explained because this substrate led to greater root growth between 52 and 138 dat. therefore, root pruning at 169 dat allowed a larger proportion of root reduction in comparison to the other tested substrates. Likewise, the plant required more time to recover from this management technique, whereby the RL at 384 dat was lower than the other treatments. According to several authors, root pruning is a nursery technique that allows control of the root growth pattern. It is usually used to increase the survival of transplanted trees. Root pruning has an important potential to control the root system growth at a relatively small volume and to increase the number of hairs (Low et al., 2011; Watson and Davis, 1987). Gilman (1992) states that each plant species has a determined characteristic shoot/root ratio; when changing the ratio with pruning techniques, the plant responds by redistributing the photo-assimilates to replace removed organs.
The angle of orientation of B2 with respect to B1 (ROA) and root orientation in C. alliodora were not generally affected by the substrates, so we can say that these specific parameters of root architecture are non plastic traits, that is, they are not influenced by the soil conditions (Chambell et al., 2005; De Kroon et al., 2005; Sultan, 2003). The orientation of B1 was determined as orthogravitropic, considering that the extension of the taproot (B1) was positively guided by gravity, while the B2 axes were unevenly distributed about the B1 axis with orientation angles less than 83° (Tab. 5, Fig. 3), because the B2 axes originated from the main root and oriented with positive direction to gravity, as would gravitropics (Arias, 2004).
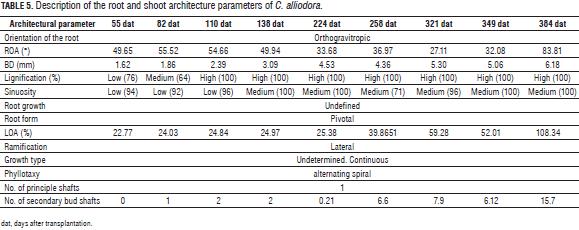
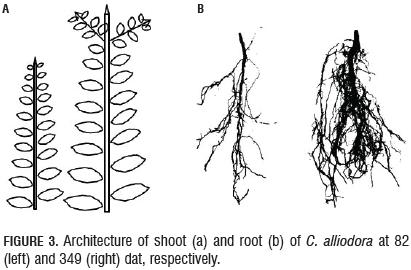
The qualitative characteristics of the root growth response, root form and sinuosity were not influenced by the substrate type during the 384 dat; then, these traits were considered non-plastic (Chambell et al., 2005; De Kroon et al., 2005; Sultan, 2003). The B1 axis of the root showed undefined growth during the 384 dat. In general, the root architecture parameters RL, BD and RDW increased between two successive samples (Fig. 2) (Arias, 2004; Barthélémy and Caraglio, 2007; Perreta and Vegetti, 2005). The root form of C. alliodora was defined as pivotal wherein the B1 axis was well differentiated, long and thick, and grew faster than the B2 roots (Fig. 3b). The sinuosity degree of B1 axis in the root of C. alliodora was not influenced by the substrates and was low until 110 dat and medium from 138 to 384 dat (Tab. 5).
Although, the degree of lignification of root axis B1 was not affected by the substrate, it increased through the vegetative stage; low until 55 dat, medium until 82 dat, and high until 384 dat (Tab. 5); during the growth cycle of woody plants, the deposit of lignin in the cell walls is necessary. Lignin is a phenolic compound with functional significance for the plant because it confers mechanical support (structural rigidity and durability) to plant tissues, enhancing lignification of the vascular elements and thereby increasing sap conduction, and it is also associated with the plant defense mechanisms (Boudet, 2000; Cervilla et al., 2009).
The shoot architecture parameters: length of shoot, leaf length, leaf dry weight and shoot dry weight, was influenced by the substrate, with the mixture soil: husk: compost showing the highest readings (Tabs. 3 and 4) due to the presence of compost, an organic product that increases shoot biomass because it provides greater amounts of essential nutrients N, P, K, Ca, Mg, S, Fe, Zn, Cu and Mn (Tab. 1) for root and organ growth in C. alliodora (Saeboa and Ferrini, 2006); Rivero et al. (2004) argue that root growth increases stability of trees, because a larger volume of soil explored by the roots leads to a better nutritional status and less plant stress, likewise, compost improves the quality of soil organic matter, contributing to a higher content of humic compounds in the soil, maintaining a balance between the microorganisms present in the substrate (Rivero et al., 2004; Pascual et al., 2002). Furthermore, this substrate contains rice husks, which provide better drainage and maintain a uniform temperature in the ground, ensuring uniform distribution of soil moisture and allowing good soil aeration (Rodríguez, 2007).
