










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.30 no.3 Bogotá Sept./Dec. 2012
CROP PHISIOLOGY
Effects of environmental factors on the morphometric characteristics of cultivated lettuce (Lactuca sativa L.)
Efecto de variables ambientales sobre caracteres morfométricos en el cultivo de lechuga (Lactuca sativa L.)
Antonio D. Ojeda1, Gustavo A. Ligarreto2 and Orlando Martínez3
1Department of Sciences Biology, Faculty of Sciences, Universidad de Los Andes. Bogota (Colombia). ojeda853@gmail.com2Department of Agronomy, Faculty of Agronomy, Universidad Nacional de Colombia. Bogota (Colombia).
3Private consultant in Biostatistical Analysis. Bogota (Colombia).
Received for publication: 21 July, 2011. Accepted for publication: 30 October, 2012.
ABSTRACT
We evaluated the phenotypic response of three varieties of lettuce Batavia Colwart, Lisa Elisa and Crespa Verde Cassabella in three growing environments: plastic greenhouse, glass greenhouse and open field on the Bogota Plateau (Chia, Cundinamarca), in order to generate useful information to improve the quality of plants grown by farmers, ensuring a healthy product with characteristics that meet market demands. The three agroecosystems were constantly monitored for the environmental variables of temperature, relative humidity and solar radiation; which showed a significant variation between the three agroecosystems. To assess the phenotypic response, we examined the effect of the environmental variables mentioned on seven morphometric variables of agronomic interest. The study showed that, for the variables germination percentage and fresh weight, the environmental effect reached a variance percentage of 79 and 44%, respectively. On the contrary, it was established that the average phenotypic response of the variables clean weight and percentage of dry mass corresponds especially to the effect of genetic varieties with a variance percentage of 35 and 45%, respectively. Similarly, for the variables specific leaf area and total leaf area, it was established that both the effects of environmental and genetic variables explain 87% (50 genetic, 37 environmental) and 56% (31 genetic, 25 environmental), respectively, of the total variability observed.
Key words: phenotypic variance, genotype x environment, phenotypic plasticity.
RESUMEN
Se evaluó la respuesta fenotípica de tres variedades de lechuga Batavia Colwart, Lisa Elisa y Crespa Verde Cassabella en tres ambientes de cultivo: invernadero de plástico, invernadero de vidrio y campo abierto en la Sabana de Bogotá (Chía, Cundinamarca) con el objetivo de generar información útil para mejorar la calidad de las plantas obtenidas por los agricultores, asegurando un producto sano y con características para satisfacer las demandas del mercado. Los tres agroecosistemas fueron caracterizados por medio de un monitoreo constante de las variables ambientales de temperatura, radiación solar y humedad relativa, lo cual evidenció una importante variación entre los tres agroecosistemas. Para evaluar la respuesta fenotípica, se analizó el efecto de las variables ambientales mencionadas sobre siete variables morfométricas de interés agronómico. El estudio demostró que para las variables porcentaje de germinación y peso fresco el efecto ambiental alcanzo un porcentaje de varianza del 79 y 44%, respectivamente. Por el contrario, se estableció que la respuesta del fenotipo promedio de las variables peso limpio y porcentaje de masa seca corresponde especialmente al efecto genético de las variedades con un porcentaje de varianza del 35 y 45%, respectivamente. De igual manera, para las variables área foliar específica y área foliar total se estableció que tanto los efectos de las variables ambientales como los efectos genéticos de las variedades explican en su orden el 87% (genético 50, ambiental 37) y 56% (genético 31, ambiental 25) respectivamente, de la variabilidad total observada.
Palabras clave: varianza fenotípica, genotipo x ambiente, plasticidad fenotípica.
Introduction
Lettuce (Lactuca sativa L.) is an annual, autogamous plant that belongs to the Asteraceae family and is cultivated in Colombia mainly in the departments of Cundinamarca, Boyaca, Valle del Cauca, Norte de Santander and Antioquia. It is sown with a wide range of varieties according to the morphological characteristics, specifically the shape of the leaves and the head: Batavia, Iceberg, Boston, Romana and loose leaves. Recent genetic studies on this species have identified several specific sites of the genome that control the morphological characteristics of the leaves, flowers and seeds (Robinson et al., 1983; Waycott et al., 1999). However, for the genus Lactuca, little is known about the ability of the genotype to produce different phenotypes, depending on the environmental conditions of its location (Valladares et al., 2007).
