



![Stomatal behavior in fruits and leaves of the purple passion fruit (Passiflora edulis Sims) and fruits and cladodes of the yellow pitaya [Hylocereus megalanthus (K. Schum. ex Vaupel) Ralf Bauer]](/img/en/prev.gif)






Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.31 no.1 Bogotá Jan./Apr. 2013
CROP PHYSIOLOGY
Cadmium and chromium effects on seed germination and root elongation in lettuce, spinach and Swiss chard
Efecto del cadmio y cromo sobre la germinación y elongación radical de lechuga, espinaca y acelga
Oscar V. Bautista1, Gerhard Fischer1 and Julián F. Cárdenas2
1Department of Agronomy, Faculty of Agronomy, Universidad Nacional de Colombia. Bogota (Colombia). ovbautistac@unal.edu.co2Master Program in Agrarian Sciences, Faculty of Agronomy, Universidad Nacional de Colombia. Bogota (Colombia).
Received for publication: 3 January, 2013. Accepted for publication: 29 March, 2013.
ABSTRACT
The La Ramada district and the Bogota River are the principal water resources used for horticultural crop production on the Bogota Plateau, which contain channel pollutant materials, including heavy metals due to domestic and industrial activities on the Plateau. These materials have effects on crop production in this zone. The present research, under laboratory conditions, aimed to evaluate the effect of three concentrations (25, 35 and 50 µM L-1) of cadmium (Cd) and chromium (Cr) on imbibition, seed germination and root elongation in lettuce (Lactuca sativa var. Batavia), Swiss chard (Beta vulgaris var. cicla "White Ribbed"), and spinach (Spinacia oleracea Hib. 424), three species widely cultivated on the Plateau. The three species used for evaluation showed a differential susceptibility response to Cd and Cr. In lettuce, fresh weight increase (imbibition) was lower with all Cd concentrations at the last day of observation and at 25 µM L-1 of Cd in Swiss chard. Cadmium reduced seed germination by up to 46%, 97% and 8% in Swiss chard, lettuce and spinach, respectively. Also, root elongation decreased in Cd treatments by up to 57%, 89% and 56%, for Swiss chard, lettuce and spinach, respectively. Chromium, which showed fewer negative effects, decreased germination by up to 29% in Swiss chard, 6% in lettuce and 34% in spinach, as compared to the control.
Key words: heavy metals, water contamination, imbibition, protrusion.
RESUMEN
La producción hortícola en la Sabana de Bogotá tiene como principales fuentes hídricas al distrito de riego La Ramada y al rio Bogotá, el cual lleva en su cauce materiales contaminantes como metales pesados, provenientes de las diferentes actividades industriales y domésticas de la Sabana. Estos contaminantes afectan la producción de cultivos en esta zona. En el presente estudio, bajo condiciones de laboratorio, se evaluó el efecto del cadmio (Cd) y cromo (Cr) en tres concentraciones (25, 35 y 50 µM L-1), sobre la imbibición, germinación y elongación radical de lechuga (Lactuca sativa var. Batavia), acelga (Beta vulgaris var. cicla "White Ribbed") y espinaca (Spinacia oleracea híbrida 424), tres especies que son ampliamente cultivadas en la zona. Se encontró una susceptibilidad diferente a los dos compuestos metálicos en cada especie. En lechuga, en el último día de la observación, las tres soluciones de Cd disminuyeron el aumento del peso fresco de las semillas (imbibición), mientras este ocurrió en acelga solamente a 25 µM L-1 de Cd. Cadmio afectó la germinación en 46%, 97% y 8% en acelga, lechuga y espinaca, respectivamente; también, este metal redujo la elongación radical hasta un 57%, 89% y 56% en las tres especies, respectivamente. El Cr no mostró efectos tan negativos, pero redujo la germinación de acelga (29%), lechuga (6%) y espinaca (34%), comparado al control.
Palabras clave: metales pesados, contaminación de agua, imbibición, protrusión.
Introduction
Lettuce (Lactuca sativa, Asteraceae), Swiss chard (Beta vulgaris, var. cicla, Amarantaceae), and spinach (Spinacia oleracea, Amarantaceae) have 78%, 95%, and 81%, respectively, of their domestic production in the department of Cundinamarca (Ministry of Agriculture and Rural Development, 2009). On the Bogota Plateau, horticultural production is one of the many economic activities that take place in the region, and is carried out in small- and medium-scale farms, which supply the local market as well as providing a significant amount of production for international markets (Miranda et al., 2008).
