










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.30 no.2 Bogotá May/Aug. 2012
CROP PROTECTION
Screening of microbial culture filtrates, plant extracts and fungicides for control of mango anthracnose
Screening de filtrados de cultivos microbianos, extractos vegetales y fungicidas para el control de la antracnosis del mango
Jairo A. Osorio1 Erika P. Martínez1and Juan C. Hío1
1Plant Pathology Laboratory, Tibaitatá Research Center, Corporación Colombiana de Investigación Agropecuaria (Corpoica). Mosquera (Colombia). josorio@corpoica.org.co. Received for publication: 23 May, 2011. Accepted for publication: 29 June, 2012.ABSTRACT
Anthracnose (Colletotrichum gloeosporioides) induces losses of up to 40% in most mango growing regions in Colombia. A series of exploratory experiments were carried out to evaluate 14 potentially antagonistic microorganisms, six plant extracts and 10 fungicides for disease control based on their ability to inhibit spore germination, mycelium growth and development of symptoms on detached fruits. In vitro experiments showed that spore germination of C. gloeosporioides was strongly inhibited (P≤0.0001) by all microbial culture filtrates, all fungicides and two plant extracts, suggesting that some of these treatments may hold promise for disease control. Similarly, all microbial filtrates, most fungicides and two plant extracts caused a significant (P≤0.0001) inhibition in mycelium growth of the pathogen. Results from this study indicated small effects of fungicides or plant extracts on anthracnose development on inoculated fruits, while microbial filtrates induced a significant reduction (P≤0.05) in lesion size. These results are very promising since current control practices rely on fungicides or costly physical treatments.
Key words: Colletotrichum gloeosporioides, fruit protection, non-conventional control methods.
RESUMEN
La antracnosis (Colletotrichum gloeosporioides) induce pérdidas de hasta 40% en la mayoría de zonas productoras de mango en Colombia. Se realizó una serie de experimentos exploratorios para evaluar 14 antagonistas potenciales, seis extractos vegetales y 10 fungicidas en el control de la enfermedad, con base en su capacidad para inhibir la germinación de esporas, el crecimiento del micelio y el desarrollo de síntomas en frutos desprendidos. Los experimentos in vitro mostraron que la germinación de esporas de C. gloeosporioides fue inhibida (P≤0,0001) por todos los filtrados crudos microbianos, todos los fungicidas y dos de los extractos vegetales; indicando que algunos de estos tratamientos pueden ser promisorios para el control de la enfermedad. Asimismo, todos los filtrados microbianos, varios fungicidas y dos extractos vegetales causaron una inhibición significativa (P≤0,0001) en el crecimiento del micelio del patógeno. Los resultados de este estudio mostraron un bajo efecto de los fungicidas ó los extractos vegetales en el desarrollo de antracnosis en frutos inoculados, mientras que los filtrados microbianos mostraron una reducción significativa (P≤0,05) en el tamaño de lesión. Esto es muy promisorio pues las prácticas actuales de control se basan en fungicidas ó tratamientos físicos costosos.
Palabras clave: Colletotrichum gloeosporioides, protección de frutos, control no convencional.
Introduction
Several varieties of mango (Mangifera indica, L.) such as Haden, Irwin, Keitt and Tommy Atkins are grown in Colombia, as well as local varieties that are used in the industry (MA DR, 2009). Mango orchards in all regions are affected by anthracnose (Colletotrichum gloeosporioides [Penz.] Penz. & Sacc., teleomorph: Glomerella cingulata [Stoneman] Spaulding & Von Schrenk); this disease induces field losses of up to 40% and postharvest losses of up to 70% in the most susceptible cultivars (Páez-Redondo, 1995; Afanador et al., 2003; MA DR, 2009) in Mexico, India, Australia and South Africa (Chowdhury et al., 2007; Kumar et al., 2007; Rojas-Martínez et al., 2008). Typical symptoms of anthracnose are sunken, necrotic, dark brown to black lesions on mango fruit (Bailey et al., 1992). The size and shape of these lesions vary with the cultivar, amount of quiescent infections and age of the infected organs (Arauz, 2000; Beno-Moualem and Prusky, 2000; Wharton and Diéguez-Uribeondo, 2004; Osorio, et al., 2006).