Recent studies have shown that plants exhibit phenotypic plasticity in ecological traits ranging from their morphology, physiology and anatomy, to those related to development and reproduction (Valladares et al., 2007). Grube et al. (2003) found that the poor development of the hypocotyl and inflorescence stems in mutants of lettuce corresponds to poor gravitropic response that affects the quality of seedlings and is associated with the expression of the mutant gene SGR1/SCR in certain environments. Other important studies have shown that both radiation and temperature are environmental factors that determine suitable seed germination and quality of lettuce seedlings. Hiroshi (1964) found that solar radiation, and especially long-wave red light, is a limiting factor in the germination of lettuce seeds. Temperatures above 24°C cause significant changes in the endosperm layer, generating a mechanical force that counteracts and may even inhibit germination (Takeba and Matsubara, 1976; Sung et al., 1997). Takaki and Zaia (1984) showed that these two environmental factors could be correlated, as high temperatures also affect the phytochromes, which are photo-transmitters acting on blue light and red light, causing an expansion of leaves, reduced germination and problems with the photoperiod.
It is known that both metabolic pathways and development are influenced by the environment, due to the expression of many genes of little effect, i.e. quantitative phenotypic traits. Thus, the environmental impact can produce large changes in the morphology and ethology of individuals. However, most of these environmental effects tend to be subtle and cause amplifications or reductions in morphometry, the number of progeny, growth rates or physiological functions. For this reason, this type of study is still considered in the development of varieties of plants of economic importance with a range of geographic adaptability and response to different cultivation techniques, providing the opportunity to develop a wide range of cultivars that meet production and consumer demands, allowing the exploration of different marketing alternatives and the creation of stability in crops with different levels of yield (Lynch and Walsh, 1998).
Thus, a proper handling of knowledge and phenotypic plasticity in crops is of vital importance because it facilitates knowledge on the phenotypic response of crop varieties or groups to heterogeneous conditions occurring in time and space in agroecosystems. This is particularly important when dealing with plants like lettuce that have numerous varieties that are cultivated during all seasons and with a wide geographical distribution with different production potentials, as in the case of Colombia. The aim of the study was to assess the phenotypic response of three varieties of lettuce (Lactuca sativa L.) Colwart Batavia, Lisa Elisa and Crespa Verde Cassabella in three growing environments: plastic greenhouse, glass greenhouse and open field on the Bogota Plateau.
Materials and methods
The study was conducted over a growing season in the facilities of the Centro de Bio-Sistemas of the Universidad de Bogota Jorge Tadeo Lozano, located on the central highway in the north of the town of Chia, Cundinamarca. We used three growing environments: traditional plastic greenhouse, glass greenhouse and open field. The environments presented a differential grade in average temperature (day/night), relative humidity and solar radiation. We used three varieties of lettuce: Colwart Batavia, Lisa Elisa and Crespa Verde Casabella. For each environment, two sowing methods were used: direct seeding and transplantation using seedlings germinated in a nursery, thus six treatments were generated. The establishment of the experiment was performed in a randomized block design in each of the mentioned environments. The experimental unit formed 32 individuals per treatment, 10 replicates were used per treatment for glass greenhouse and open field, while for plastic greenhouse, there were six replications.
Sowing and seedlings
The cultivation of the plant varieties consisted of sowing half of the seeds in the growing environments of plastic greenhouse, glass greenhouse and open field. The other half was germinated for a month in the nursery. Afterwards, the seedlings showing two to four developed leaves were transplanted to the growing environments.
Each environment had an area of 200 m2, with a capacity for three plastic covered beds with the dimensions 25 x 2 m, with eight sowing rows and space for 640 individuals per bed for glass greenhouse and open field and four beds of 25 x 2 m with space for 288 individuals per bed for plastic greenhouse. In the nursery, plastic trays were used for germination of up to 200 individuals with a space of 1.5 cm3 each, to obtain 1,000 individuals per variety.
As a substrate, we used a mixture of 30% soil, 30% rice husk, 30% coconut fiber and 10% compost. The seedlings were placed individually in plastic bags with a capacity of 0.6. One to two daily irrigations were performed in the three growing environments. Within 60 d of the commencement of the cultivation, foliar application was performed for development and at 75 d one application of pesticide was carried out to control aphids.
