










Services on Demand
Journal
Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.31 no.1 Bogotá Jan./Apr. 2013
SOILS, FERTILIZATION & MANAGEMENT OF WATER
Artropodes do solo durante o processo de decomposição da matéria orgánica
Edaphic arthropods during organic matter decomposition
Alexander Silva de Resende1, Eduardo Francia Carneiro Campello1 , Gabriela Tavares Arantes Silva2 , Khalil Menezes Rodrigues2, Willian Roberson Duarte de Oliveira2 e Maria Elizabeth Fernandes Correia1
1Agrobiologia, Empresa Brasileira de pesquisa Agropecuária (Embrapa). Seropédica (Brasil). alexander.resende@embrapa.br2Universidade Federal Rural do Rio de Janeiro. Seropédica (Brasil).
Received for publication: 12 May, 2010. Accepted for publication: 29 March, 2013.
RESUMO
Em sistemas agroflorestais (SA F), a poda de árvores e arbustos é usada como fonte de nutrientes para as culturas principais, tornando importante a determinação das taxas de decomposição dos mesmos. Entre os fatores que influenciam a decomposição de material vegetal, a fauna do solo tem um papel de destaque. O objetivo deste trabalho foi analisar a dinâmica entre as populações da fauna de artrópodes e o processo de decomposição de material vegetal. Para a avaliação, utilizaram-se sacolas (litter bags) contendo 50 g de folhas frescas de gliricídia ou ingá, dispostas sobre o solo em um SA F. O material foi coletado aos 8, 14, 20, 27, 41, 57 e 78 dias após a implantação do experimento, com duas coletas adicionais somente de ingá aos 128 e 239 dias. Depois da coleta, se determinou a matéria seca contida em cada sacola. Avaliou-se também a associação da fauna de artrópodes com cada material em cada coleta, através de funis Berlese-Tüllgren e triagem em laboratório. Na gliricídia houve um aumento na abundância da fauna a partir dos 14 dias, com um posterior declínio aos 41 dias. No ingá, o incremento da abundância da fauna só ocorreu a partir dos 57 dias. A fauna do solo varia durante o processo de decomposição e se apresenta de forma diferente em espécies de velocidades de decomposição contrastantes, sendo o teor de polifenol a característica que mais parece influenciar esse comportamento. Nem todos os grupos faunísticos acompanharam o processo de decomposição, mas o grupo Isopoda foi o que melhor acompanhou essa dinâmica.
Palavras chave: fauna do solo, sistemas agroflorestais, nutriente, mineralização.
ABTRACT
In agroforestry systems tree pruning is a source of nutrients for the main crops, which makes it important to determine the decomposition rate of these residues. Among the factors influencing plant material decomposition, soil fauna plays an outstanding role. Thus, the relation between these two parameters was the objective of the current work. The evaluation made use of litter bags filled with 50 g of leaves of gliricídia or ingá and scattered on the ground of an agroforestry system. These materials were collected on days 8, 14, 20, 27, 41, 57 and 78 after setting the experiment, plus two additional ingá sample collections on days 128 and 239. After collection, dry matter content and arthropod fauna (using Berlese-Tüllgren funis and laboratory screening) associated to plant material in each bag were determined. Gliricídia showed an increase in fauna abundance since day 14, to decline after day 41. In the case of ingá, fauna only increased after day 57. Soil fauna was observed to vary during the decomposition process and as a function of plant species decomposition rate, polyphenol content being the most influencing factor. While not all faunal groups accompanied the decomposition process, Isopoda was the one that most closely did so.
Key words: soil fauna, agroforestry, nutrients, mineralization
Introdução
Os sistemas agroflorestais (SAF) são formas de uso e manejo do solo nos quais espécies lenhosas (árvores, arbustos e palmeiras) são utilizadas em associação com culturas e/ou animais na mesma área, de maneira simultânea ou sequencial. Eles têm como objetivo promover o equilíbrio das funções ecológicas aliadas à rentabilidade econômica. Tais benefícios são provenientes das interações entre a componente arbórea e as demais componentes do sistema (Passos, 2003; Brienza Júnior et al., 2009).