Various anthracnose control methods are used by farmers including fungicides, calcium carbonate, antagonists and sanitation (Benítez et al., 2004; Sundravadana et al., 2006; Korsten, 2006; Korsten et al., 1997; Stockwell and Stack, 2007). In Colombia, fungicide sprays and occasional sanitation are used by mango growers once symptoms of the disease are visible, but the effectiveness of these measures is limited (Osorio et al., 2000; Osorio et al., 2006; Castro et al., 2007; Moral et al., 2009).
Fungicide tolerance in populations of C. gloeosporioides was reported more than 20 years ago (Liyanage et al., 1992), and has resulted in reduced biological activity and increased fungicide residues on harvested fruits. This has prompted efforts to develop alternate methods for anthracnose control on various crops (Cheng et al., 2006), including prediction models based on weather information (Dodd, 1991; Arauz, 2000). Methods used for post-harvest anthracnose control include fungicides, biocides, calcium carbonate, chemical inducers, microorganisms (Benítez et al., 2004), hyperbaric pressure treatments, antioxidants, radiation, heat shock and, -at the experimental stage- genetic manipulation (Korsten, 2006).
Three recent studies carried out by Corpoica examined the survival strategy and population structure of the anthracnose pathogen (C. gloeosporioides) that affects mango orchards in Colombia (Ospina and Osorio, 2005; Cadavid et al., 2007; Martínez et al., 2009) and tamarillos (Reyes et al., 2007); the results suggested the need to explore diverse alternative control methods for this disease. Here, we report the results from laboratory studies carried out to determine the biological activity of microbial antagonists, plant extracts and fungicides against C. gloeosporioides; focusing on the source of the inoculum and symptom suppression in mature fruits.
Pathogen isolates. Two single spore isolates (Cg-917 and Cg-935), obtained from "Hilacha" mango fruits harvested in Coello (Tolima, Colombia) and identified as C. gloeosporioides by morphology, growth on selective media and species-specific rDNA amplification profiles, were used in this study. The two isolates were similar in spore morphology, colony growth habit, pathogenicity and SS R (microsatellites) profiles, according to previous studies (Cadavid et al., 2007). The two isolates also showed similar germination and mycelium growth responses to fungicides, plant extracts and culture filtrates in exploratory trials.
Microorganisms and liquid culture filtrates. Isolates of potential antagonists were obtained from field samples (leaves, flowers, fruits) collected between October/2006 and March/2007 at two locations (Sevilla, Magdalena and Espinal/Guamo, Tolima). The plant material was processed for microbial isolation according to published protocols (Paterson and Bridge, 1994; Barnett et al., 2000; Schaad, 2001). The fungal, yeast and bacterial colonies were purified and stored at -20°C in 20% glycerol stock solutions. Identification of fungal and bacterial isolates was made by morphological comparisons and standard biochemical tests, respectively.
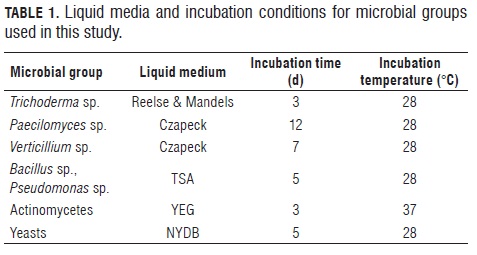
Five liquid media were prepared to obtain crude culture filtrates: reelse and mandels medium for Trichoderma, Czapeck medium for Verticillium and Paecilomyces growth, TSA medium for bacteria, YEG medium for actinomycetes and NY BB medium for yeasts. Pure colonies of each isolate were used to inoculate 500 mL flasks containing 250 mL of a specific liquid media, before incubation in an orbital shaker, following specific protocols (Tab.1) (Paterson and Bridge, 1994; Donadio et al, 2002; Abril et al., 2008). Following incubation, microbial biomass was separated by Filtration through cheesecloth, centrifugation (8000 g, 20 min) and filtration through millipore membrane filters (0.1 or 0.3 micrometers in diameter, according to the type of microorganism).