Morphometric
The following variables were evaluated at the end of cultivation in eight individuals per experimental unit.
Fresh weight (g) by taking the aerial part of each plant and weighing it.
Clean weight (g) corresponded to the amount of biomass that was suitable for marketing, with aerial biomass removed, avoiding decaying leaves, with attacks from pests or diseases.
Useful mass index: ratio of clean weight over fresh weight.
Biomass (%): each plant was packed in a paper bag and dried in an oven at 72°C for 96 h, then the dry weight was measured and the value obtained with fresh weight was determined from the percentage of dry mass.
Total leaf area (cm2) was determined from digital photographs of leaves processed with ImageJ 1.40 program for Macintosh. Knowing the total weight of the leaves and the leaf area /leaf weight ratio made it possible to estimate the total leaf area of each individual.
Specific leaf area (cm2 g-1) corresponded to the amount of biomass contained in g cm-2 of leaf area, calculated from the total leaf area of an individual over total leaf weight.
Climate monitoring
For determining the relative humidity of each environment, a sensor comprised of a pair of thermocouples coupled to a data logger mark COX was used, one with a dry bulb and the other with a wet bulb. The sensors were suspended in PVC cylinders to ensure quality and representativeness of the data collection for 3.5 months of monitoring. To measure the total global solar radiation, diagonal transects were conducted with six sampling points with weather stations per environment.
Data analysis
Statistical analyzes were performed using SPSS 16.0 for Macintosh. Variance components were estimated by a linear model with effects of genotypes and fixed environments (Lynch and Walsh, 1998).
Results and discussion
The life cycle of the crop was different in each variety. The shortest was with Lisa Elisa, with 10 weeks from planting to harvest, followed by Crespa Verde Cassabella with about 12 weeks after sowing and variety Batavia Colwart with 15 weeks.
Environmental factors
The data showed that the entire study area has about 12 h of sunlight, from 6:00 AM until 5:50 PM. The intensity of the radiation varies significantly between different types of greenhouses; the glass greenhouse had the lowest average radiation with 126.56 W m-2 and maximum radiation with 301 W m-2, followed by the plastic greenhouse with an average radiation of 175.93 W m-2 and maximum radiation of 565 W m-2, and finally the open field with an average radiation of 359.13 W m-2 and maximum radiation of 1,008 W m-2. This is because of the light reflectance of surfaces, that is, the property of materials such as plastics and glass reflect light. For the glass greenhouse, it is important to note that during the day, the degree of light incidence changes with respect to the sun's position and angle of the various building panels forming the greenhouse glass, so the glass reflectance varies, behaving like a mirror during certain hours of the day. Additionally, glass, being a heavier material than plastic, needs a stronger support structure and thicker beams, creating a physical barrier to light.
With regard to temperature, all environments showed two sets of normally distributed data with different averages during the 12 h of light and 12 h of darkness which together presented bimodal dispersion for each environment. This allowed us to characterize each environment according to the average daytime temperature and average overnight temperature. The open field was shown to be the environment with the lowest average day/night temperature of 15/10°C and a maximum temperature of 22°C, followed by the plastic greenhouse (17/12°C, max. 30°C), the nursery (18/12°C, max. 25°C) and then the glass greenhouse (23/13°C, 35°C max.). The minimum temperature showed no significant differences between the environments and ranged around 5.3°C. This is because the glass, unlike plastic, has the property of being transparent to solar radiation, both visible and infrared; however, it is opaque to the far-end infrared radiation emitted by heated bodies (plants, the ground, etc.) and hence the glass greenhouse acts as a heat trap where temperature increases both during the day and the night.
For the amount of moisture retained by the environment, a proportional relationship with respect to temperature was found. This was expected since a temperature rise increases the water retention capacity of the air and consequently decreases the value of relative humidity (Fig.1). This pattern was shown for all environments, with the only difference being the amount of water retained by the air, where the open field proved to be the lowest moisture environment with an average of 4 g water per kg air and a maximum of 46 g of water per kg air, followed by the plastic greenhouse and finally the glass greenhouse. Like temperature, the minimum amount of moisture in the air did not show significant differences between the environments and ranged around 0.10 g water per kg air. This is because the high temperatures of the glass greenhouse produced humidity in the environment, mainly in the air, by increasing levels of water retained by the air.