In the initial period of growth until 138 dat, shoot organs in C. alliodora plants form part of the organogenesis and extension morphogenetic processes of the shoot and leaves to form metamere, basic morphological units to shoot formation in C. alliodora (Perreta and Vegetti, 2005; Volkenburgh, 1999). According to Barthélémy and Caraglio (2007), in the active growth phase (logarithmic), the shoot apical meristem develops new leaves and shoots, which then elongate to form metamere. Subsequently, between 138 and 384 dat, the processes of overlapping and reiteration in metamere allow the construction of the shoot growth model in C. alliodora (Fig. 3) (Perreta and Vegetti, 2005).
During leaf ontogeny, the orientation angle of the leaves was affected by the substrates (Tabs. 3 and 4) with the mixture soil: compost: husk having the highest influence; the compost probably promoted greater accumulation of organic compounds in the leaves, which increased the LOA (Saeboa and Ferrini, 2006). Between 55 and 384 dat, leaf lamina development events occurred. As the growth cycle of the plants unfolded, dry matter was translocated and accumulated in the leaves as processes of cell division and expansion which are essential for the formation of leaf biomass. Therefore, an increase in LDW was also seen (Fig. 4), leading to the leaves descending due to increased biomass, hence, the leaf angle with the horizontal line increased as the leaves were growing (Volkwnburg, 1999).
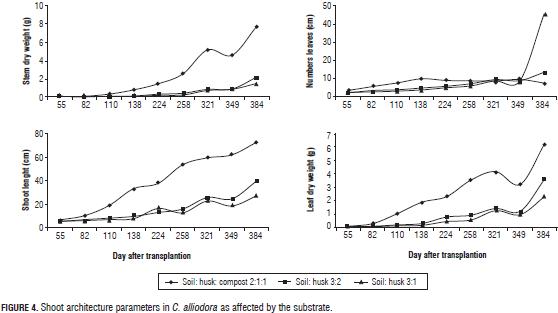
During the 384 dat evaluated, the architectural pattern of C. alliodora corresponds to the "Fagerlind" model proposed by Vester (2002) for Cordia, which is characterized by a single sympodial orthotropic axis with continued growth and the LOA was plagiotropic with angle orientations of 108° on average for the 384 dat.
C. alliodora presented lateral ramification, with the presence and organization of secondary axes towards the apex of the shoot, the leaf arrangement along the principle axis or phyllotaxy was determined as alternating spiral because at each metamere, a leaf is inserted at each node, in an alternating spiral arrangement along the principal axis (Fig. 3, Tab. 5). According to the pattern of activity of the apical meristem shoot in C. alliodora, the growth rate was established as indeterminate and continuous, i.e. the apical meristem of the shoot did not transform into another structure (inflorescence, tendril, parenchyma ball, etc.) that would stop the growth of the plant, constant shoot growth was observed throughout the evaluation period of 384 dat (Fig. 3) (Vester, 2002; Perreta and Vegetti, 2005; Barthélémy and Caraglio, 2007; Moglia and Giménez, 2006).
Effect of interaction substrate:mycorrhizal fungus on the root and shoot architecture parameters in Cordia alliodora
In the interaction of the three substrates: mixture soil: husk: compost 2:1:1, soil: husk 3:2 and soil: husk 3:1 with mycorrhizal fungi: HM0 (without mycorrhizal fungus), Kuklospora colombiana, Glomus manihotis and Acaulospora lacunosa no statistical differences were seen for any of the variables from the root and shoot architecture evaluated during the 384 dat, because the effects of the application of mycorrhizal fungi did not invoke an immediate response in the architectural parameters evaluated (Tabs. 2 and 3); however, it is important to highlight the advantages of the use of mycorrhizal fungi in agroforestry systems: the use of arbuscular mycorrhizae in tree farms and agricultural systems contribute to mineral nutrient uptake by the plant due to a greater volume of rhizosphereic soil due to mycorrhizal hyphae (Schuler et al., 2001).