It is estimated that the La Ramada irrigation district, located in the middle basin of the Bogota River, provides about 6.6% of the total water demand for irrigation of the Plateau while 93.4% comes from derivations on the banks of the river (SIA C, 2002). The Bogota River, along its course, drains the water of 6,000 km2, where about 8 million inhabitants live. In this basin, about 26% of the country′s economic activity is generated. The industrial type wastewater discharges release grease, oil and heavy metals such as cadmium (Cd), chromium (Cr), copper (Cu), lead (Pb), nickel (Ni) and other hazardous wastes (Conpes, 2004) into the river water. The sources of the Cd contaminants are predominantly restricted to industries of textiles, packaging materials, chemicals, fertilizers, paints, cements, machinery, electronics and automotive (Ideam and Dama, 2003). Traditional and continuous use of Bogota river water as a source of irrigation for crops is considered a source of contamination of soil and plants (Gonzalez and Mejia, 1995).
Heavy metals are defined as elements having a density greater than 5 g cm-3, such as copper, nickel, iron, zinc, manganese, molybdenum which are considered essential, and cobalt as beneficial to plants (Marschner, 2012); for their growth, senescence and energy processes. However, when the concentration of heavy metals exceeds the specified threshold, the levels lead to the malfunction of enzymes, oxidative stress, replacement of essential nutrients and other toxic effects on plants. Within the group of heavy metals, Cd is particularly toxic to the majority of plants and animals even at low concentrations (Shukla et al., 2007).
It has been shown that the effect of trace metals on imbibition, germination and root length may be linked to the ability of compounds to penetrate the seed coat and affect physiological processes associated with germination, growth and development (Seregin and Kozhevnikova, 2005). According to Kranner and Colville (2011), germination is affected in two ways by metals; the first is due to its toxicity and the second by the effect on water uptake during imbibition.
The objective of this study was to determine the effect of the heavy metals Cd and Cr on the germination of seeds of lettuce, Swiss chard and spinach in regards to imbibition, germination percentage and radical elongation, in order to have a reference of early stage adaptability to the conditions of irrigation with contaminated water.
Materials and methods
Seeds of lettuce Lactuca sativa var. Batavia, Swiss chard Beta vulgaris var. cicla "White Ribbed" and spinach Spinacia oleracea hybrid 424 were obtained from Impulsemillas (Bogota).
Treatments and variables
Treatments consisted of imbibition of 50 seeds of each species in 2 mL of the solutions presented in Tab. 1.
All seeds were treated with solutions of Cd and Cr and a control (distilled water) for a period of 12 d for lettuce and Swiss chard, and in the case of spinach, 15 d. Seeds were placed in Petri dishes of 7 cm in diameter. Germination took place on a filter paper disc No. 3 in a growth chamber (Versatile environmental test chamber, Sanyo, San Diego, CA). Temperatures in the chamber were, for lettuce and spinach, 20°C and for spinach, 15°C, while relative humidity was 70%.
Heavy metal solutions were replaced every third day to keep the concentration of the solutions constant and, also, to provide appropriate moisture for imbibition and germination of the seeds.
The imbibition curve of the seeds was determined for each treatment through registration of seed fresh mass at intervals of 24 h for 3 d for lettuce and 5 d for Swiss chard and spinach.
The protrusion (visible emergence of the radicle through the seed coat) and germination (radicle size = 5 mm length) of the seeds were measured daily for the first 7 d, thereafter, they were recorded at 9, 12 and 15 d. The germinated seeds were removed from the Petri dishes, leaving only the seeds that were still in the process of germination.
The mean and variation of germination time of a population of seeds are useful parameters to determine the response of the population to environmental influences. For this reason, we calculated the percentage of protrusion (PP ) and percentage of final germination (PG), the mean protrusion time (MPT) and mean germination time (MGT) and the mean velocity of protrusion (MVP) and mean velocity of germination (MVG), in accordance with Ranal and Santana (2006) and Anjum and Bajwa (2005), as follows:
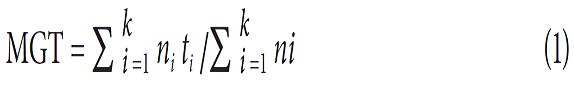

The percentage of protrusion (PP ) and the percentage of germination (PG) were taken as the sum of protruding or germinated seeds over the number of seeds per Petri dish as a percentage (ecuations 3 and 4), as according to Akinci and Akinci (2010):


Experimental design
We used a completely randomized design with four replicates, applying the SAS statistical analysis software; an Anova was carried out and the treatment means were evaluated by the Tukey test (P=0.05) to determine differences between treatments for each of the variables assessed.
