










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.31 no.2 Bogotá May/Aug. 2013
CROP PHYSIOLOGY
Evaluation of low light intensity at three phenological stages in the agronomic and physiological responses of two rice (Oryza sativa L.) cultivars
Evaluación de la baja intensidad lumínica en tres etapas fenológicas sobre la respuesta agronómica y fisiológica en dos cultivares de arroz (Oryza sativa L.)
Hermann Restrepo1 and Gabriel Garcés 2
1Department of Agronomy, Faculty of Agronomy, Universidad Nacional de Colombia. Bogota (Colombia). hrestrepod@unal.edu.co2Sectional Saldaña, Fedaración Nacional de Arroceros (Fedearroz). Saldaña (Colombia).
Received for publication: 1 February, 2012. Accepted for publication: 5 June, 2013.
ABSTRACT
A study was carried out to assess the effect of two irradiance levels (100 and 50% natural light) at three different growth stages on yield and physiological characteristics of two rice cultivars (Fedearroz 50 and Fedearroz 733). The plants were exposed to low irradiance for 5 days consecutive by using black net cloth (about 50% of normal) at the panicle primordium, flowering and grain filling stages. The obtained results showed that the leaf chlorophyll content (SPAD readings) was higher in rice leaves under low irradiance. The chlorophyll content from the shade treatment apparently remained constant until the grain filling phase, then decreased slightly in the"F50" rice plants and remarkably in the"F733" plants. Stomatal conductance (gs) was negatively affected by shading, with the effect being more adverse in the"F733" plants. At the flowering and grain filling phases, grain yield was reduced by the low light treatments by around ∼20% in the"F50" plants. While, in"F733", they were only affected by shading at the grain filling stage, causing a decrease of around 25%. These results seem to indicate that "F773" may have a better capacity for partitioning dry matter than "F50" in spite of the fact that the gas exchange characteristics were conditioned by low irradiance conditions at the reproductive and ripening phases.
Key words: shading, yield, stomatal conductance, growth stages.
RESUMEN
Un estudio fue desarrollado para estimar el efecto de dos niveles de radiación (100% y 50% de la radiación solar) en tres diferentes fases de rendimiento y en las características fisiológicas de dos cultivares de arroz (Fedearroz 50 and Fedearrozz F733). Las plantas se expusieron al sombramiento por 5 días consecutivos usando una polisombra (cerca del 50% de lo normal) al inicio del primordio floral, floración y llenado de grano. Los resultados obtenidos mostraron que el contenido de clorofila (lecturas SPAD) era mayor en las hojas de arroz bajo sombramiento. El contenido de clorofilas de hojas procedentes del tratamiento de baja radiación permaneció constante hasta llenado de grano, luego disminuyó ligeramente en plantas de"F50" y considerablemente en plantas de "F733". La conductancia estomática (gs) fue afectada negativamente por el sombramiento, siendo el efecto negativo más notorio en las plantas de "F733". El rendimiento también disminuyó por causa del sombramiento alrededor de un ∼20% en plantas de "F50" en las etapas fenológicas de floración y llenado de grano. Mientras,"F733" solamente fueron afectadas por el sombramiento en llenado de grano, causando una disminución del ∼25%. Estos resultados parecen indicar que "F733" podrían tener una mejor capacidad en distribuir la materia seca que "F50" a pesar del hecho que las propiedades de intercambio gaseoso de la hoja estuvieron condicionadas por la condición de baja radiación en las etapas reproductivas o de maduración.
Palabras clave: sombramiento, rendimiento, conductancia estomática, etapas de desarrollo.
Introduction
Rice (Oryza sativa L.) is the world"s single most important food crop, being the primary food source for more than one-third of the world"s population (Shaiful-Islam et al., 2009). In Colombia, rice occupies a total area of 420,721 ha, with a production of 2,262,055 t in 2010 (Fedearroz, 2011). In Colombian conditions, light intensity varies throughout the year, causing a significant effect on the growth and productivity of rice (Castilla et al., 2010). A reduction of around 20-50% of the solar radiation has been reported in rice-growing areas at certain periods of the year in Colombia (Diago and Barrero, 2003; Garcés et al., 2005). The yield of field-grown rice mainly depends on the solar radiation throughout the growth period, especially during the reproductive and/or grain filling stages (Fageria, 2007). Low irradiance (LI) during the reproductive and/or ripening stages has an adverse effect on potential yield because the photosynthetic activity in the leaves of rice cultivars decreases (Srivastava, 2011). In addition, rice plants in low irradiance environments have shown physiological responses such as: changes in chlorophyll and rubisco content (Hidema et al., 1991).