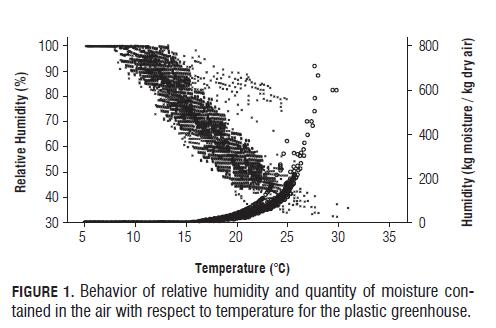
The glass greenhouse had a high temperature environment with very high peak levels (35°C) and maintained a significantly higher amount of moisture compared to the other environments. These characteristics made the glass greenhouse an environment with a low contribution to the quality of the lettuce due to decomposition of the lower leaves, growth of microorganisms such as Botrytis sp. and the passage from vegetative to floral with seed production due to high temperatures.
The plastic greenhouse had radiation levels and moderate temperatures without significant differences from the nursery, except the amount of moisture retained by the air, which was higher and more constant in the nursery. Meanwhile, the open field showed the typical characteristics of the Bogota Plateau with high solar radiation, low moisture retained in the air and lower average temperatures during the day and night with a low of 0°C. There was also a bigger effect from slug and aphid attack.
Phenotypic variables
The germination percentage was found to be lower in the glass greenhouse than in the other two environments, and no significant differences were found between the plastic greenhouse and open field (Fig.2). This is related to the high temperatures of the glass greenhouse (23/18°C, max. 35°C) and is consistent with the results of Sung et al. (1998) who concluded that temperatures above 24°C for prolonged periods of time affect or even inhibit germination in lettuce. Takaki and Zaia (1984) and Yazgan et al. (2008) found that high temperatures and humidity could decrease the range of action of the active form of phytochrome, which acts directly on the germination of lettuce seeds. Additionally, other studies showed that high temperatures for germination of lettuce are also related to changes in the endosperm that act as a mechanical force which opposes germination (Takeba and Matsubara, 1976; Sung et al., 1997).
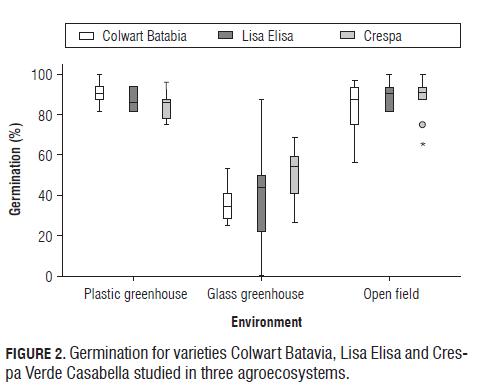
By analyzing the variance components, the results indicate that 79.19% of the variability observed in the germination corresponds to the effect of the environment, the expected result due to the wide effect of temperature and humidity on the phytochromes and endosperm (Tab.1). The effect due to the genetic component of the varieties was very low (0.05%), as well as the effect of the genotype x environment interaction (1.69%). The other 19% corresponds to the residual effects of the model (Tab.1).
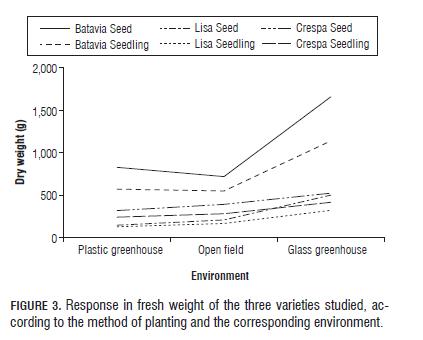
For fresh weight, an increased behavior was observed for the three varieties within the glass greenhouse which could be related to the high temperature thereof. All varieties showed a remarkable developmental trend from vegetative to floral with very elongated stems of 30-40 cm. However, the only variety which showed various branches of the central stem, flower formation and initial seed stages was Batavia. This could be related to the evolutionary history of the genus Lactuca, which comes from a wild ancestor of Europe and Asia (Lactuca scariola) found in temperate zones (Quintero et al., 2000; Gagliano, 2006).
Thus, being a plant of the genus Lactuca conditioned to climates warmer than the Bogota Plateau, it is expected that exposure to high temperatures and dry climates produces changes in the vegetative state and induces flowering with subsequent seed production as a result of favorable environmental conditions for reproduction and viable offspring (Pinzón, 1990). Results that agree with Osorio and Lobo (1983), who claim that, even though lettuce is an annual plant and blooms in the same cycle, high temperatures cause flowering. They also claim that the optimum average temperature for the normal development of the aerial part is 15 to 18°C with a maximum of 21°C depending on the variety.