Results
Lettuce
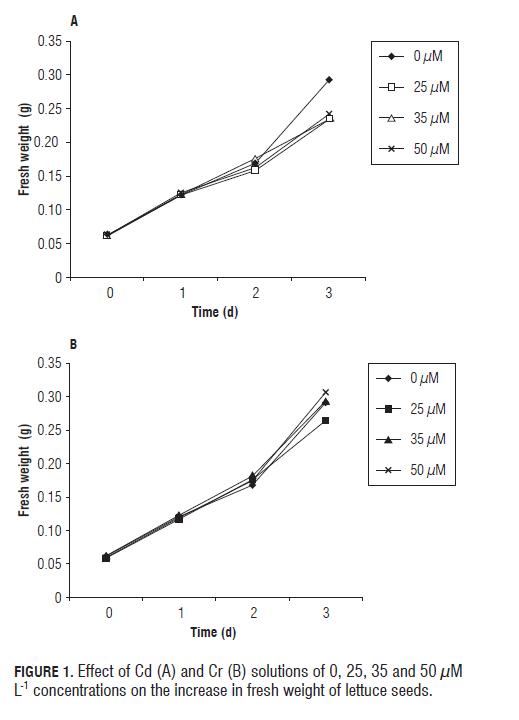
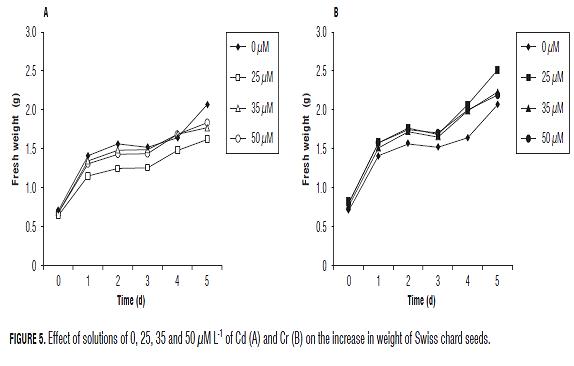
In both heavy metal solutions, the fresh weight (imbibition) of lettuce seeds increased with time of observation (Fig.1). At the third day of the treatments, all Cd solutions decreased seed fresh weight significantly (P=0.05), as compared to the control, while with Cr, only 25 µM L-1 tended to reduce imbibition of the seeds.
Concerning the effect of the two metals on lettuce protrusion percentage (PP ), the results showed no significant difference between the control and the treatments; the maximum difference was at the highest dose of Cr with a reduction of 5.5%, as compared to the control. The largest effect of Cd was evident at the highest concentration applied, with a decrease of 4.5%.
There was a slight tendency for the application of Cd and Cr to increase the number of protruding seeds per day and decrease protrusion time, but these variables did not show significant differences. The peak of mean velocity of protrusion (at 23.91 protruding seeds/day) was caused by the treatment of the highest Cd concentration, while the control treatment resulted in the lowest MVP (22.93). Moreover, the lowest MPT was seen with the medium concentration of Cd, with an average value of 2.16, and the highest MPT was obtained with the low Cd concentration.
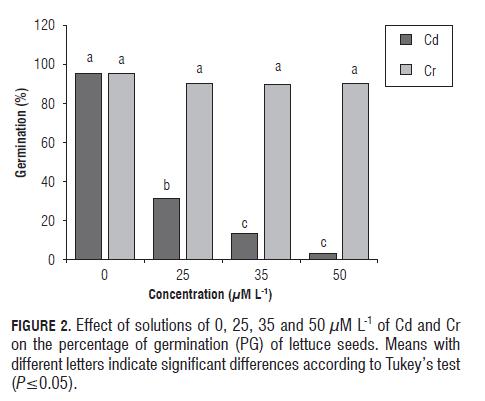
The results of germination percentage (PG) in lettuce (Fig.2) showed significant differences (P=0.05) between the treatments and the control at all levels of Cd, which caused a reduction of 64%, 82.5% and 92.5% for the concentrations of 25, 35 and 50 µM L-1, respectively. The maximum effect of Cr caused a decrease in the PG of only 5.5% with the medium concentration.
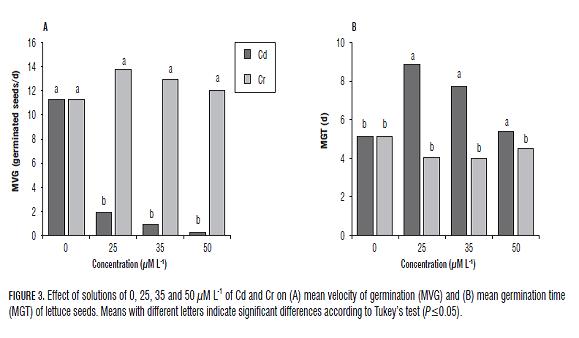
Cadmium significantly reduced MVG by up to between 9.3 and 11.1 seeds/day (P=0.05) (Fig.3a), while the Cr increased MVG by between 1 and 2 seeds/day. The MGT increased with the application of Cr for 3 d whereas MGT with Cd was lower than in the control until 1 d, showing a statistical difference for all concentrations applied (Fig.3b).
