










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.30 no.2 Bogotá May/Aug. 2012
POSTHARVEST PHYSIOLOGY & TECHNOLOGY
Content of the cyanogenic glucoside amygdalin in almond seeds related to the bitterness genotype
Contenido del glucósido cianogénico amigdalina en semillas de almendra con relación al genotipo con sabor amargo
Guillermo Arrázola1, Raquel Sánchez P.2, Federico Dicenta2, and Nuria Grané3
1Food Engineering Program. Faculty of Agricultural Sciences. Universidad de Córdoba. Monteria (Colombia). guillermo.arrazola@ua.es2Department of Plant Breeding, Centre of Edafology and Applied Biology of the Segura-Consejo Superior de Investigaciones Científicas (CEBAS -CSI C). Murcia (Spain).
3Departamento of Analytical Chemistry, Nutrition and Food Science, Faculty of Sciences, University of Alicante. Alicante (Spain). Received for publication: 29 August, 2010. Accepted for publication: 29 June, 2012.
ABSTRACT
Almond kernels can be sweet, slightly bitter or bitter. Bitterness in almond (Prunus dulcis Mill.) and other Prunus species is related to the content of the cyanogenic diglucoside amygdalin. When an almond containing amygdalin is chopped, glucose, benzaldehyde (bitter flavor) and hydrogen cyanide (which is toxic) are released. This two-year-study with 29 different almond cultivars for bitterness was carried out in order to relate the concentration of amygdalin in the kernel with the phenotype (sweet, slightly bitter or bitter) and the genotype (homozygous: sweet or bitter or heterozygous: sweet or slightly bitter) with an easy analytical test. Results showed that there was a clear difference in the amount of amygdalin between bitter and non-bitter cultivars. However, the content of amygdalin did not differentiate the other genotypes, since similar amounts of amygdalin can be found in the two different genotypes with the same phenotype.
Key words: kernel taste, benzaldehyde, hydrogen cyanide, Prunus.
RESUMEN
Las semillas de almendras pueden ser dulces, ligeramente amargas y amargas. El amargor en almendro (Prunus dulcis Mill.) y en otras especies de Prunus se relaciona con el contenido de la amígdalina diglucósido cianogénico. Cuando una almendra que contiene amigdalina se tritura, produce glucosa, benzaldehído (sabor amargo) y ácido cianihídrico (que es tóxico). El estudio es realizado durante dos años, con 29 variedades de almendra diferentes para la amargura o amargor, se ha realizado con el fin de relacionar la concentración de la amígdalina en el núcleo con el fenotipo (dulce, ligeramente amargo y amargo) y el genotipo (homocigota: dulce o amargo o heterocigótico: dulce o amarga un poco) por un ensayo de análisis fácil. Los resultados mostraron que existía una clara diferencia en la cantidad de amigdalina entre amargo / cultivares no amargo. Sin embargo, el contenido de amigdalina no diferenciaba entre los otros genotipos, ya que cantidades similares de amigdalina se puede encontrar en los dos genotipos diferentes con el mismo fenotipo.