Fig.3 shows a significant change for both treatments and seedling of three varieties in the glass greenhouse; this behavior is accentuated in the Batavia Colwart variety. It is noted that the seed and seedling treatments behave similarly in that they describe the same parallel linearity within the varieties. Seed treatments for the three varieties in the different environments showed a higher phenotype average than the seedling treatments. This could indicate that individuals that grow and develop in the same environmental conditions produce a better adaptation to the environment represented by the average value of the phenotype. As to the contribution of the variance to fresh weight, it was determined that most of the same corresponds to the environmental effect (43.76%), followed by genetic variety effect (14.49%) and finally the effect of the interaction (13.34%), which is relatively high (Tab.1).
For clean weight, the variance components indicate that the variety factor explains the highest percentage (34.97%) of the observed variance, followed by the environmental component (18.48%) and the interaction (3.56%) (Tab.1). This is because the clean weight assessment criterion depends largely on morphologic appearances of the variety such as shape, length and texture of the leaves, among others, which determine the level of plant material contamination with pathogens and soil that significantly affects its value as commercial material.
With regard to the above, the variable useful mass was only seen in the environments plastic greenhouse and open field because the lettuce obtained in the glass greenhouse was not useful due to its low quality for the market for the reasons stated above (Fig.4). However, a better performance of the variety Crespa Verde Cassabella was seen in the glass greenhouse over the open field, which together with the results of germination percentage suggests that this strain may have some level of thermo-tolerance. This coincides with studies of Sung et al. (1998), Gray (1975), Nascimento (2003) and Grazia et al. (2001), who evaluated several germinating lettuce genotypes that performed satisfactorily at higher temperatures. This result agrees with those reported by Osorio and Lobo (1983) who stated that flowering at high temperatures is more common in varieties of loose leaf and iceberg such as Lisa and Batavia respectively.
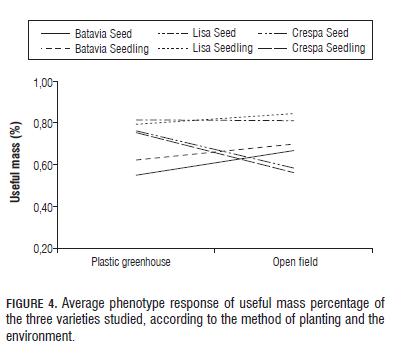
Fig.4 shows significant interaction for the varieties and planting methods. First, the variety Lisa Elisa is seen to be very stable between environments with respect to useful mass production, with no significant changes in the scale and range of the average phenotype. Regarding the variety Crespa Verde Cassabella, one sees that direct seeding gives a small improvement of adaptation (average phenotypic value) and that its performance is superior in the plastic greenhouse over open field, which suggests that it is a variety sensitive to the environmental conditions of the Bogota Plateau. By contrast, the variety Batavia Colwart showed better adaptation for treatments with seedlings, and showed better performance in open field as compared to plastic greenhouse, which coincides with the traditional way of growing this variety on the Bogota Plateau, and the low agronomic management requirements in the soil, irrigation and seedlings.
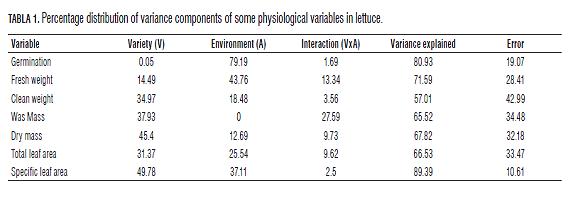
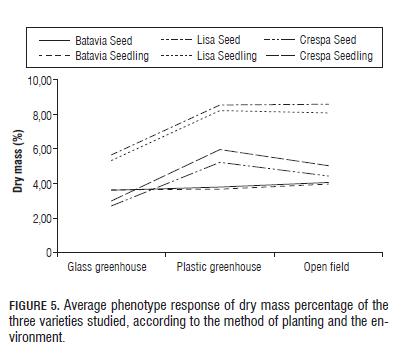
The results of dry mass (Fig.5) show that the most profitable variety in terms of production is the variety Lisa Elisa, which presented the highest values of average phenotype over the other varieties in all environments, which coincides with previous findings regarding the useful mass in the performance of Crespa Verde Cassabella in plastic greenhouse. However, in this case, the seedling treatment proved to be most profitable in terms of biomass production for this strain. Similarly, the results coincide for the variety Batavia Colwart, showing better performance in the open field, with little difference between the methods of planting. The variance components analysis indicates that the genetic component of the varieties explains 45.40% of the total variance observed, followed by the environmental component (12.69%) and a relatively high interaction component (9.73%) (Tab.1).