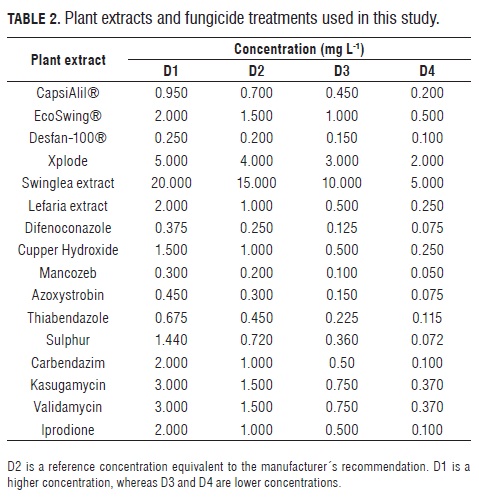
Plant extracts. Five commercially available plant extracts were obtained from local suppliers: CapsiAlil® (Sulfur compounds of Liliaceaes+Protoalcaloides of Solanaceous), Eco- Swing® (terpeniods from Swinglea glutinosa), Desfan-100® (Citrus seed extract), Xplode SL (capsaicin+alisin), swinglea extract (Terpeniods). Additionally, an aqueous extract of the xeric lefaria cactus (Cereus deficiens) was prepared by grinding and drying tissue from the middle portion of fully extended branches. A 20 g sample was fully homogenized in 400 mL of distilled water and filtered through cheesecloth before use. Four different concentrations of each extract were evaluated.
Fungicides. Ten commercial fungicides were used in this study: Score® 250EC (difenoconazole), Kocide® 101 (copper hydroxide), Alarm® 80WP (mancozeb), Amistar® 50WG (azoxystrobin), Mertec® 500SC (thiabendazole), Elosal® 720SC (sulphur), Carbendazim AC® (carbendazim), Kasumin ® 2SL (kasugamycin), Validamycin (validamycin) and Rovral® 5OWP (iprodione). Four concentrations of each fungicide were evaluated.
Spore germination assays. Exploratory tests were performed with isolates Cg-917 and Cg-935 in order to determine spore germination and standardize incubation conditions (Cerón et al., 2005). Fresh colonies of the two isolates were produced on Marthur´s medium incubated at 28°C in darkness for 10 d. Spore suspensions (1·105 spores/ mL) were prepared on either a) sterile water amended with Tween®-80 (0.05% v/v) or b) a 10% aqueous sucrose solution amended with Tween-80 (0.05% v/v); and incubated at 28°C and 100% relative humidity on glass slides. Germination counts were carried out every 2 h and for up to 24 h, germination was higher on water (10-15%) as compared to the sucrose solution (3%). Based on these results, all germination assays used 1.5 mL of spore suspensions of 1·105spores/mL in water + Tween®-80 (0.05% v/v), using Eppendorf tubes incubated at 27°C for 18 h.
A completely randomized design with three repetitions was used to compare the inhibitory activity of crude filtrates from liquid cultures of 14 microbial isolates (six fungi, four bacteria, two actinomycetes, and two yeasts) on spore germination of the isolate Cg-917. For this, spore suspensions were mixed at a 1:1 ratio with the culture filtrates in 2 mL Eppendorf tubes, and incubated as described above. Following incubation, 50 µ:L were taken from each tube and poured into a Neubauer chamber to determine germination.
A randomized block design displayed in a factorial arrangement with three repetitions was used to compare six plant extracts in four concentrations (Tab.2) by their inhibitory activity on spore germination of the isolate Cg-935 of the mango pathogen. Spore suspensions prepared as described were mixed at a 1:1 ratio with each one of the plant extract treatments, incubated and evaluated as described above.
To test the efficacy of ten fungicides, at different concentrations (Tab.2), to inhibit spore germination of the C. gloeosporioides isolate Cg-935, a completely randomized design displayed in a factorial arrangement with four repetitions was used. Spore suspensions were mixed at a 1:1 ratio with each one of the fungicide treatments, incubated and evaluated as described above.
Mycelium growth assays. A completely randomized design with three repetitions was used in an experiment to test the effect of culture filtrates from 14 microbial isolates (six fungi, four bacteria, two actinomycetes, and two yeasts) on mycelium growth of the C. gloeosporioides isolate Cg-917. Petri plates were prepared by mixing 9 mL of PDA with 1 mL of culture filtrate and left to solidify, inoculated with 5-mm disks of PDA + mycelium of Cg-917 and incubated at 27°C in the dark for 7 d. After this period, the colony diameter was measured.
Mycelium growth of the isolate Cg-935 in the presence of plant extracts (Tab.2) was determined in a randomized block experiment with a factorial arrangement and three repetitions. Petri plates were prepared by mixing 9 mL of PDA with 1 mL of plant extract at each concentration. Once solidified, 5-mm disks of PDA with actively growing Cg-935 mycelium were placed in the center of each plate. Inoculated plates were incubated at 27°C in the dark for 7 d, at which time the colony diameter was measured.
The effect of ten fungicides at different concentrations (Tab.2) on mycelium growth of the C. gloeosporioides isolate Cg-935 was tested in a completely randomized experiment with a factorial arrangement and four repetitions.