Palabras clave: sabor amargo, benzaldehído, ácido cianhídrico,
Introduction
Prunus kernels accumulate cyanogenic diglucoside amygdalin, which is related to bitterness (McCarty et al., 1952; Conn, 1980; Frehner et al., 1990; Møller and Seigler, 1991; Swain et al., 1992; Poulton and Li, 1994; Dicenta et al., 2002; Franks et al., 2008; Sánchez-Pérez et al., 2008). Upon disruption of tissue containing amygdalin this will be degraded by b-glucosidases with release of glucose, benzaldehyde and hydrogen cyanide (Morant et al., 2008). This two-component system, each of them chemically inert, will provide the plant with a chemical defense against herbivores, insects and pathogens (Conn, 1969; Nahrstedt, 1985; Jones, 1998; Morant et al., 2003; Zagrobelny et al., 2004; Nielsen et al., 2006; Zagrobelny et al., 2007a; Zagrobelny et al., 2007b), as a source of energy reserves. In the almond (Prunus dulcis Mill.), bitterness is controlled by a single gene, Sk (Sweet kernel gene), which is dominant over the bitter gene (Heppner, 1923; Heppner, 1926; Grasselly and L'amandier, 1972; Kester et al., 1977; Gharbi, 1981; Vargas and Romero, 1988; Dicenta and García, 1993; Dicenta et al., 2007). The Sk gene has been localized in linkage group five in different genetic linkage maps, but its function is still unknown. As a result of the development of genetic linkage maps, some molecular markers have been found close to the Sk locus (Joobeur et al., 1998; Bliss et al., 2002; Sánchez-Pérez et al., 2007). However, a marker allele associated with the bitterness allele remains to be identified. Recently, amygdalin and, its precursor, prunasin (cyanogenic monoglucoside) content, and the enzymes involved in the amygdalin pathway, have been studied in four different almond genotypes (Sánchez-Pérez et al., 2008). Distinct cellular localizations of the enzymes involved in the degradation pathway, possibly involving a seed coat prunasin hydrolase, have also been suggested to be related to bitterness in the almond (Sánchez-Pérez et al., 2008). Furthermore, an enzyme involved in the biosynthetic pathway has also been related to bitterness in the almond (Franks et al., 2008).
On the other hand, most of the cultivated almond cultivars are heterozygous for this trait, mainly sweet, although some of them are slightly bitter. When two heterozygous cultivars are crossed in breeding programs, 25% of the offspring will be bitter and therefore eliminated from the breeding process, since the industry is mainly interested in sweet-kernelled cultivars. This disadvantage reduces the efficiency of breeding programs, and it is especially frequent when a self-compatible progenitor is used since most of the self-compatible cultivars are heterozygous for bitterness (Grasselly and Crossa-Raynaud, 1980; Dicenta, 1991). This is indeed happening in the American Breeding Program, since the bitter P. webbi has been used as a selfcompatible progenitor. Consequently, 50% of the offspring will be bitter when the other progenitor used in the cross is heterozygous, such as Carmel or Livingstone (Ledbetter and Pyntea, 2000; Gradziel et al., 2001). Fortunately, a study of these descendants has enabled the breeders to classify the cultivars used as progenitors as homozygous or heterozygous (Grasselly and Crossa-Raynaud, 1980; Dicenta, 1991; Vargas et al., 2001), which is a very arduous (3-4 years), and therefore expensive, task. The main objective of this research is to study the variability of the amygdalin content in a representative set of almond cultivars, its relation with the phenotype (sweet, bitter or slightly bitter) and the genotype (sweet homozygous, bitter homozygous or heterozygous), in order to identify the bitterness genotype of an almond cultivar with an easy analytical test.
Material and methods
Plant material
The content of amygdalin was analyzed for two years in 23 cultivars and in six descendants of the cross Garrigues × Tuono, provided by the Almond Breeding Program of the Centre of Edafology and Applied Biology of the Segura- Consejo Superior de Investigaciones Científicas (CEBAS - CSI C) CEBAS -CSI C, Murcia (Spain).
The phenotype and genotype of some of these cultivars were previously determined by different authors (Grasselly and Crossa-Raynaud, 1980; Dicenta, 1991; Vargas et al., 2001):
- Sk/Sk (sweet): Del Cid, Ferragnès, Peraleja, Primorskii, Ramillete, Titan. Sk/Sk (sweet): Marcona, Desmayo Largueta, Atocha. Sk/Sk (slightly bitter): Garrigues, Genco, Tuono. Sk/Sk (bitter): S3060, S3062, S3076, S3108, S3112, S3126. Sk/Sk or Sk/Sk (genotype unknown, but sweet): Achaak, Bonita, Carretas, CEBAS , Colorada, Ferraduel, La Mona, Pajarera, Planeta, Rumbeta, and Tioga.
Fifty mature almond fruits were collected per cultivar and the hull and shell removed. Kernels were frozen, lyophilized in a Cryodos Telstar lyophilizer for 48h at -85°C at 1 Pa, ground, homogenized, and analyzed. Two independent analyses per sample were performed.