Leaf area of all plants in general is a plastic characteristic that responds to the mean conditions of solar radiation to produce shade for the lower parts or to improve the capture of light energy (Jarma et al., 2006). The results of total leaf area (Fig.6) showed a gradual increase in leaf area in low radiation environments (plastic and glass greenhouse) thereby confirming that the increase in area corresponds to an adjusting plant mechanism to low sunlight conditions to improve light energy uptake.
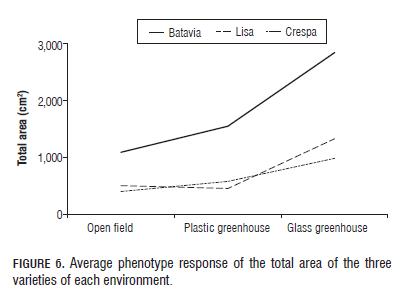
The analysis of variance components for this variable indicates that both the genetic component and the environmental component explained most of the variance observed with values of 31.37 and 25.54% respectively. An effect was observed from the interaction which explains a relatively high 9.62% of the total variance of the observed total leaf area.
Fig.7 shows that for specific leaf area there is a general increase in the amount of area per gram for all varieties in environments with less radiation (plastic and glass greenhouse). This is consistent with the results obtained for total area since several studies indicate that plants grown under shade or low solar radiation conditions tend to increase specific leaf area in order to increase the distribution of photosynthates at the expense of root growth or increase in leaf area, optimizing light energy capture (Páez et al., 2000). The variance components analysis shows that the genetic component of the varieties and the environmental component explained most of the variance observed with 49.78 and 37.11% followed by the interaction which explains a relatively low 2.50% of the total variance (Tab.1). This corresponds, as regards the ordinal distribution, with the results of total area and is important since certain authors (Hay and Porter, 2006) state that leaf area is also a characteristic that is set during the development of the plant, thus involving the expression of genes through metabolic pathways.
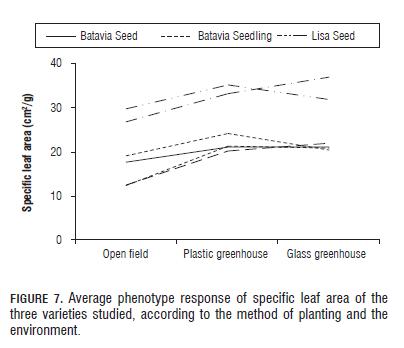
The lower specific leaf area in seed treatments corresponds to a rapid change from the vegetative to the floral stage as a result of the adaptive response to environmental conditions, and a change in energy use, from synthesis of leaf area to mainly synthesis of flowers and seeds. In the case of the variety Colwart Batavia, the difference is not noticeable because this was the only variety that presented an advanced floral state for both seed and seedling planting methods.
Conclusions
The environmental conditions significantly affected the production of the three varieties of lettuce tested and there was a differential behavior for quality for the environments. It was determined that temperature is a decisive factor in the germination of lettuce. Solar radiation causes significant responses in leaf area of adult individuals and thus affects quality. The humidity of the environment is associated with the amount of useful mass with humid environments with relatively high temperatures generating an environment conducive to the growth of microorganisms that affect quality.
The Lisa Elisa variety may be the most profitable in terms of useful mass production which proved very stable in open field and plastic greenhouse.
Acknowledgements
The authors thank the Centro de Bio-Sistemas of the Universidad de Bogota Jorge Tadeo Lozano for providing field instruments and laboratories to conduct research, as well as for the contribution of scientific and support staff that assisted with the development of this research.