The root elongation of lettuce was affected differently by each metal. All Cd treatments interfered visibly and significantly (P=0.0001) with root elongation at the highest concentration (50 mM L-1). At 12 d of the experiment, the growth restriction was up to 2.7 cm, as compared to the control. Chromium did not cause adverse effects on this variable and even the concentration of 35 µM L-1 showed a stimulatory effect, which was significant when compared to the control. The effect of the two metals was constant and the differences were shown throughout the experiment (Fig.4).
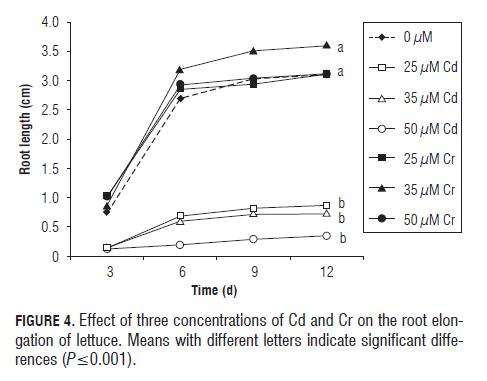
Swiss chard
The Swiss chard seed imbibition showed statistically significant differences in the 5th and last days of the test. The difference was recorded between the lowest concentrations of Cd vs. Cr (Fig.5), with a difference between them of 0.894 g in seed weight.
The maximum value of PP reached by Swiss chard seeds was 74% in the control, Cd caused the decline of this variable by 18%, 24% and 20% with the low, medium and high concentrations, respectively. Chromium caused a reduction of this variable by 1%, 10% and 18% with the low, medium and high concentrations, respectively (Fig.6). A significant difference from the control was observed only in the case of the treatment with 35 µM Cd L-1.

The MVP and MPT of Swiss chard seeds were altered most by the application of 35 mM L-1 of Cd (7.1 seeds/day and 5 d, respectively), while the most severe effect of Cr on these variables was caused by the concentration of 50 µM L-1 , which resulted in a MVP value of 8.1 seeds/day and 4.7 d for MPT.
The PG of Swiss chard seeds coincided with the behavior of PP , MVP and MGT, with the most severe effect of Cd at the dose of 35 µM L-1 (43, 5%), and for Cr at the highest dose, with a PG of 50%; despite the variation caused, neither treatment was significant.
The maximum root length of Swiss chard was achieved by the low concentration of Cr with a value of 3.7 cm, which was 0.05 cm above the control (Fig.7). The treatment of 50 µM L -1 reached a maximum value of 1.5 cm, causing a 2.1 cm reduction with respect to the control and the treatment of low Cr concentration. The treatments showed a highly significant difference (P=0.0001) which occurred from day 6 to 12.
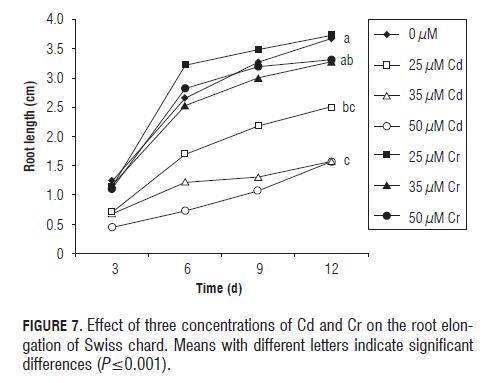
Spinach
The spinach seed imbibition, using solutions of Cd and Cr, had no significant effect on the increase in seed weight (Fig.8 ), nor was a differential effect from the metals or the different concentrations found. The protrusion of the spinach seeds revealed no significant difference between the treatments; however, the Cd application of 25 µM L-1 decreased PP by up to 19%. On the other hand, the effect of Cr on the inhibition of PP was greatest at 35 mM L-1 , reducing this variable by 4% and being less harmful when compared to Cd at the same concentration.
The MVP and MPT of spinach were affected similarly by Cd and Cr. The Low Cd concentration (25 µM L-1) had the greatest effect on the two variables, decreasing by 3.1 the number of protruding seeds each day and delaying the protrusion by 1.8 d. The other treatments showed a similar behavior to each other, but none of the effects were statistically significant.
The maximum PG observed in spinach was 88.5%, achieved by the treatment of 25 µM L-1 of Cr, which despite being 2.5% higher than the control seeds, was not statistically significant nor was the treatment of 25 mM L-1 of Cd which reduced the PG of spinach by 22%. The MGT and MVG of spinach showed a similar behavior to that described for protrusion of these variables, where the greatest effect was generated by the Cd treatment at a concentration of 25 µM L-1.