Amygdalin content
Chromatographic determination of amygdalin levels was carried out for 0.2 g of kernel samples and extracted with 10 mL of methanol for 12 h at room temperature. Kernel extractions were done with 0.1 g of polyvinylpolypirrolidone (Sigma) or active carbon (Norit CNR 115) to ensure the freedom from pigments, which can interfere with the chromatography. Determination was performed isocratically in a waters high-performance liquid chromatography, Controler model 600 (HPLC) system, following a procedure similar already described (Kajiwara et al., 1983), under the following conditions: Waters Symmetry column 250 x 4.6 mm, flow rate 1.5 mL min-1, acetonitrile:water 20:80 as eluent, 20 μL of sample, and detection under UV at 218 nm.
Results, expressed as mg amygdalin/100 g of dry weight (DW), were tested using analysis of variance and Tukey´s test to determine differences between years and cultivars.
Results and discussion
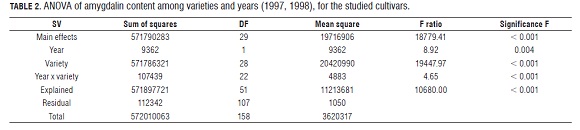
Analysis of variance of the amygdalin content showed significant differences between cultivars and between years as well as the year x cultivar interaction (Tab.1).
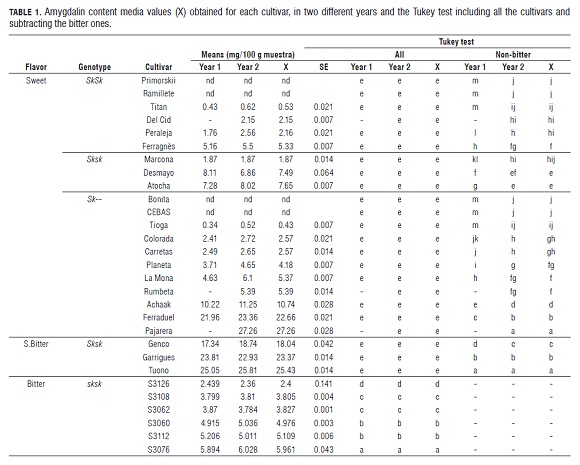
For the majority of the cultivars (Ramillete, Bonita, Marcona etc.), the content of amygdalin was similar in both years ( Tab.2). However, in the first year the content of some cultivars was slightly lower (Achaak, Colorada, Carretas, Ferraduel, S3060, S3076, etc.), or slightly higher (Desmayo Largueta, Garrigues, S3126, S3062, and S3112).
In general, there was a high variability of the amygdalin content between the studied cultivars. The average content of the bitter cultivars was much higher than the non-bitter ones ( Tab.2).
Tukey's test with all the cultivars showed clear differences between bitter and non-bitter (sweet and slightly bitter) cultivars, but did not between or within sweet and slightly bitter ones. Within the bitter cultivars, four different groups were identified.
In general, the kernel taste of cultivars is related to amygdalin content, although with some variability. In sweet cultivars such as Primorskii, Ramillete, Bonita and CEBAS , amygdalin was not detected, but in others such as Ferraduel or Pajarera it was until 27 mg/100 g DW. In slightly bitter cultivars such as Garrigues or Tuono, amygdalin content was until 25 mg/100 g DW. In bitter cultivars, the detected levels were between 2400 mg/100 g DW (S3126) and almost 6000 mg/100 g DW (S3076).
The number of plant species with cyanogenic glucosides is more than 2650 (Seigler and Brinker, 1993; Bak et al., 2006). Studies on passion fruit seeds (Passiflorae) obtained 3.1 mg/100 g DW (Chassagne et al., 1996), in lime 7000 mg/100 g DW of linamarin, and in bamboo 500 mg/100 g DW of taxiphyllin (Shahidi and Wanasundara, 1997
Furthermore, in linen seeds, linamarin (300 mg/100 g DW) and linustatin (6000 mg/100 g DW) were detected.