Because no fertilizer was applied to the plants, the results indicate that the evaluated mycorrhizal fungi promoted a better use and development of mineral elements present in the substrates. Therefore, for sustainable use and management of resources in production systems, the use of mycorrhizal fungi is important, which are highly effective in compensating for the reduction or removal of chemical inputs. Similarly, more mobile ions present in the soil solution such as NO3, P, Zn, Cu and Mo, and to a less extent K and S, are more readily available to the plant as a result of mycorrhizae. The absorption of less mobile mineral elements depends on the area of soil encompassed by the roots of the plant; in this case, mycorrhizal roots have advantages over non-mycorrhizal roots because the external mycelium extends farther than the root hairs (Guerra, 2008).
Root and shoot architectural parameters of C. alliodora plants presented in the first two stages of plant growth curves: a logarithmic phase that occurred until 138 dat for BD, RL, RDW, until 224 dat for SDW, until 321 dat for NL, until 224 dat for SL and until 258 dat for LDW; subsequently an exponential growth stage for these variables occurred until 384 dat (Figs. 5 and 6).
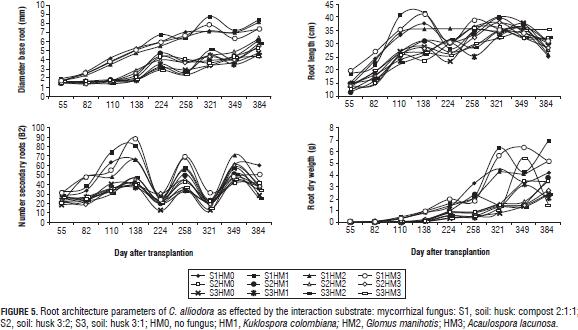
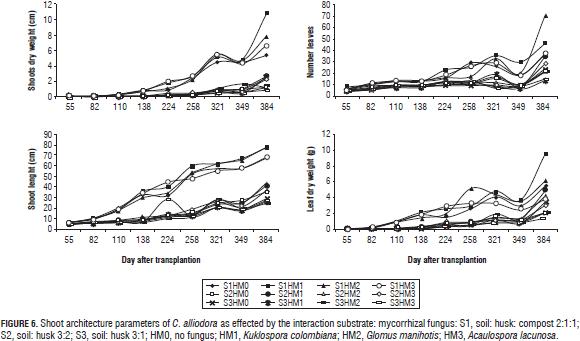
In general, plants grown in the substrate soil: husk: compost interacting with Kuklospora colombiana, Glomus manihotis and Acaulospora lacunosa presented higher averages than other interactions. Studies conducted to evaluate the effect of combined application of organic substrates and mycorrhizal fungi on plant growth claim that organic substrates favor mycorrhizal association (Franco et al., 2008; Cuenca et al., 2007; Maciel et al., 2002; Wang and Yong-Shi, 2008), leading to changes in the root structure with more numerous, longer lateral roots (De Luca et al., 2007; Blanco and Salas, 1997; Calderón et al., 2000). Redel et al. (2006) and Maciel et al. (2002) state that the use of mycorrhizal fungi provides nutritional benefits to the plant when there is an application of chemical fertilizers, which are reflected in higher growth. This is confirmed by Donoso et al. (2008) who also clarifies that the use of mycorrhizae with the application of an organic fertilizer such as compost greatly favors growth.
The parameters B2 and RL increased up to 138 DAT due to the evaluated interaction substrate:mycorrhizal fungus, however at 224 dat they degreased as a result of root pruning on the C. alliodora plants (Fig. 5).
The types of ramification and phyllotaxy of Cordia alliodora were not affected by the interaction substrate: mycorrhizal fungus, which is considered as non-dynamic parameter, the phyllotaxy was determined as lateral (Fig. 3) and the type of growth in accordance with the pattern of activity in the apical meristem was established as indeterminate and continuous (Palacio and Rodríguez, 2007).
Conclusions
The substrate type affected the response of the shoot and root architecture parameters of C. alliodora, with the mixture soil: husk: compost (2:1:1) having contributed to the best conditions for growth and development of the plants. The application of the mycorrhizal fungi Kuklospora colombiana, Glomus manihotis and Acaulospora lacunosa did not affect the shoot and root architecture parameters of C. alliodora.
Acknowledgments
The authors express their gratitude to Geoambiente for providing the nursery area to carry out this study; Biology Program students: María Camila Villegas Infante, Laura Villamizar Carrillo, Esyevit Karina González Roa for contributing to the processing of the plant material and Fabio Guacaneme Castiblanco for assistance with the statistical analysis. This study was funded by the Vicerrectoría Académica de la Pontificia Universidad Javeriana.