Entre as funções ecológicas que os sistemas agroflorestais podem ter, destaca-se a ciclagem de nutrientes. Este processo inclui o enriquecimento das camadas superficiais do solo com matéria orgânica a partir da transferência de biomassa, sendo regulado principalmente pela velocidade de decomposição e mineralização deste material vegetal depositado (Silva et al., 2006).
Os sistemas biológicos envolvidos no processo de decomposição incluem três componentes: um recurso decomponente, ou seja folhas ou raízes que fornecem a matéria orgânica do solo; os microrganismos, que são mediadores da maior parte das transformações químicas; e macro-organismos, que criam condições convenientes para a atividade microbiana em escalas definidas de espaço e tempo (Wardle e Lavelle, 1997). Cabe ressaltar que a qualidade do material a ser decomposto é um dos fatores mais importantes. Neste sentido, sob as mesmas condições edafoclimáticas, a velocidade de decomposição dos resíduos de diversas espécies vegetais varia em função da qualidade do material decomponente (Gama-Rodrigues et al., 2003).
Os organismos do solo também podem ter efeitos reguladores fortes na decomposição (Lavelle, 1993), sendo classificados em microrganismos, compreendendo as bactérias e fungos; e a fauna do solo, que é dividida em: micro, meso e macrofauna (Correia e Oliveira, 2000).
Os microrganismos são a componente biótica que atua diretamente no processo de decomposição, pois tem o complemento enzimático mais apropriado para a quebra da serapilheira. A fauna do solo, principalmente a que consome diretamente os resíduos vegetais, desempenha o papel de transformadora da serapilheira (Wardle e Lavelle, 1997). Por causa do seu limitado aparato enzimático, a fauna do solo atua principalmente na fragmentação de detritos vegetais, acondicionando-os para a ação dos microrganismos; na humificação e na redistribuição de partículas orgânicas e minerais do solo (Correia e Oliveira, 2000). Assim, os microrganismos e a fauna do solo possuem funções complementares, sendo que a natureza da interação entre eles regula o processo de decomposição.
O objetivo deste trabalho foi observar a dinâmica da comunidade da fauna de artrópodes no processo de decomposição, verificando: 1) se há relação entre a abundância da fauna e a velocidade de decomposição da serapilheira de duas espécies vegetais com composição química contrastante; e 2) se todos os grupos presentes acompanham a taxa de decomposição.
Materiais e métodos
Amostras de folhas frescas de Gliricidia sepium (Jacq) Steud. (gliricídia) e Inga marginata Willd. (ingá) foram coletadas no mês de setembro de 2005. A coleta foi realizada na área jardinada da Embrapa Agrobiologia, município de Seropédica, RJ (22°46′ S, 43°41′ O) situada a 33 m de altitude. A região apresenta clima de verões úmidos e invernos secos, com temperatura média anual de 24,6°C e precipitação de 1.200 mm, sendo os meses de julho e agosto os mais secos (Ramos et al., 1973). A área possui um Planossolo de baixa fertilidade, com uma superfície arenosa e logo abaixo uma camada argilosa e compacta.
As amostras consistiram apenas nas folhas intermediárias dos ramos, evitando assim a coleta de folhas jovens e em estágio de maturidade avançada, toda vez que, com a senescência, há translocação de nutrientes entre os órgãos da planta (Mason, 1980). Amostras de 50 g de folhas frescas de cada espécie vegetal foram colocadas em bolsas de malha plástica com abertura de 5 mm (litter bags). Seis bolsas para cada material vegetal foram distribuídas aleatoriamente num sistema agroflorestal na Fazendinha Agroecológica da Embrapa Agrobiologia, sendo depositadas sobre a superfície do solo. A matéria seca inicial foi determinada antes da exposição a campo, secando-se as folhas de seis bolsas em estufa a 65°C até peso constante. Levada a cabo aos 8, 14, 20, 27, 41, 57 e 78 d após a instalação do experimento, a avaliação da velocidade de decomposição e liberação de nutrientes contou com três repetições por espécie e por data de amostragem. Para Inga semialata, realizaram-se duas coletas adicionais, aos 128 e 239 d, por sabidamente apresentar decomposição mais lenta que a gliricídia (Silva et al., 2006).