Petri plates were prepared by mixing 9 mL of PDA with 1 mL of fungicide solution. Once solidified, 5-mm disks of PDA with actively growing mycelium of either isolate of C. gloeosporioides were placed in the center of each plate. Inoculated plates were incubated at 27°C in the darkness for 7 d, at which time the colony diameter was measured.
Fruit protection assays. The best treatments from all previous experiments were tested for anthracnose control on detached mango fruits. A completely randomized experiment replicated three times was used to evaluate the protection ability of 14 microbial filtrates on fruits inoculated with the isolate Cg-917 of the anthracnose pathogen. Similarly, a completely randomized experiment in a factorial arrangement replicated three times was used to test the effectiveness of five fungicides on fruits inoculated with the isolate Cg-935 of the mango pathogen. Likewise, a completely randomized factorial experiment was used to test the effectiveness of two plant extracts at two concentrations each at preventing anthracnose development on fruits inoculated with the isolate Cg-935.
Mango fruits (hilacha cultivar, E2 growth stage) were washed, superficially disinfected with 1% NaOCl, and rinsed in distilled water. Treatments of fruits with culture filtrates from antagonists, plant extracts or fungicide solutions consisted of a 30-min immersion, followed by air drying at room temperature. Treated fruits were then inoculated by placing two 5-mm disks of a sporulating colony of the pathogen on the fruit surface (5 cm apart). After inoculation, the fruits were kept inside plastic boxes (two fruits per box) containing moist paper towels, at 25ºC in alternating 12/12 dark/light conditions. Lesion diameter, resulting from inoculation, was assessed after 8 d.
Data analysis. All statistical analyses were performed with SAS ® version 9.3. Standard AN OVA and Tukey Studentized Range Test (HSD) for mean separation (P≤0.05) were performed on data from spore germination and mycelium growth experiments. Similarly, data from fruit protection assays were analyzed by AN OVA and treatment means were separated using HSD (P≤0.05).
Results
Biological activity of microbial culture filtrates. Four exploratory tests were initially carried out in 2007 with four microbial groups (bacteria, fungi, actinomycetes, yeasts), resulting in 14 isolates being selected for further testing against the pathogen C. gloeosporioides isolate Cg-917 (Tab.3). Culture filtrates from all 14 isolates reduced spore germination (P≤0.0001) in the range of 43 to 69% as compared to the control treatment; and some of them induced germ tube malformations (data not shown).The Tukey (HSD) test indicated greater effects for the isolates Actin-38, Bacte-15, Bacte-40, Bacte-59, Paeci-85 and Trich-19. Mycelium growth of the pathogen on filtrate-amended medium was reduced by all microbial isolates (P≤0.0001), with greater reductions (90- 95% colony growth reduction) induced by the isolates Actin-35, Bacte-59, Bacte-40, Bacte-73 and Yeast-05 (Tab.3).
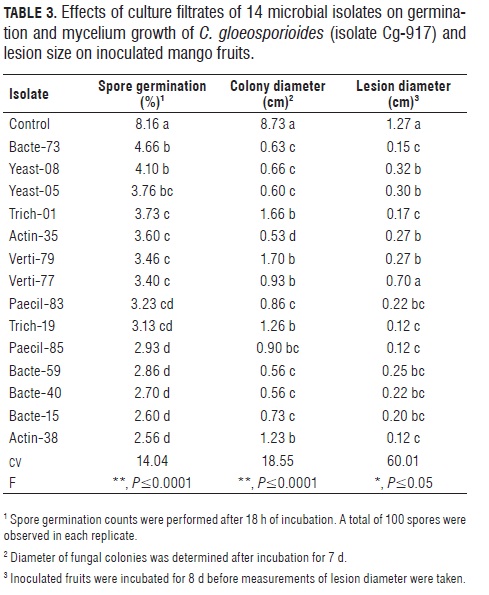
Mango fruits that were superficially sprayed with culture filtrates of five isolates (Paeci-85, Trich-19, Trich-1, Bacte-73 and Actin-38) before inoculation with Cg-917, showed lesion size reductions around 80% when compared with the untreated control (P≤0.05); however, caution should be exercised when interpreting these results due to the high variation coefficient in this experiment. This suggests the need for additional studies with a larger number of fruits to take into consideration factors like the presence of quiescent infections on the surface and the nutritional status of fruit tissues, which lead to intrinsic variability among mango fruits of the same developmental stage E-2.