Literature cited
Gagliano, E. 2006. Presentación de cultivo: Lechuga. Bol. Noved. Hortíc. 2, 1-2. [ Links ]
Gray, D. 1975. Effects of temperature on the germination and emergence of lettuce (Lactuca sativa) seeds. Ann. Appl. Biol. 86, 77-86. [ Links ]
Grazia, J., P.A. Tittonell, and Á. Chiesa. 2001. Efecto de la época de siembra, radiación y nutrición nitrogenada sobre el patrón de crecimiento y el rendimiento del cultivo de lechuga (Lactuca sativa L.). Invest. Agr. Prod. Prot. Veg. 16(3), 356-365. [ Links ]
Grube, R.C., E.B. Brennan, and E.J. Ryder. 2003. Characterization and genetic analysis of a lettuce (Lactuca sativa L.) mutant, weary, that exhibits reduced gravitropic response in hypocotyls and inflorescence stems. J. Exp. Bot. 54(385), 1259-1268. [ Links ]
Hay, R.K.M. and J.R. Porter. 2006. The Physyology of crop yield. Blackwell Publishing, Oxford, UK. pp. 35-47. [ Links ]
Hiroshi, I. 1964. The effects of temperature on the germination y radicle growth of photosensitive lettuce seed. Plant Cell Physiol. 5(4), 429-439. [ Links ]
Jarma, A., T. Rengifo, and H. Araméndiz-Tatis. 2006. Fisiología de estevia (Stevia rabaudiana) en función de la radiación en el Caribe Colombiano. II. Análisis de crecimiento. Agron. Colomb. 24(1), 38-47. [ Links ]
Lynch, M. and B. Walsh. 1998. Genetics and analisys of quantitative traits. Genotype x environmental interaction. Sinauer Associates, Sunderland, MA. [ Links ]
Osorio, J. and M. Lobo. 1983. Lechuga. pp. 515-526. In: Hortalizas. Technical Assistance Manual No. 28. Instituto Colombiano Agropecuario (ICA), Bogota. [ Links ]
Nascimento, W.M. 2003. Preventing Thermoinhibition in a thermosensitive lettuce genotype by seed inhibition at low temperature. Sci. Agríc. 60(3), 477-480. [ Links ]
Páez, A., V. Paz, and J.C. López. 2000. Crecimiento y respuesta fisiológicas de plantas de tomate cv. Río Grande en la época mayo-julio. Efecto del sombreado. Rev. Fac. Agron. (LUZ) 17, 173-184. [ Links ]
Pinzón, H. 1990. Producción de semillas de lechuga bajo invernadero (Lactuca sativa L.). M.Sc. thesis. Faculty of Agronomy, Universidad Nacional de Colombia. Bogota. [ Links ]
Quintero, I., J. Zambrano, M. Cabrita, and R. Gil. 2000. Evaluación en campo y postcosecha de nueve cultivares de lechuga Lactuca Sativa L. Rev. Fac. Agron. (LUZ) 17, 482-491. [ Links ]
Robinson, R.W., J.D. Mccreight, and E.J. Ryder. 1983. The genes of lettuce and closely related species. Pl. Breed. Rev. 1, 267-293. [ Links ]
Sung, Y., D.J. Cantliffe, and R.T. Nagata. 1997. Structural changes in lettuce seed during germination altered by genotype, seed maturation temperature and priming. HortScience 32, 526. [ Links ]
Sung, Y., D.J. Cantliffe, and R.T. Nagata. 1998. Seed developmental temperature regulation of thermotolerance in lettuce. Soc. Hort. Sci. 123(4), 700-705. [ Links ]
Takaki, M. and V.M. Zaia. 1984. Effect of light and temperature on the germination of lettuce seeds. Planta 160(2), 190-192. [ Links ]
Takeba, G.O. and S. Matsubara. 1976. Analysis of temperature effect on the germination of New York lettuce seeds. Plant and Cell Physiol. 17(1), 91-101. [ Links ]
Valladares, F., E. Gianoli, and J.M. Gómez. 2007. Ecological limits to plant phenotypic plasticity. New Phytologist. 176, 749-763. [ Links ]
Waycott, W., S.B. Fort, E.J. Ryder, and R.W. Michelmore. 1999. Mapping morphological genes relative to molecular markets in letuuce (Lactuca sativa L.). Heredity 82, 245-251. [ Links ]
Yazgan, S., S. Ayas, C. Demirtas, H. Büyükcangaz, and B.N. Candogan. 2008. Déficit irrigation effects on lettuce (Lactuca sativa var. Olenka) yield in unheated greenhouse condition. J. Food Agric. Environ. 6(2), 357-361. [ Links ]