Em cada data de amostragem, as litter bags foram retiradas do solo e transportadas para o laboratório, onde o material vegetal foi seco em estufa a 65°C até atingir peso constante. Em seguida foi manualmente triado e triturado para a determinação do teor de nitrogênio segundo metodologia descrita por Embrapa (1999).
A avaliação da fauna do solo e do material em decomposição contou com seis repetições por data de amostragem e por espécie. Uma vez retiradas as litter bags, coletaram-se amostras de solo utilizando um gabarito de 0,25 x 0,25 m até uma profundidade de 5 cm. Cada amostra foi transferida para o laboratório, sendo adicionada em funis da bateria de extratores Berlese-Tüllgren (Correia e Oliveira, 2000), os quais tinham em sua base um recipiente de vidro contendo 150 mL de álcool (50%). Após a transferência de todas as amostras para os funis, a bateria de extratores foi vedada completamente. Acima dos funis, acenderam-se lâmpadas de 40 W, as quais permaneceram assim por 13 d, conforme recomendado por Correia e Oliveira (2000). O conteúdo de cada frasco proveniente dos extratores foi triado, em placas de Petri sob lupa binocular. Os artrópodes foram separados, contados e identificados ao nível de ordem e os dados expressos em número de indivíduos por m2.
A avaliação dos resultados baseou-se na estatística descritiva, estimando-se a média, o desvio e o erro padrão da média.
Resultados e discussão
As duas espécies vegetais apresentaram taxas de decomposição bem contrastantes. A gliricídia apresentou massa final de apenas 9% do valor inicial aos 78 d, enquanto o ingá ainda manteve 72% da sua massa inicial aos 239 d (Fig.1). Esses resultados são reflexos da composição química de cada espécie. Após 14 d de exposição a campo, a gliricídia apresentou uma relação (lignina + polifenol)/N duas vezes menor que o ingá. O mesmo foi observado para a relação C:N, que foi de 19 para o ingá e de 14 para a gliricídia (Tab.1). Esses resultados estão de acordo com Silva et al. (2006), quem, trabalhando na mesma área, encontraram correlação negativa entre os parâmetros supracitados e a velocidade de decomposição.
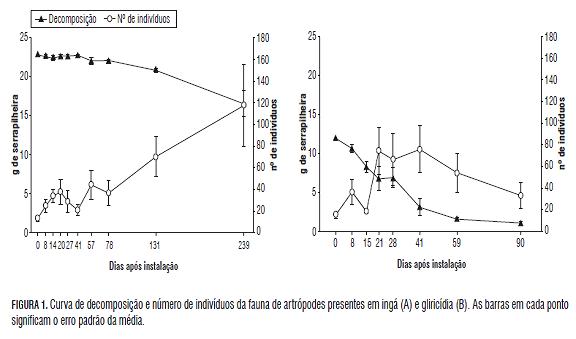
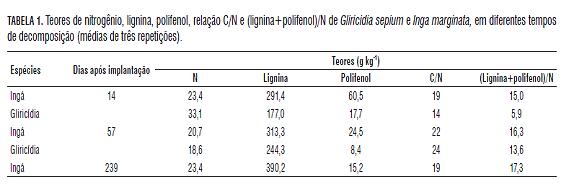
A presença de altos teores dessas moléculas implica numa redução na taxa de decomposição. A lignina interage com a parede celular fornecendo proteção mecânica à celulose contra a degradação. Já os polifenóis podem formar complexos com as proteínas e a parede celular (tornando-as física e quimicamente inacessíveis aos decompositores) ou reagir com grupamentos amino, acarretando uma correlação negativa com a mineralização do nitrogênio (Mafongoya et al., 1998).
O efeito da qualidade do resíduo vegetal sobre a biota do solo regula a extensão pela qual essa mesma biota facilita a decomposição da serapilheira (Wardle e Lavelle, 1997). Assim, resíduos com qualidades diferentes apresentarão valores diferentes no número total de indivíduos presentes, o que por sua vez, poderá alterar a velocidade de decomposição. Neste trabalho observaram-se valores distintos na abundância da fauna de artrópodes presente nas diferentes datas de coleta, possivelmente mais associados às condições climáticas do que a fatores relacionados às características químicas do material (Tab.2).