Biological activity of fungicides. Ten fungicides were initially tested on spore germination and mycelium growth assays, and five were selected for fruit protection assays; the data from these experiments are presented as general treatment means in Tab.4. The variation coefficient for the spore germination experiment was moderate to high (37.6%), suggesting that factors related to fungicide solubility in water or conditions for optimal contact or uptake by the fungal spores were not adequately controlled in the preparation of the spore germination assays. Dose effects of three fungicides (copper hydroxide, kasugamycin, validamycin) on colony growth were statistically significant, while two fungicides (azoxystrobin and thiabendazole) had a significant dose effect on spore germination.
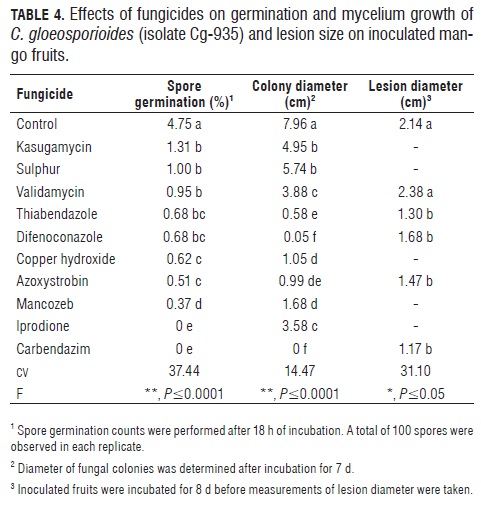
Eight fungicides reduced (P≤0.0001) spore germination by 80-100%, with two of them (carbendazim and iprodione) causing total inhibition (Tab.4). Mycelium growth of the pathogen was significantly inhibited by all tested fungicides (P≤0.0001), but the strongest inhibition presented in a culture medium amended with carbendazim or difenoconazole. Based on these results, five fungicides (azoxystrobin, carbendazim, thiabendazole, difenoconazole and validamycin) were used in fruit protection assays where significant (P≤0.05) but moderate effects were observed for three fungicides (carbendazim, azoxystrobin and thiabendazole), with lesion size reductions around 50% in relation to the unprotected control.
Biological activity of plant extracts. Tab.5 shows the effect of six plant extracts on germination, colony growth and fruit infection by the C. gloeosporioides isolate Cg-935. Most plant extracts tested did not produce a strong effect on the pathogen. Among the plant extracts, Desfan and Ecoswing had significant dose effects on mycelium growth of the pathogen, with higher effects at the highest dose as detected in the colony growth tests. However, this effect was not evident on spore germination or fruit infection assays. Therefore, data from plant extract experiments are presented as general treatment means of plant extracts.
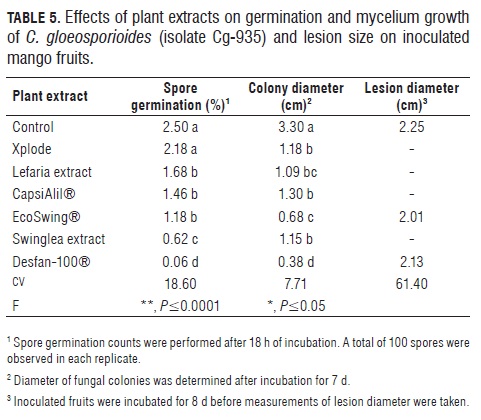
Reduction of spore germination was statistically significant (P≤0.0001) with one treatment (Desfan), inducing 98% inhibition of germination; this plant extract also caused a significant (P≤0.05) reduction (88%) in colony growth. When applied on the mango fruit surface however, none of the extracts significantly reduced lesion size (Tab.5).
Discussion
The effects of C. gloeosporioides inoculum on mango include: immediate lesion development following infection of the flowers, establishment of quiescent infections on immature fruits, or causation of anthracnose lesions following new infections of physiologically mature fruits (Arauz, 2000). Since heavy losses result from fruit anthracnose, we focused on features of the pathogen important for the onset of infection and development of fruit symptoms. Various studies describing the lifecycles of anthracnose pathogens (Bailey et al., 1992) suggest that spore germination needs to be blocked in order to prevent quiescent infections and reduce fruit disease. Other studies (Benito et al., 2000; Osorio et al., 2006; Moral et al., 2009) indicate that flower infection as well as quiescent infections on foliage could be important sources of inoculum for fruit infection.