Amygdalin has also been detected in other fruit tree kernels such as plum (4,100 mg/100 g), apricot (2,394 mg/100 g), cherry (2,306 mg/100 g) and apple (739 mg/100 g) (Lucas and Sotelo, 1983). In the case of apricot, different concentrations of amygdalin have been detected: 4,400 and 6,500 mg/100 g DW (Femenia et al., 1995), or 5,500 and 7,000 mg/100 g DW (Stoewsand et al., 1975; Briggs and Yuen, 1978; Mandenius et al., 1983; Stosic et al., 1987; Voldrich et al., 1989; Gómez et al., 1998; Negri et al., 2008).
In bitter almond kernels, amygdalin levels were 5,808 mg/100 g DW (Corradi and Micheli, 1982; Shahidi and Wanasundara, 1997). Another analysis determined 926 mg of amygdalin/100 g DW for the bitter variety Sassari 11 and 106 mg/100 g DW in the sweet variety Arrubia, but in the sweet variety Texas, amygdalin was not detected (Usai and D'hallewin, 1992). Amygdalin content in sweet cultivars and slightly bitter ones from Sicily and Apulia was higher than the American or Russian ones, except for the American variety Perlees (Barbera et al., 1987). In fact, the content detected by these authors was much higher than the content detected in this study for cultivars such as Genco (390 versus 18 mg/100 g) and Tuono (750 versus 25 mg/100 g). Recently, 9 µmol/100 mg fresh weight (FW) of amygdalin (4,082 mg amygdalin/100 g FW) was detected in the bitter genotype S3067 (Sánchez-Pérez et al., 2008).
In general, the differences in amygdalin levels found in bitter cultivars of different Prunus species are within the range determined in this study (2,400-6,000 mg/100 g), with the exception of the results already mentioned (Barbera et al., 1987; Shahidi and Wanasundara, 1997).
In order to detect some differences within the non-bitter group, we analyzed the data without the bitter cultivars, since their amygdalin content was much higher. Analysis of variance only with non-bitter cultivars showed significant differences between cultivars and between years, the same as when all the genotypes were included ( Tab.3).
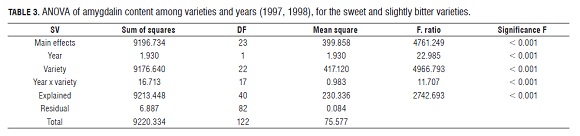
The Tukey's test did not differentiate sweet and slightly bitter cultivars (Tab.2). In general, sweet cultivars had less amygdalin than the slightly bitter ones, although some sweet cultivars (Ferraduel and Pajarera) had similar or higher amygdalin content than some slightly bitter ones. Tukey's test neither distinguised between homozygous and heterozygous sweet cultivars. In general, sweet homozygous cultivars (Sk/Sk) contained lower levels of amygdalin than the sweet heterozygous ones, but Marcona (Sk/Sk) had a lower content than Del Cid, Peraleja and Ferragnès (Sk/Sk). Despite the lack of differences between Sk/Sk and Sk/Sk sweet cultivars, we tried to classify the cultivars of an unknown genotype into these groups. So, Bonita, CEBAS and Tioga may be Sk/Sk and Achaak, Ferraduel and Pajarera Sk/Sk. The rest of the sweet cultivars could not be classified based on the amygdalin content.
It has been shown that the heterozygous slightly bitter cultivars, like Garrigues, tend to generate more slightly bitter descendants than other heterozygous sweet cultivars (Dicenta and García, 1993). Therefore, the slightly bitter cultivars may pass to the offspring the capacity to produce the slightly bitter taste, apart from the level of amygdalin.
Conclusions
We can conclude that there is a high variability in amygdalin content among the studied cultivars, being this content characteristic for each variety and quite stable year by year.
The bitter cultivars showed amygdalin contents much higher than the non-bitter ones; and slightly bitter ones had higher contents than sweet ones, but with some exceptions.
Finally, the amount of amygdalin is not a useful tool to identify Sk/Sk and Sk/sk cultivars. Therefore, further studies will be focused on the development of molecular markers for screening populations of fruit trees such as the almond with an easy PCR-based method.