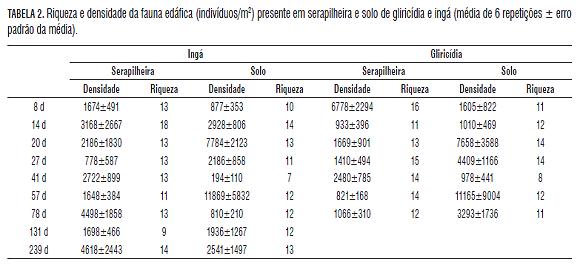
Foram encontrados 21 grupos diferentes de artrópodes, sendo que os mais numerosos foram: Acari, Formicidae, Coleoptera, Diptera, Isopoda, Collembola, Hemiptera e Heteroptera. Os outros grupos foram representados por apenas alguns indivíduos.
A variação da comunidade de artrópodes ocorreu de forma bastante aleatória, não sendo possível determinar um padrão de comportamento. Esse resultado corrobora a noção de que os grupos da fauna edáfica apresentam funções diversas na dinâmica do solo, podendo participar de forma direta ou indireta no processo de decomposição, o que leva à formação de complexas relações competitivas e mutualísticas entre diferentes grupos (Correia e Oliveira, 2000). Mudanças na qualidade da serapilheira também são citadas como passíveis de influenciar as interações bióticas e, portanto, a composição taxonômica da população de organismos do solo, mas não em direções previsíveis (Wardle e Lavelle, 1997; Gatiboni et al., 2009).
Acari foi o mais numeroso dentre os grupos encontrados, correspondendo a 69% do total de indivíduos (Tab.3). Este grupo é muito abundante e amplamente distribuído na natureza. Várias das suas espécies tendem a ser partenogenéticas, promovendo maior produção de ovos, favorecendo a sua distribuição e, algumas vezes, um aumento considerável da população (Franklin et al., 2004; Tripathi et al., 2005). Embora haja espécies decompositoras, sua alta densidade pode não resultar em maior influência sobre a decomposição (Franklin et al., 2004; Tripathi et al., 2005).
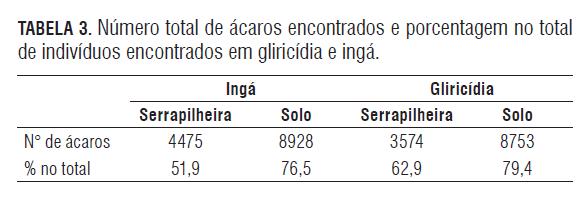
Estudos têm mostrado que o grupo Acari de artrópodes raspadores pode ser mais importante na mobilização de nutrientes do que na contribuição à perda de massa (Hanlon e Anderson, 1979). Eles possuem um aparato bucal capaz de raspar a matéria orgânica no momento em que estão se alimentando da microflora aderida aos resíduos (Seartedt, 1984), o que lhes permite manter altos níveis populacionais, embora com porca influência sobre as primeiras etapas do processo de decomposição.
Já o grupo Formicidae possui estrutura social, tendendo a se localizar em determinados pontos. As formigas recolhem resíduos ricos em nutrientes da área que rodeia o ninho e os concentram perto da entrada do mesmo, tornando o solo mais rico em matéria orgânica, N e P do que o solo ao redor (Wagner e Jones, 2006). O grupo exerce influência sobre a decomposição, no entanto, a direção é muito pontual, gerando um ambiente heterogêneo e, portanto, não apresentando grande relevância em estudos da magnitude do presente.
Devido a sua reduzida participação no processo de decomposição, Acari e Formicidae foram excluídos da análise, a qual mostrou uma correlação direta entre o número de indivíduos de cada grupo faunístico e a velocidade de decomposição das espécies vegetais estudadas (Fig.1).