Protecting mango fruits from anthracnose development is a common practice which relies mostly On fungicide spray programs or physical treatment following harvest (Dodd, 1991; Chowdhury et al., 2007); consequently, finding alternative treatments is desirable but little progress has been made. Fruit anthracnose development in mango may be influenced by a complex array of conditions that determine the effects of the inoculum and the infection of host tissues (Wharton and Diéguez-Uribeondo, 2004). The results from this study are preliminary and represent only a small set of conditions related to the development of this disease; promising treatments identified in this study should be tested for effectiveness on anthracnose control under a wider set of conditions and a greater number of pathogen isolates, representing the diversity and complexity of the C. gloeosporioides population associated with mango.
Separate experiments with microbial filtrates, fungicides and plant extracts were conducted, whereby spore germination was assessed after 18 h of incubation. Spore germination is affected by factors like age, temperature and light or chemical composition of the medium (Abril et al., 2008); which could explain the variation in germination levels observed among the experiments. In this study, overall spore germination levels were low, suggesting that the fungus may have unidentified internal constraints or regulatory mechanisms for survival similar to spore dormancy, as described for several plant pathogens (De Wolf and Isard, 2007). Alternatively, C. gloeosporioides spores may lose viability very rapidly after removal from their source, but data were not taken to support this hypothesis. Spore germination of C. gloeosporioides was strongly inhibited by microbial culture filtrates, fungicides and two plant extracts, suggesting that some of these treatments may hold promise for disease control. These should be tested more thoroughly, particularly microbial isolates since they also caused a highly significant inhibition of mycelium growth of the pathogen originating from the mango tissue surfaces and therefore, could have the ability to adapt and colonize host tissues. In addition, more experiments with fungicides and plant extracts are necessary to account for factors related to product solubility in water and optimize exposure or uptake by the fungal spores.
Colony growth of C. gloeosporioides on culture medium was stable across the experiments and appears to be a reliable trait to study sensitivity of the pathogen to various products. Mycelium growth in natural settings could be targeted for control at two specific stages: after spore germination but before penetration or during activation of quiescent infections on the fruit surface. Postharvest treatments for anthracnose control focus on this later stage of the interaction (Arauz, 2000; Arias and Carrizales, 2007). All microbial filtrates, most fungicides and two plant extracts (Tabs. 3, 4, 5) caused a strong inhibition in mycelium growth of the isolates Cg-917 and Cg-935 of the pathogen; therefore, it would be of interest to evaluate some of these products for their ability to specifically inactivate quiescent infections of C. gloeosporioides on mango fruits. Mycelium growth, and its associated enzymatic activity (Huckelhoven, 2007), may be responsible for flower damage by C. gloeosporioides in mango; therefore, flower protection by some of the products tested in this study is worth trying, particularly with microorganisms which may be able to colonize developing reproductive structures.
The lesion size of E-2 mango fruits inoculated with sporulating colonies of the isolates Cg-917 or Cg-935 showed high variation coefficients (31-61%) across the experiments; some intrinsic factors of the used plant material or differences in the inoculum load could explain this variability in the results. Mango fruits at the E-2 growth stage were used for inoculations based on previous studies; however there is some degree of uncertainty about the age of fruits of similar size and shape; this could explain why the reaction to the inoculation was not uniform. Results from this study indicated only small effects of fungicides or plant extracts on anthracnose development in inoculated fruits, while microbial filtrates showed significant reduction in lesion size. This is very promising since current control practices rely on fungicides or costly physical treatments (Kumar et al., 2007; Sundravadana et al., 2007).
Anthracnose development from late infections of mature fruits and from quiescent infections of immature fruits may proceed at different rates. These differences may be due to: a) the metabolic stage of the inoculum, b) the amount of active propagules at the infection site on the fruit surface and c) the time taken for initial colonization or lesion progress (Moral et al., 2009). All these factors have to be taken into consideration in future studies aiming to test potential control products with detached mango fruits, since natural disease development takes place not only from recent infection events but also from quiescent infections occurring in early stages of fruit growth. Furthermore, how much spore load migrates to the developing mango fruit and from what sources is a question that needs to be answered in order to understand some of the conditions for disease onset on the fruit and design or adjust control practices based on chemical or biological products.
Conclusions
Spore germination and mycelium growth of C. gloeosporioides can be inhibited or reduced in vitro by fungicides, plant extracts and microbial liquid culture filtrates; these types of experiments can be used to identify substances with high biological activity against the pathogen.