Literature cited
Bak, S., S.M. Paquette, M. Morant, A.V. Rasmussen, S. Saito, N. Bjarnholt, M. Zagrobelny, K. Jørgensen, S. Osmani, T. Hamann, H.T. Simonsen, R. Sánchez-Pérez, T.B. Van-Heeswijck, B. Jørgensen, and B.L. Møller. 2006. Cyanogenic glycosides, a case study for evolution and application of cytochromes P450. Phytochem. Rev. 5(2-3), 309-329. [ Links ]
Barbera, G., L. Di Marco, G. Fatta Del Bosco, and P. Inglese. 1987. Behaviour of 26 almond cultivars growing under rainfed and semiarid conditions in Sicily. pp. 17-32. In: Colloque du Grempa. Reus, Spain. [ Links ]
Bliss, F.A., S. Arulsekar, M.R. Foolad, A.M. Becerra, A. Gillen, M.L. Warburton, A.M. Dandekar, G.M. Kocsisne, and K.K. Mydin. 2002. An expanded genetic linkage map of Prunus based on an interspecific cross between almond and peach. Genome 45(3), 520-529. [ Links ]
Briggs, D.R. and D. Yuen. 1978. Determination of cyanide in apricot kernels. Proc. Nutr. Soc. Austral. 3(1), 103-104. [ Links ]
Chassagne, D., J.C. Crouzet, L.C. Bayonove, and R.L. Baumes. 1996. Identification and quantification of passion fruit cyanogenic glycosides. J. Agric. Food Chem. 44(12), 3817-3820. [ Links ]
Conn, E.E. 1969. Cyanogenic glycosides. J. Agric. Food Chem. 17(3), 519-526. [ Links ]
Conn, E. E. 1980. Cyanogenic compounds. Annu. Rev. Plant Physiol. 31(1), 433-451. [ Links ]
Corradi, C. and G. Micheli. 1982. Total hydrocyanic acid content of macaroons. Industry Alimentary 21(1), 459-466. [ Links ]
Dicenta, F. 1991. Mejora genética del almendro (Prunus dulcis Miller) por cruzamientos intervarietales: herencia de caracteres y selección. Ph.D. thesis. Universidad de Murcia, Murcia, Spain. [ Links ]
Dicenta, F. and J.E. García. 1993. Inheritance of the kernel flavour in almond. Heredity 70(1), 308-312. [ Links ]
Dicenta, F., P. Martínez-Gómez, N. Grané, M.L. Martín, A. León, J.A. Cánovas, and V. Berenguer. 2002. Relationship between cyanogenic compounds in kernels, leaves, and roots of sweet and bitter kernelled almonds. J. Agric. Food Chem. 50(7), 2149-2152. [ Links ]
Dicenta, F., E. Ortega, and P. Martínez-Gómez. 2007. Use of recessive homozygous genotypes to assess the genetic control of kernel bitterness in almond. Euphytica 153(1-2), 221-225. [ Links ]
Femenia, A., C. Rosello, A. Mulet, and J. Canellas. 1995. Chemical composition of bitter and sweet apricot kernel. J. Agric. Food Chem. 43(2), 356-361. [ Links ]
Franks, T.K., A. Yadollahi, M.G. Wirthensohn, J.R. Guerin, B.N. Kaiser, M. Sedgley, and C.M. Ford. 2008. A seed coat cyanohydrin glucosyltranserase is associated with bitterness in almond (Prunus dulcis) kernels. Funct. Plant Biol. 35(1), 236-246. [ Links ]
Frehner, M., M. Scalet, and E.E. Conn. 1990. Pattern of the cyanide potential in developing fruits. Plant Physiol. 94(1), 28-34. [ Links ]
Gharbi, A. 1981. Résultats préliminaires des croisements intervariétaux d'amandier réalisés en Tunisie. Options Meditérr. 81(1), 23-35. [ Links ]
Gómez, E., L. Burgos, C. Soriano, and J. Marín. 1998. Amygdalin content in the seeds of several apricot cultivars. J. Sci. Food Agric. 77(2), 84-186. [ Links ]
Gradziel, T.M., P. Martínez-Gómez, F. Dicenta, and D.E. Kester. 2001. The utilization of related Prunus species for almond variety improvement. Amer. Pomol. Soc. J. 55(2), 100-108. [ Links ]