Para a gliricídia, observou-se um aumento na abundância da fauna a partir dos 14 d, quando a taxa de decomposição passou a ser mais intensa, com um posterior declínio a partir dos 41 d, quando a quantidade de massa remanescente era inferior ao 30% da existente inicialmente. Para o ingá, o incremento na abundância da fauna só ocorreu a partir dos 57 d (coincidindo com o começo da perda de matéria seca nos resíduos) e continuou até os 239 d. Avaliando-se as características químicas do material nessas datas (Tab.1), nota-se que os teores de polifenol dos resíduos de ingá diminuíram de 60 para 25 g kg-1 entre os 14 e 57 d após a implantação do experimento. Embora sejam necessários estudos mais detalhados, os resultados sugerem que o teor de polifenol contido nos resíduos vegetais é determinante para o crescimento da população dos agentes decompositores da fauna do solo e, consequentemente, para a taxa de decomposição do material. A partir destes resultados, buscar-se-á um valor ótimo de liberação de compostos recalcitrantes, a partir do qual se desencadeie o processo de decomposição.
Essa diferença de comportamento da fauna caracteriza a influência de resíduos com diferentes composições químicas. Em geral, os resíduos com menor relação C:N são mais vulneráveis ao efeito dos agentes da decomposição, acelerando essa perda. Além disso, esses resíduos oferecem menos resistência à ação da fauna saprófaga, para a qual se tornam mais atraentes, influenciando assim positivamente o tamanho da comunidade faunística e oferecendo melhores condições para os microrganismos. Estes, pela sua vez, irão alterar a relação C: nutriente, favorecendo assim uma nova colonização de artrópodes saprófagos (Tian et al., 1997). Com o passar do tempo, a fração mais lábil da serapilheira reduze-se consideravelmente, restando unicamente a fração recalcitrante, que, composta basicamente de celulose e lignina, vem a ser mais resistente.
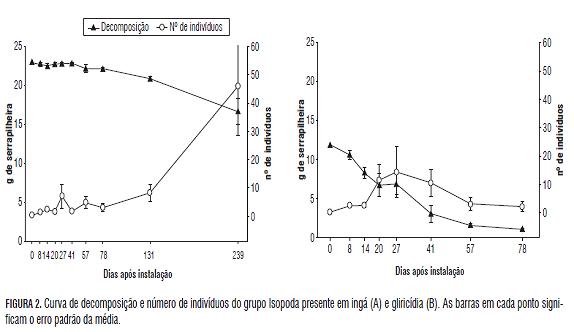
Os grupos encontrados não mostraram padrão algum de variação de densidade, mas, entre eles, Isopoda apresentou o comportamento mais representativo da fauna saprófaga como um todo (Fig.2). O grupo é essencialmente saprófago, com sua dieta composta na maior parte de material orgânico deteriorado, como serapilheira, madeira apodrecida, fungos e capas bacterianas. é citado como um regulador- -chave das funções de decomposição e ciclagem de nutrientes no ambiente, e a sua diversidade diminui rapidamente em paisagens agrícolas e silviculturais intensivamente trabalhadas e em ambientes pobres em que a matéria orgânica é escassa ou de baixa qualidade (Paoletti e Hassall, 1999), o que demonstra que o grupo é extremamente sensível às mudanças ambientais. Estes resultados reforçam a importância do grupo como um potencial indicador em estudos sobre a fauna do solo e a decomposição da serapilheira.
Conclusão
A fauna do solo varia durante o processo de decomposição de resíduos vegetais e se apresenta de forma diferente em espécies de velocidades de decomposição contrastantes. O teor de polifenol é a característica que mais parece influenciar este comportamento. O grupo Isopoda é o que apresenta maior correspondência com o processo de decomposição.
Agradecimentos
À Embrapa Agrobiologia, Universidade Federal Rural do Rio de Janeiro, CNP q e Faperj pelo apoio e suporte financeiro e pela bolsa de produtividade em pesquisa do primeiro autor.