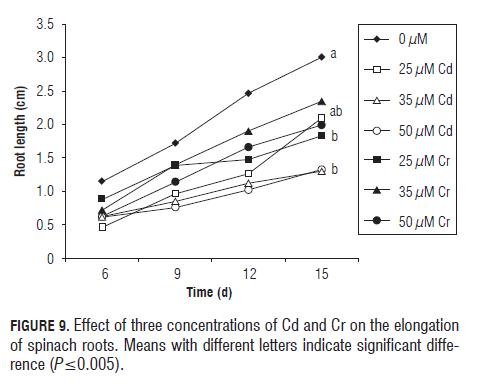
The root elongation of spinach Fig.9 was affected greatest at concentrations of 35 and 50 µM L-1 of Cd, with a growth reduction of 1.7 cm. Chromium also caused the reduction of spinach roots by 1.2 cm in comparison with the control, but significant differences only were recorded with the treatment of 25 mM L-1.
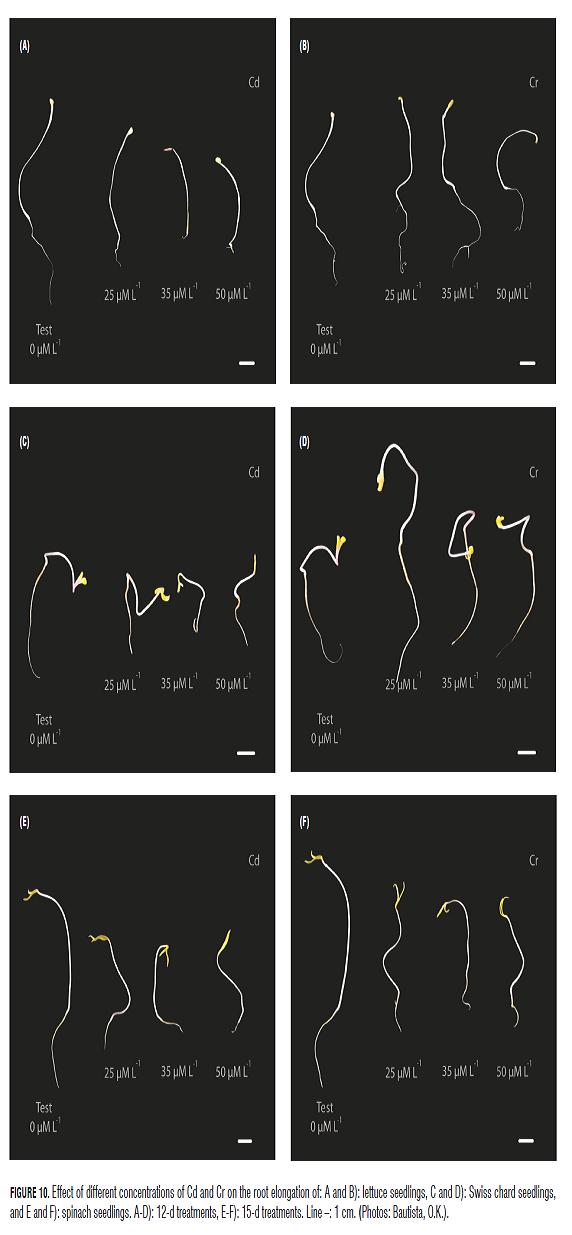
The graphic record of the experiment (Fig.10) shows the symptomatology of the treatments. In general, we can appreciate the crinkling caused by treatments of Cd and their intensity in inhibiting elongation. In the case of Cr necrosis, it is evident in some tissues of the radicle and also, a positive effect can be seen at low concentrations.
Discussion
The toxic effects of Cd had a greater impact than those of Cr. During germination, the lettuce showed a greater sensitivity to this heavy metal toxicity than Swiss chard and spinach. The results obtained in this investigation confirm the protective function of the living endosperm tissues that completely enclose the embryo (Sung et al., 2008; Kranner and Colville, 2011; Li et al., 2005), as it was observed that although the Swiss chard and spinach seeds were exposed to metal concentrations of up to 50 µM L-1, the germination percentages reached 46% and 85%, respectively. However, the subsequent process (root growth) was seriously affected and these effects were positively correlated with the concentration of the metal, which was observed more clearly in Cd treatments than with Cr.
Effects of Cd and Cr on seed imbibition
The water intake by the seeds was different between the species due to the distinct seed morphology of the species considered in this study and the protection that the seed coat gives against stressors, wherein, different degrees of permeability for different trace elements exist (Li et al., 2005). In this study, significant differences were observed between the treatments in the case of lettuce and Swiss chard, while spinach showed no significant difference, contrary to the increased sensitivity of seeds to stress after imbibition observed by Li et al. (2005).