Grasselly, C. and P. L'amandier. 1972. Caractères morphologiques et physiologiques des variétés, madalité de leurs transmissions chez les hybrides de première génération. Ph.D. thesis. University of Bordeaux, Bordeaux, France. [ Links ]
Grasselly, Ch. and P. Crossa-Raynaud. 1980. L'Amandier. Maisonneuve et Larose, Paris. [ Links ]
Heppner, J. 1923. The factor for bitterness in the sweet almond. Genetics 8(4), 390-392. [ Links ]
Heppner, J. 1926. Further evidence on the factor for bitterness in the sweet almond. Genetics 11(6), 605-606. [ Links ]
Jones, D.A. 1998. Why are so many food plants cyanogenic. Phytochemistry 47(2), 115-162. [ Links ]
Joobeur, T., M.A. Viruel, M.C. De Vicente, B. Jáuregui, J. Ballester, M.T. Dettori, I. Verde, M.J. Truco, R. Messeguer, I. Battle, R. Quarta, E. Dirlewanger, and P. Arús. 1998. Construction of a saturated linkage map for Prunus using an almond x peach F2 progeny. Theor. Appl. Genet. 97(2), 1034-1041. [ Links ]
Kajiwara, N., C. Tomiyama, T. Ninomiya, and Y. Hosogai. 1983. Determination of amygdalin in apricot kernel and processed apricot by high performance chromatography. J. Food Hyg. Soc. Jpn. 24(1), 42-46. [ Links ]
Kester, D.E., P.E. Hansche, W. Beres, and R.N. Asay. 1977. Variance components and heritability of nut and kernel traits in almond. J. Amer. Soc. Hort. Sci. 102(3), 64-266. [ Links ]
Ledbetter, C.A. and M.A. Pyntea. 2000. The significance of kernel bitterness in the context of breeding almond for self-compatibility. J. Genet. Breed. 54, 221-225. [ Links ]
Lucas, B. and A. Sotelo. 1983. A simplified test for the quantitation of cyanogenic glucoside in wild and cultivated seeds. Nutr. Rpt. Intl. 29(3), 711-719. [ Links ]
Mandenius, C.F., L. Buelov, and B. Danielsson. 1983. Determination of amygdalin and cyanide in industrial food samples using enzymic methods. Acta Chem. Scand. 37(8), 739-742. [ Links ]
McCarty, C.D., J.W. Leslie, and H.B. Frost. 1952. Bitterness of kernels of almond x peach hybrids and their parents. Amer. Soc. Hort. Sci. 59(1), 254-258. [ Links ]
Møller, B.L. and D.S. Seigler. 1991. Biosynthesis of cyanogenic glycosides, cyanolipids and related compounds. pp. 563-609. In: Singh, B.K. (ed.). Plant amino acids, biochemistry and biotechnology, Marcel Dekker, New York, NY . [ Links ]
Morant, M., S. Bak, B.L. Møller, and D. Werck-Reichhart. 2003. Plant cytochromes P450: tools for pharmacology, plant protection and phytoremediation. Curr. Opin. Biotechnol. 14(2), 151-162. [ Links ]
Morant, A.V., K. Jørgensen, C. Jørgensen, S.M. Paquette, R. Sánchez- Pérez, B.L. Møller, and S. Bak. 2008. b-Glucosidases as detonators of plant chemical defense. Phytochemistry 69(9), 1795-1813. [ Links ]
Nahrstedt, A. 1985. Cyanogenic compounds as protecting agents for organisms. Plant Syst. Evol. 150(1-2), 35-47. [ Links ]
Negri, P., D. Bassi, E. Magnanini, M. Rizzo, and F. Bartolozzi. 2008. Bitterness inheritance in apricot (P. armeniaca L) seeds. Tree Genet. Genomes 4(4), 767-776. [ Links ]
Nielsen, K.A., M. Hrmova, J.N. Nielsen, K. Forslund, S. Ebert, C.E. Olsen, G.B. Fincher, and B.L. Møller. 2006. Reconstitution of cyanogenesis in barley (Hordeum vulgare L.) and its implications for resistance against the barley powdery mildew fungus. Planta 223(5), 1010-1023. [ Links ]