Literature cited
Arias, T. 2004. Arquitectura de raíces y vástagos de Vismia baccifera y de raíces de Vismia macrophylla (Clusiaceae). Caldasia 26(2), 333-358. [ Links ]
Barthélémy, D. and Y. Caraglio. 2007. Plant architecture: a dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. Ann. Bot. 99(3), 375-407. [ Links ]
Becerra, N., X, Marquínez, and E. Barrera. 2002. Anatomía y morfología de los órganos vegetativos de las plantas vasculares. Universidad Nacional de Colombia, Bogota. [ Links ]
Blanco, F. and E. Salas. 1997. Micorrizas en la agricultura: contexto mundial e investigación realizada en Costa Rica. Agron. Cost. 21(1), 55-67. [ Links ]
Boudet, A. 2000. Lignins and lignification: selected issues. Plant Physiol. Biochem. 38(1/2), 81-96. [ Links ]
Calderón, L., L. Gómez, F. Blanco, and L. Uribe. 2000. La inoculación con Glomus manihotis sobre el crecimiento y desarrollo de plantas de yuca producidas in vitro, en la fase de aclimatización. Agron Cost. 24(2), 25-29. [ Links ]
Cervilla, L., M. Rosales, M. Rubio-Wilhelmi, E. Sánchez-Rodríguez, B. Blasco, J. Rios, L. Romero, and J. Ruiz. 2009. Involvement of lignification and membrane permeability in the tomato root response to boron toxicity. Plant Sci. 176, 545-552. [ Links ]
Chambell, M.R., J. Climent, R. Alía, and F. Valladares. 2005. Phenotypic plasticity: a useful framework for understanding adaptation in forest species. Invest. Agrar. Sist. Recur. For. 14(3), 334-344. [ Links ]
CONPES, Consejo Nacional de Política Económica y Social. 2010. Política nacional de competitividad y productividad. Document 3668. Bogota. [ Links ]
Cuenca, G., A. Cáceres, G. Oirdobro, Z. Hasmy, and C. Urdaneta. 2007. Las micorrizas arbusculares como alternativa para una agricultura sustentable en áreas tropicales. Interciencia 32(1), 23-29. [ Links ]
De Kroon, H., H. Huber, J. Stuefer, and J. Van Groenendael. 2005. A modular concept of phenotypic plasticity in plants. New Phytol. 166, 73-82. [ Links ]
De Luca, L., C. Flores, and Y.S. Sarandon. 2007. Incidencia de sustratos de fabricación casera sobre el índice de micorrización y la morfología radical en plantines de tomate. Rev. Bras. Agroec. 2(2), 1016-1020. [ Links ]
Di Lucca, M. 1995. Curso de modelos de crecimiento y producción forestal. INTA, Bariloche, Argentina. [ Links ]
Donoso, E., G. Lobos, and N. Rojas. 2008. Efecto de Trichoderma harzianum y compost sobre el crecimiento de plántulas de Pinus radiata en vivero. Bosque 29(1), 52-57. [ Links ]
Fageria, N., V. Baligar, and Y. Li. 2008. The role of nutrient efficient plants in improving crop yields in the twenty first century. J. Plant. Nut. 31(6), 1121-1157. [ Links ]
Franco, A., D. Castañeda, C. Úsuga, F. Gómez, and C. Lopera. 2008. Efecto de la micorrización y la fertilización en la acumulación de biomasa en plantas de banano (Musa Aaa cv. Gran Enano) (Musaceae). Rev. Fac. Nac. Agron. Medellin 61(1), 4269-4278. [ Links ]
Garber, S., R. Monserud, and D. Maguire. 2008. Crown recession patterns in three conifer species of the Northern Rocky Mountain. For. Sci. ProQ. Agr. J. 54(6), 628-633. [ Links ]
Gilman, E. 1992. Effect of root pruning prior to transplanting on establishment of southern magnolia in the landscape. J. Arboric. 18(4), 197-201. [ Links ]
Guerra S., B.E. 2008. Micorriza arbuscular. Recurso microbiológico en la agricultura sostenible. Tec. Marcha 21(1), 191-201. [ Links ]
Hallé, F.R., R.A. Oldeman, and P.B. Tomlinson. 1978. Tropical trees and forests. An architectural analysis. Springer, Berlin. [ Links ]