The effect of Cd on the water intake of lettuce seed was significant on the 3rd day; all treatments of Cd caused an increased reduction of seed weight as compared to the control; these results may be due to the interference of Cd in water uptake by seeds, which has also been demonstrated in sorghum and pea seeds (Siddiqui et al., 2009). There exists evidence that cadmium permeates the cytosol through calcium channels in the plasmalemma resulting in changes of the cell-water relationship (Perfus-Barbeoch et al., 2002; Milone et al., 2003).
In our study, the Cd concentration effects on seed imbibition did not differ from each other. These results are contrary to Li et al. (2005) who used concentrations between 0.2 and 10 mM, finding a correlation between the concentration of the metal and its effect on water uptake by seeds.
The absence of Cr treatment effects on seed imbibition may be due to several factors, such as lower toxicity of Chromium in the Cr3+ state as compared to Cr6+ (Scoccianti et al., 2006). According to our results, it is not possible to state that the observed inhibition of PG is due to a deficient water uptake, since some treatments showed a certain inhibition in this variable but others did not. These results have to be further validated in future studies. Also, Mihoub et al. (2004) found no reduction in water uptake by Pisum sativum seeds; however, these authors observed that Cd interferes with the transport and the mobility of reserve nutrients present in the seeds.
Effects of Cd and Cr on protrusion
The metals tested caused no significant effects on the protrusion of lettuce and spinach seeds. Heavy metals inhibit cell division but not necessarily protrusion, which is carried out by this process and therefore might not be affected (Li et al., 2005). Furthermore, the permeability of seed coats can restrict the entry of heavy metals to the cells to avoid affecting the metabolism of the seed (Kozhevnikova and Seregin, 2005).
Effects of Cd and Cr on germination
Germination was one of the variables most affected by the application of Cd and Cr, in general, different concentrations showed distinct behaviors, which may be associated with differences in the permeability of the species seed coat.
The seeds appear to be designed to closely monitor environmental conditions; germination inhibition appears to be the first defense mechanism that a seed exhibits when environmental conditions are adverse (Li et al., 2005). The effect of metals on the previously explained protrusion have obvious implications for germination, there were seeds that showed successful protrusion but did not reach the stipulated radicle length (=5 mm) to be considered germinated, behavior that has been reported by Li et al. (2005) in Arabidopsis thaliana. This result may be due to toxic effects of metals that come into direct contact with the tissues of the developing seedling once protrusion occurs because prior to this, embryo tissues are not in direct contact with the metal due to the existing protection offered by the seed coat, which can prevent the concentration from entering the seed and, more specifically, the tissues of the embryo, and thereby reaching a level high enough to cause toxicity and affect PP (Kranner and Colville, 2011; Seregin and Kozhevnikova, 2005).
Cadmium decreased germination of all three species, but its effect was only significant in lettuce seeds. In general, accumulation of heavy metals differs between species; Ramos et al. (2002) demonstrated that the accumulation of heavy metals not only differs between species but also between the plant organs of the same species, with greater accumulation in tissues of roots than in other tissues.
Shaukat et al. (1999) reported Cd toxicity in the seeds of two species. Cadmium toxicity in the seed may be associated with its interaction with calmodulin, causing inhibition of the transport of reserve substances in the seed and/or interference with the hormonal status of the seed during germination (Raouhi et al., 2010).
Chromium affected germination of Swiss chard seeds to a greater degree. Rout et al. (2000) obtained similar results with 200 µM Cr, which reduced the PG of the weed species Echinochloa colona by 25%. Moreover, the spinach and Swiss chard PG were not as severely affected, which is consistent with results obtained by Akinci and Akinci (2010) who found no significant differences in the PG of Cucumis melo when applying a Cr concentration of 24 µM. The above mentioned observations suggest a difference in the level of susceptibility to Cr of the assessed species.
Effects of Cd and Cr on root elongation
We found that in addition to the effects that the two metals could cause on PG, the seeds that reached the state of germination were then subjected to stress caused by continuous exposure to applied metal salts, showing visible signs of darkening of the plumule and radicle and the stunting of the latter with all treatments of Cd and the higher concentrations of Cr.
The root elongation of the lettuce decreased significantly (P=0.0001) in all treatments and samplings of Cd with respect to the control, showing a relationship between the metal concentration and degree of growth inhibition. Root elongation was the variable most affected by the application of Cd in lettuce and Swiss chard. The results in the three species studied showed a similar behavior for root elongation, as seen in previous studies, for example Bowiea volubilis had an inhibited root size when treated with Cd at concentrations of 3 µM L-1 (Street et al., 2007), and the inhibition of wheat elongation appeared only at concentrations as low as 4.4 µM L-1 (Liu et al., 2008).