Poulton, J.E. and C.P. Li. 1994. Tissue level compartmentation of (R)-amygdalin and amygdalin hydrolase prevents large-scale cyanogenesis in undamaged Prunus seeds. Plant Physiol. 104(1), 29-35. [ Links ]
Sánchez-Pérez, R., W. Howad, F. Dicenta, P. Arús, and P. Martínez- Gómez. 2007. Mapping major genes and quantitative trait loci controlling agronomic traits in almond. Plant Breed. 126(3), 310-318. [ Links ]
Sánchez-Pérez, R., K. Jørgensen, C.E. Olsen, F. Dicenta, and B.L. Møller. 2008. Bitterness in almonds. Plant Physiol. 146(3), 1040-1052. [ Links ]
Seigler, D.S. and A.M. Brinker. 1993. Characterization of cyanogenic glycosides, cyanolipids, nitroglycosides, organic nitro compounds and nitrile glucosides from plants. pp 51-93. In: Dey, P.M. and J.B. Harborne (eds.). Methods in plant biochemistry, alkaloids and sulfur compounds. Academic Press, London. [ Links ]
Shahidi, F. and P.K. Wanasundara. 1997. Cyanogenic glycosides of flaxseeds. pp. 171-185. In: Shahidi, F. (ed.). Antinutrients and phytochemicals in food. ACS Symposium Serie 622, 171-185. [ Links ]
Stoewsand, G.S., J.L. Anderson, and R.C. Lamb. 1975. Cyanide content of apricot kernels. J. Food Sci. 40(5), 1107-1120. [ Links ]
Stosic, D., M. Gorunovic, and B. Popovich. 1987. Preliminary toxicological study of the kernel and oil of some species of the genus Prunus. Plantees Medicales et Phytotherapie 21(1), 8-13. [ Links ]
Swain, E., C.P. Li, and J.E. Poulton. 1992. Tissue and subcellular localization of enzymes catabolizing (R)-amygdalin in mature Prunus serotina seeds. Plant Physiol. 100(1), 291-300. [ Links ]
Usai, M. and G. D'hallewin. 1992. Cyanogenic glucosides contained in different organs of bitter and sweet almond. pp. 233-236. In: EUR 14081 Amélioration génétique de deux espèces de fruits secs méditerranéens: L'amandier et le pistachier. Proceedings of the VIII Grempa Meeting. Nîmes, France. [ Links ]
Vargas, F.J. and M.A. Romero. 1988. Comparación entre descendencias intervarietales de almendro en relación con la época de floración y la calidad del fruto. Repport EUR 11557, 59-72. [ Links ]
Vargas, F.J., M.A. Romero, and I. Batlle. 2001. Kernel taste inheritance in almond. Options Méditerr. 56, 129-134. [ Links ]
Voldrich, M., H. Opatova, V. Kyzling, and J. Hrdlicka. 1989. Determination of hydrogen cyanide and cyanogenic glycosides in fresh and preserved fruits. Czech J. Food Sci. 7(4), 271-278. [ Links ]
Zagrobelny, M., S. Bak, A.V. Rasmussen, B. Jørgensen, C.M. Naumann, and B.L. Møller. 2004. Cyanogenic glucosides and plantinsect interactions. Phytochemistry 65(3), 293-306. [ Links ]
Zagrobelny, M., S. Bak, C.T. Ekstrøm, C.E. Olsen, and B.L. Møller. 2007a. The cyanogenic glucoside composition of Zygaena filipendulae (Lepidoptera: Zygaenidae) as effected by feeding on wild-type and transgenic lotus populations with variable cyanogenic glucoside profiles. Insect Biochem. Mol. Biol. 37(1), 10-18. [ Links ]
Zagrobelny, M., S. Bak, C.E. Olsen, and B.L. Møller. 2007b. Intimate roles for cyanogenic glucosides in the life cycle of Zygaena filipendulae (Lepidotera, Zygaenidae). Insect Biochem. Mol. Biol. 37(11), 1189-1197 [ Links ]