Hunt, R., D.R. Causton, B. Shipley, and P. Askew. 2002 .A modern tool for classical plant growth analysis. Ann. Bot. 90, 485-488. [ Links ]
Lynch, J. 1995. Root architecture and plant productivity. Plant Physiol. 109, 7-13. [ Links ]
Low, M., L. Whitfield, A. Rutter, and B.A. Zeeb. 2011. The effects of pruning and nodal adventitious roots on polychlorinated biphenyl uptake by Cucurbita pepo grown in field conditions. Env. Pol. 159, 769-775. [ Links ]
Maciel, N., N. Mogollón, and C. Cásares. 2002. Efectos del sustrato y la fertilización en el crecimiento de Spathiphyllum 'Petite'. Proc. Interam. Soc. Trop. Hort. 46, 103-105. [ Links ]
Moglia, J. and A.M. Giménez. 2006. Análisis de la arquitectura vegetal: resultados preliminares de la arquitectura vegetal de Prosopis alba y P. nigra. Facultad de Ciencias Forestales, Universidad Nacional de Santiago, Santiago. [ Links ]
Palacio, K. and N. Rodríguez. 2007. Plasticidad fenotípica en Lippia Alba (Verbenaceae) en respuesta a la disponibilidad hídrica en dos ambientes lumínicos. Act. Biol. Col. 12(5), 187-198. [ Links ]
Palomeque, E. 2009. Sistemas agroforestales, una alternativa del campo. In: http://www.monografias.com/trabajos-pdf2/sistemas-agroforestales/sistemas-agroforestales.pdf; consulted: March, 2012. [ Links ]
Pascual, J.A., C. García, T. Hernández, S. Lerma, and J.M. Lynch. 2002. Effectiveness of municipal waste compost and its humic fraction in suppressing Pythium ultimum. Mic. Eco. 44, 59-68. [ Links ]
Perreta, M.G. and A. Vegetti. 2005. Patrones estructurales en las plantas vasculares: una Revisión. Gay. Bot. 62(1), 9-19. [ Links ]
Redel, Y., R. Rubio, and F. Borie. 2006. Efecto de la adición de residuos de cosecha y de un hongo micorrizógeno sobre el crecimiento de trigo y parámetros químicos y biológicos de un Andisol. Agric. Tec. 66(2), 174-184. [ Links ]
Rivero, C., T. Chirenje, G. Martínez. 2004. Influence of compost on soil organic matter quality under tropical conditions. Geoderma 123, 355-361. [ Links ]
Rodríguez, G. 2007. Efecto de la cobertura del suelo con cascarilla de arroz en el crecimiento y rendimiento del tomate de ramillete. Cien. Inv. Ag. 34(3), 225-230. [ Links ]
Saeboa, A. and F. Ferrini. 2006. The use of compost in urban green areas. A review for practical application. Urb. For. Urb. Green. 4, 159-169. [ Links ]
Salazar, R., C. Soihet, and J. Méndez. 2000. Manejo de semillas de 100 especies forestales de América Latina. CATIE, Turrialba, Costa Rica. [ Links ]
Shiralipoudre, A., D. Mcconnelaln, and S. Smith. 1992. Uses and benefits of MSW compost: a review and an assessment. Biom. Bioen. 3(34), 267-279. [ Links ]
Schuler, A., D. Schwarzott, and C. Walker. 2001. A new fungal phylum, the Glomeromycota: phylogeny and evolution. Mycol. Res. 105, 1413-1421. [ Links ]
Sultan, S. 2003. Phenotypic plasticity in plants: a case study in ecological development. Evol. Develop. 5(1), 25-33. [ Links ]
Sussex, I.M. and N.M. Kerk. 2001. The evolution of plant architecture. Ann. Bot. 5, 33-37. [ Links ]
Valla, J.J. 2005. Botánica: morfología de las plantas superiores. Hemisferio Sur, Buenos Aires. [ Links ]
Volkenburgh, V.E. 1999. Leaf expansion - an integrating plant behaviour. Planta 22, 1463-1473. [ Links ]
Vester, H. 2002. Modelos arquitectónicos. En la flora arbórea de la península de Yucatán. Bol. Soc. Bot. Méx. 71, 45-57. [ Links ]
Wang, F.Y. and Z. Yong-Shi. 2008. Biodiversity of arbuscular mycorrhizal fungi in China: a Review. Adv. Env. Biol. 2(1), 31-39. [ Links ]
Watson, G. and T. Davis. 1987. The effect of root pruning on the root system of nursery trees. J. Arboric. 13(5), 126-130. [ Links ]