Development of anthracnose lesions on inoculated mango fruits was strongly reduced by filtrates from the liquid culture of various microorganisms, suggesting that some antagonists may be potential biocontrol agents of C. gloeosporioides and should be tested further in the field and for postharvest conditions.
Acknowledgements
The authors thank Mr. Jorge Arguelles for helping with the data analysis, Mr. Jaime Aguirre for his work in the fungicide experiments and Mr. José Dimas Segura for collecting field samples for microbial isolation.
Literature cited
Abril, M., K.J. Curry, B.J. Smith, and D.E. Wedge. 2008. Improved microassays used to test natural product-based and conventional fungicides on plant pathogenic fungi. Plant Dis. 92, 106-112. [ Links ]
Afanador, L., D. Minz, M. Maymon, and S. Freeman. 2003. Characterization of Colletotrichum isolates from tamarillo, passiflora, and mango in Colombia and identification of a unique species from the genus. Phytopathology 93, 579-587. [ Links ]
Arauz, L.F. 2000. Mango anthracnose: economic impact and current options for integrated management. Plant Dis. 84, 600-611. [ Links ]
Arias, B. and L. Carrizales. 2007. Control químico de la antracnosis del mango (Mangifera indica L.) en pre y postcosecha en el municipio de Cedeño, estado Monagas, Venezuela. Bioagro 19(1), 1-8. [ Links ]
Bailey, J.A., R.J. O'Connell, R.J. Pring, and C. Nash. 1992. Infection strategies of Colletotrichum species. pp. 88-120.In: Bailey, J.A. and M.J. Jeger (eds.). Colletotrichum: Biology, Pathology and control. CAB, Wallingford, UK. [ Links ]
Barnett, J.A., R.W. Payne, and D. Yarrow. 2000. Yeasts: characteristics and Identification, 3rd ed. Cambridge University Press, Cambridge, UK. [ Links ]
Benítez, T., A. Rincón, M. Limón, and A.C. Codón. 2004. Biocontrol mechanisms of Trichoderma strains. Intl. Microbiol. 7, 249-260. [ Links ]
Benito, E.P., M. Arranz, and A. Eslava. 2000. Factores de patogenicidad de Botrytis cinerea. Rev. Iberoam. Micol. 17, s43-s46. [ Links ]
Beno-Moualem, D. and D. Prusky. 2000. Early events during quiescent infection development by C. gloeosporioides in unripe avocado fruits. Phytopathology 90(5), 553-559. [ Links ]
Cadavid, M., S. Kelemu, and J.A. Osorio. 2007. Genetic diversity in the anthracnose pathogen infecting tropical fruits in Colombia. Phytopathology 97(7), S-16. [ Links ]
Castro, A., J. Romero, J.A. Osorio, E. Martínez, and J. Hio. 2007. Distribución temporal y espacial de fuentes de inoculo de Colletotrichum acutatum en el follaje sano de tomate de árbol. Fitopatol. Colomb. 31(1)1-5. [ Links ]
Cerón, L., M. Higuera, M. Sánchez, S. Bustamante, and G. Buitrago. 2005. Crecimiento y desarrollo de Colletrotrichum gloesporioides f. alatae durante su cultivo en medios liquidos. Acta Biol. Colomb. 11(1), 99-109. [ Links ]
Cheng, W., H. Ishii, and K. Nishimura. 2006. Fungicide sensitivity and phylogenetic relationships of anthracnose fungi isolated from various fruits in Japan. Plant Dis. 90, 506-512. [ Links ]
Chowdhury, M.; M. Rahim, K. Khalequzzaman, M. Humauan, and M. Alam. 2007. Effect of plant extracts and time of application on incidence of anthracnose, yield and quality of mango. Intl. J. Sustain. Crop Prod. 2(5), 59-68. [ Links ]
De Wolf, E.D. and S.A. Isard. 2007. Disease cycle approach to plant disease prediction. Annu. Rev. Phytopathol. 45, 203-220. [ Links ]
Dodd, J.C. 1991. Pre and postharvest control of mango anthracnose in the Philippines. Plant Pathol. 40, 576-583. [ Links ]
Donadio, S., P. Monciardini, R. Alduina, and P. Mazza. 2002. Microbial technologies for the discovery of novel bioactive metabolites. J. Biotechnol. 99, 187-198. [ Links ]