In lettuce, chromium, instead of inhibiting root growth, tended to stimulate it at 35 µM. Also, Jun et al. (2009), found that, in six species of legumes, root elongation of seedlings presented a higher sensitivity to Cr than that of germination. Lintschinger et al. (1997) reported that Cr can stimulate plant growth, which can be confirmed in our study for the case of lettuce (35 µM) and Swiss chard (25 µM) but not for spinach, which had a root growth that was reduced by Cr applications. Previous studies support this difference in behavior between species, e.g. Scoccianti et al. (2006) found that the application of CrCl3 in concentrations from 0.1 to 10 mM L-1 reduced the root growth of celery seedlings (Apium graveolens), while Pennisetum americanum and Parkisonia aculeate roots treated with 160 µM L-1 of Cr were not affected by this metal (Shaukat et al., 1999). Barceló et al. (1993) clearly demonstrated the beneficial influence of Cr on the chloroplast ultrastructure in Fe-deficient plants.
Conclusions
The obtained results show that the processes of germination and root elongation are affected differently in lettuce, Swiss chard and spinach; Cd being more toxic than Cr in the three species tested.
The germination of lettuce was more severely affected than the other two and the level of inhibition of germination percentage was related to the concentration of Cd. Root elongation was the variable most affected by the heavy metals applied, with lettuce being more susceptible to Cd than Swiss chard and spinach. The latter two responded similarly to the application of Cd. Spinach root elongation was more susceptible to Cr that those of Swiss chard and lettuce.
The results of this study, under laboratory conditions, suggest that germination processes will be severely affected when heavy metal-contaminated water is used.
Cited literature
Akinci, I. and S. Akinci. 2010. Effect of chromium toxicity on germination and early seedling growth in melon (Cucumis melo L.). Afr. J. Biotechnol. 9(29), 4589-4594. [ Links ]
Anjum, T. and R. Bajwa. 2005. Importance of germination indices in interpretation of allelochemical effects. Int. J. Agr. Biol. 7(3), 417-419. [ Links ]
Barceló, J., C. Poschenrieder, M.D. Vazquez, B. Gunse, and J.P. Vernet. 1993. Beneficial and toxic effects of chromium in plants: Solution culture, pot and field studies. Studies in Environmental Science No. 55. 5th International Conference on Environmental Contamination, Morges, Switzerland. [ Links ]
Conpes. 2004. Estrategia ambiental para el manejo del río Bogotá. Documento Conpes 3320. Consejo Nacional de Política Económica y Social. Departamento Nacional de Planeación. Ministerio de Ambiente, Vivienda y Desarrollo Territorial, Bogota. [ Links ]
González, S. and L. Mejía. 1995. Contaminación con cadmio y arsénico en suelos y hortalizas de un sector de la cuenca del río Bogotá. Suelos Ecuatoriales 25, 51-56. [ Links ]
Ideam and Dama. 2003. VI Fase de seguimiento de efluentes industriales y corrientes superficiales de Bogotá D.C. Instituto de Hidrología, Meteorología y Estudios Ambientales (Ideam) y Departamento Técnico Administrativo del Medio Ambiente (Dama), Bogota. [ Links ]
Jun, R., T. Ling, and Z. Guanghua. 2009. Effects of chromium on seed germination, root elongation and coleoptiles growth in six pulses. Int. J. Environ. Sci. Tech. 6(4), 571-578. [ Links ]
Kranner, I. and L. Colville. 2011. Metals and seeds: Biochemical and molecular implications and their significance for seed germination. Environ. Exp. Bot. 72, 93–105. [ Links ]
Li, W., M. Khan, S. Yamaguchi, and Y. Kamiya. 2005. Effects of heavy metals on seed germination and early seedling growth of Arabidopsis thaliana. Plant Growth Regul. 46, 45-50. [ Links ]
Lintschinger, J., N. Fuchs, H. Moser, R. Jager, T. Hlebeina, G. Markolin, and W. Gossler. 1997. Uptake of various trace elements during germination of wheat, buckwheat and quinoa. Plant Food Hum. Nutr. 50(3), 223-237. [ Links ]
Liu, D., J. Zou, M. Wang, and W. Jiang. 2008. Hexavalent chromium uptake and its effects on mineral uptake, antioxidant defence system and photosynthesis in Amaranthus viridis L. Bioresour. Technol. 99, 2628-2636. [ Links ]