Literatura citada
Brienza Júnior, S., R.Q. Maneschy, M. Mourão Júnior, A.B. Gazel Filho, J.A.G. Yared, D. Gonçalves e M.B. Gama. 2009. Sistemas agroflorestais na amazônia brasileira: análise de 25 anos de pesquisa. Pesq. Flor. Bras. 60, 67-76. [ Links ]
Correia, M.E.F. e L.C. Oliveira. 2000. Fauna de solo: aspectos gerais e metodológicos. Documento Técnico No. 112. Embrapa Agrobiologia, Seropédica, Brasil. [ Links ]
Embrapa, Empresa Brasileira de Pesquisa Agropecuária. 1999. Manual de análises químicas de solos, plantas e fertilizantes. Embrapa Informação Tecnológica, Brasília. [ Links ]
Franklin, E., T. Hayek, E.P. Fagundes e L.L. Silva. 2004. Oribatid mite (Acari: Oribatida) contribution to decomposition dynamics of leaf litter in primary forest, second growth, and polyculture in the central Amazon. Braz. J. Biol. 64, 59-72. [ Links ]
Gama-Rodrigues, A.C., N.F. Barros e M.L. Santos. 2003. Decomposição e liberação de nutrientes do folhedo de espécies florestais nativas em plantios puros e mistos no sudeste da Bahia. Rev. Bras. Ciênc. Solo 2, 1021-1031. [ Links ]
Gatiboni, L.C., J.L.M. Coimbra, L.P. Wildner e R.B.N. Denardin. 2009. Modificações na fauna edáfica durante a decomposição da palhada de centeio e aveia preta em sistema plantio direto. Biotemas 22(2), 45-53. [ Links ]
Hanlon, R.D. e J.M. Anderson. 1979. The effects of Collembola grazing on microbial activity in decomposing leaf litter. Oecologia 38, 93-99. [ Links ]
Lavelle, P., E. Blanchart, A. Martin, S. Martin, A. Spain, F. Toutain, I. Barois e R.A. Schaefer. 1993. Hierarchical model for decomposition in terrestrial ecosystems: application to soils of the humid tropics. Biotropica 25(2), 130-150. [ Links ]
Mafongoya, P.L., K.E. Giller, e C.A. Palm. 1998. Decomposition and nitrogen release patterns of tree pruning and litter. Agrof. Syst. 38, 77-97. [ Links ]
Mason, C.F. 1980. Decomposição. Universidade de São Paulo, São Paulo, Brasil. [ Links ]
Paoletti, M. G., Hassall, M. Woodlice. 1999. (Isopoda: Oniscidea): their potential for assessing sustainability and use as bioindicators. Agric. Ecosyst. Environ. 74, 157-165. [ Links ]
Passos, C.A.M. 2003. Aspectos gerais dos sistemas agroflorestais. Universidade Federal do Mato Grosso, Cuiabá, Brasil. [ Links ]
Ramos, D.P., A.F. Castro, e N.M. Camargo. 1973. Levantamento detalhado de solos da Aréa da Universidade Federal Rural do Rio de Janeiro. Pesq. Agropec. Bras. 8, 1-27. [ Links ]
Seartedt, T.R. 1984. The role of microarthropods in decomposition and mineralization processes. Ann. Rev. Entomol. 29, 25-46. [ Links ]
Silva, G.T.A., L.V. Matos, P. de O. Nóbrega, E.F.C. Campello, e A.S. de Resende. 2006. Correlação entre a composição química e a velocidade de decomposição e liberação de nitrogênio de folhas de dez espécies em um sistema agroflorestal (CD-ROM). In: VI Congresso Brasileiro de Sistemas Agroflorestais. Rio de Janeiro, Brasil. [ Links ]
Tian, G., L. Brussaard, B.T. Kang e M.J. Swift. 1997. Soil faunamediated decomposition of palnt residues under constrained environmental and residue quality conditions. pp. 107-123.In: Cadisch, G. e K.E. Giller (eds.). Driven by nature: plant litter quality and decomposition. CAB International, Wallingford, UK. [ Links ]
Tripathi, G., R. Kumari e B.M. Sharma. 2005. Associação de mesofauna do solo com decomposição de restos vegetais. Científica 33(2), 148-151. [ Links ]
Wagner, D. e J.B. Jones. 2006. The impact of harvester ants on decomposition, N mineralization, litter quality, and availability of N to plants in the Mojave Desert. Soil Biol. Biochem. 38, 2593-2601. [ Links ]
Wardle, D.A. e P. Lavelle. 1997. Linkages between soil biota, plant litter quality and decomposition. pp. 107-123. In: Cadisch, G. e K.E. Giller (eds.). Driven by nature: plant litter quality and decomposition. CAB International, Wallingford, UK. [ Links ]
Zimmer, M. 2004. Effects of temperature and precipitation on a flood plain isopod community: A field history. Eur. J. Soil Biol. 40, 130-146, [ Links ]