Huckelhoven, R. 2007. Cell wall-associated mechanisms of disease resistance and susceptibility. Annu. Rev. Phytopathol. 45, 101-127. [ Links ]
Korsten, L. 2006. Advances in control of postharvest diseases in tropical fresh produce. Intl. J. Postharv. Technol. Innov. 1, 48-61. [ Links ]
Korsten, L., E. De Villiers, R.C. Wehner, and J.M. Kotzet. 1997. Field spray of Bacillus subtilis and fungicides for control of pre-harvet fruit disease of avocado in South Africa. Plant Dis. 81, 455. [ Links ]
Kumar, A., N. Reddy, K. Reddy, and M. Devi. 2007. Evaluation of fungicidal resistance among Colletotrichum gloeosporioides isolates causing mango anthracnose in Agri Export Zone of Andhra Pradesh, India. Plant Pathol. Bull. 16, 157-160. [ Links ]
Liyanage, H., M. Millan, and H. Kistler. 1992. Two genetically distinct populations of Colletotrichum gloeosporioides from citrus. Phytopathol. 82(11), 1371-1376. [ Links ]
MADR, Misterio de Agricultura y Desarrollo Rural. 2009. Anuario estadístico de frutales y hortalizas 2004-2008 y sus calendarios de siembras y cosechas. Bogota. [ Links ]
Martínez, E., J. Hio, J.A. Osorio, and M.F. Torres. 2009. Identification of Colletotrichum species causing anthracnose on Tahiti lime, tree tomato and mango. Agronomía Colombiana 27 (2): 211-218. [ Links ]
Moral, J., R.Oliveira, and A. Trapero. 2009. Elucidation of the disease cycle of olive anthracnose caused by Colletotrichum acutatum. Phytopathology 99, 548-556. [ Links ]
Osorio, J.A., E. Viáfara, and N. Murcia. 2000. Aspectos epidemiológicos y de manejo de la antracnosis de los cítricos. pp. 41-48. In: Memorias del Seminario Nacional sobre Enfermedades de los Cítricos. Pereira, Colombia. [ Links ]
Osorio, J.A., M.F. Torres, J.C. Hio. 2006. Detección y análisis de la distribución de infecciones latentes de Colletotrichum acutatum J. H. Simmonds en árboles de frutales tropicales. Fitopatol. Colomb. 29(2), 45-52. [ Links ]
Ospina, C.A. and J.A. Osorio. 2005. Caracterización de poblaciones del género Colletotrichum causante de la antracnosis de los cítricos. Fitopatol. Colomb. 29(1), 15-22. [ Links ]
Páez-Redondo, A. 1995. Uso de variedades tolerantes: alternativa para el Manejo de antracnosis (Colletotrichum gloeosporioides) en mango (Mangifera indica). AS COLFI-Informa 21, 36-39. [ Links ]
Paterson, R. and P.D. Bridge. 1994. Biochemical tecniques for filamentous fungi. IMI Technical Handbooks No. 1. CAB, Wallingford, UK. [ Links ]
Reyes, A., J.A. Osorio, E. Martínez, and J. Hio. 2007. Estudio de la relación fenológica reproductiva y el comportamiento de las infecciones latentes de Colletotrichum acutatum en tomate de árbol. Fitopatol. Colomb. 31(2), [ Links ] 37-42.
Rojas-Martínez, R., E. Zavaleta, D. Nieto, and M. Acosta. 2008. Virulence and genetic variation of Colletotrichum gloeosporioides. (Penz.) Penz. and Sacc. on mango (Mangifera indica L.) cv. Haden. Rev. Mex. Fitopatol. 26, 21-26. [ Links ]
Schaad, N.W. 2001 Laboratory guide for identification of plant pathogenic bacteria. Departament of Plant Pathology, University of Georgia, St. Paul, MN . [ Links ]
Stockwell, V. and J. Stack. 2007. Using Pseudomonas spp for integrated bio-control. Phytopathology 97, 244-249. [ Links ]
Sundravadana, S., D. Alice, S. Kuttalam, and R. Samiyappan. 2006. Control of mango anthracnose by azoxystrobin. Tunis. J. Plant Protection 1, 109-114. [ Links ]
Sundravadana, S., D. Alice, S. Kuttalam, and R. Samiyappan. 2007. Efficacy of azoxystrobin on Colletotrichum gloeosporiodes Penz growth and on controlling Mango anthracnose. J. Agr. Biol. Sci. 2(3), 10-15. [ Links ]
Wharton, P.S. and J. Diéguez-Uribeondo. 2004. The biology of Colletotrichum acutatum. Anales del Jardín Botánico de Madrid 61, 3-22. [ Links ]