Marschner, P. (ed.). 2012. Marschner's mineral nutrition of higher plants. 3rd ed. Academic Press, London. [ Links ]
Mihoub A., A. Chaoui, and E. El Ferjani. 2005. Changements biochimiques induits par le cadmium et le cuivre au cours de la germination des graines de petit pois (Pisum sativum L.). C.R. Biologies 328, 33-41. [ Links ]
Milone, M.T., C. Sgherri, H. Clijsters, and F. Navari-Izzo. 2003. Antioxidative responses of wheat treated with realistic concentration of cadmium. Environ. Exp. Bot. 50, 265-276. [ Links ]
Ministerio de Agricultura y Desarrollo Rural. 2009. Anuario estadístico de frutas y hortalizas 2004-2008 y sus calendarios de siembras y cosechas. Bogota. [ Links ]
Miranda, D., C. Carranza, C. Rojas, G. Fischer, and J. Zurita. 2008. Acumulación de metales pesados en suelo y plantas de cuatro cultivos hortícolas, regados con agua del río Bogotá. Rev. Colomb. Cienc. Hortic. 2(2), 180-191. [ Links ]
Perfus-Barbeoch, L., N. Leonhardt, A. Vavasseur, and C. Forestier. 2002. Heavy metal toxicity: cadmium permeates through calcium channels and disturbs the plants water status. Plant J. 32, 539-548. [ Links ]
Ramos, I., E. Esteban, J. Lucena, and A. Garate. 2002. Cadmium uptake and subcellular distribution in plants of Lactuca sp. Cd-Mn interaction. Plant Sci. 162, 761-767. [ Links ]
Ranal, M. and D. de Santana. 2006. How and why to measure the germination process? Rev. Brasil. Bot. 29(1), 1-11. [ Links ]
Rahoui, S., A. Chaoui, and E. El Ferjani. 2010. Membrane damage and solute leakage from germinating pea seed under cadmium stress. J. Hazardous Materials 178, 1128 -113. [ Links ]
Rehman, F., N. Ahmad, K. Masood, J. Peralta-Videa, and F. Ahmad. 2009. Heavy metal toxicity in plants. pp. 71-98. In: Arshraf, M., M. Ozturk, and M. Ahmad (eds.). Plant adaptation and phytoremediation. Springer Science, London. [ Links ]
Rout, G., S. Sanghamitra, and P. Das. 2000. Effects of chromium and nickel on germination and growth in tolerant and nontolerant populations of Echinochloa colona (L.). Chemosphere 40, 855-859. [ Links ]
Scoccianti, V., R. Crinelli, B. Tirillini, V. Mancinelli, and A. Speranza. 2006. Uptake and toxicity of Cr(III ) in celery seedling. Chemosphere 64, 1695-1703. [ Links ]
Seregin, I. and D. Kozhevnikova. 2005. Distribution of cadmium, lead, nickel, and strontium in imbibing maize caryopses. Russ. J. Plant Physiol. 52, 565-569. [ Links ]
Shaukat, S., M. Mushtaq, and Z. Siddiqui. 1999. Effect of cadmium, chromium and lead on seed germination, early seedling growth and phenolic contents of Parkinsonia aculeata L. and Pennisetum americanum (L.) Schumann. Pak. J. Biol. Sci. 2, 1307-1313. [ Links ]
Shukla, P., S. Dubey, and U. Rai. 2007. Preferential accumulation of cadmium and chromium: Toxicity in Bacopa monnieri L. under mixed metal treatments. B. Environ. Contam. Toxicol. 78, 252-257. [ Links ]
SIA C. 2002. Rio Bogotá, un recurso hídrico en recuperación. Sistema de Información Ambiental de Colombia. http://www.siac.gov.co/contenido/contenido.aspx?catID=322&conID=415; consulta 25 de agosto de 2011. [ Links ]
Siddiqui, S., M. Meghvansi, M. Wani, and F. Jabee, 2009. Evaluating cadmium toxicity in the root meristem of Pisum sativum L. Acta Physiol. Plant. 31, 531-536. [ Links ]
Srivastava, S., R. Nigam, S. Prakash, and M. Srivastava. 1999. Mobilization of trivalent chromium in presence of organic acids: A hydroponic study of wheat plant (Triticum vulgare). Bull. Environ. Contam. Toxicol. 63, 524-530. [ Links ]
Street, R., G. Kulkarni, W. Stirk, C. Southway, and J. Van Staden. 2007. Toxicity of metal elements on germination and seedling growth of widely used medicinal plants belonging to hyacinthaceae. B. Environ. Contam. Toxicol. 79, 371-376. [ Links ]
Sung, Y., D.J. Cantliffem R.T. Nagata, and W.M. Nascimento. 2008. Structural changes in lettuce seed during germination at high temperature altered by genotype, seed maturation temperature, and seed priming J. Amer. Soc. Hort. Sci. 133(2), 300–311. [ Links ]