As mentioned above, agronomical, morphological and physiological responses of rice under low irradiance conditions have been widely reported on by several authors (Viji et al., 1997; Lakshmi-Prada et al., 2004; Singh, 2005; Fageria, 2007; Moula, 2009). However, reports on the same are not available in Colombia. Consequently, understanding the physiological responses of Colombian rice cultivars to LI environments is important for developing crop management techniques or breeding programs in order to improve yield potential. For these reasons, the aim of this paper was to study the influence of two irradiance levels (0 and 50% shading) during initiation of the panicle primordium, flowering or grain-filling stages on yield components and physiological mechanisms such as chlorophyll fluorescence, stomatal conductance, and photosynthesis in two rice cultivars (cv. Fedearroz 50 (F50) and cv. Fedearroz 733 (F733)). "F50" rice plants have been widely grown by Colombian growers during the last decade, however; this cultivar has shown a reduction in cultivated area due to its high susceptibility to low irradiance periods. On the other hand, "F733" rice plants have been recently introduced to rice growers and little is known about its physiological response to LI condition in rice-growing areas of Colombia.
Material and methods
A field study was carried out between June and October of 2010 at "Centro de Investigación Las Lagunas" experimental farm in Saldaña (3°54"47,45"" N, 74°59"8,47"" W), Colombia. Two indica rice cultivars, "F50" and "F733", were used. The plants were grown on sandy loam soil that contained organic matter at 1.5%. Each rice cultivar was planted on plots (6 x 7 m). Each plot was provided with 230 kg ha-1 N, 36 kg ha-1 P2O5, and 96 kg ha-1 K2O as a top dressing about 45 d before heading. The average daytime temperature (06:00-18:00 HR) was ∼29.5°C. The average nighttime temperature (18:00-06:00 HR) was ∼25.6°C. Relative humidity was 68.1% in the daytime and 85.3% at night. The maximum irradiance at noon was about 1516 W m-2. The experiment lasted 120 d.
Both cultivars were split into four treatments: a) plants without shading (control), b) plants under shaded conditions at the initiation of the panicle primordium (IP), c) at flowering and d) plants under shade at grain-filling (GF). Plants were exposed to low irradiance for five consecutive days by using black net cloth (about 50% of the solar radiation) in order to achieve short-term low irradiance stress. Then, the black net cloth was removed.
Measurements of net photosynthetic gas exchange (Pn) and stomatal conductance (gs) were taken on a fully expanded attached leaf of rice seedlings by a portable photosynthesis system (LI-6200; LICOR, Lincoln, NE) using a 250-cm3 cuvette, and a leaf-porometer (SC - 1 Decagon Devices, Pullman, WA), respectively. Three sets of 3-d measurements (referred to as initiation of the panicle primordium (IP), flowering (F) and grain filling (GF), respectively) were carried out between 09:00 and 13:00 HR. During all photosynthesis measurements, conditions within the cuvette were: PAR greater than 800 µmol m-2 s-1, leaf temperature 27±5°C, and leaf to air water vapor pressure difference 1.8±0.5 kPa.
Leaf chlorophyll fluorescence measurements were carried out on 31 July (IP), 20 August (F) and 13 September (GF) using a continuous excitation chlorophyll fluorescence analyzer (Handy PEA; Hansatech Instruments, Kings Lynn, UK). The leaves were acclimated to the dark using light weight leaf clips for at least 20 min before measurements were taken. Baseline (F0) and maximum (Fm) fluorescences were measured and variable (Fv = Fm - F0) fluorescence and the ratio of variable fluorescence to maximum fluorescence (Fv/Fm) were calculated from these data. Chlorophyll readings were also taken using a chlorophyll meter (SPAD-502; Minolta, Ramsey, NJ) as a nondestructive tool for estimating leaf Chlorophyll (Markwell et al., 1995). SPAD readings were also recorded on 31 July (IP), 20 August (F) and 13 September (GF).
The trial was manually harvested on 5 Oct. The yield paddy was approximately estimated with a grain content moisture of 14%. The grain yield was calculated by using the following formula:
kg ha-1=(kg/plot per 42 m2) x 10,000 (1)
Means and standard errors were obtained for each data. The data were subjected to analysis of variance by using a factorial design. Where a significant F-test was observed, mean separation among the treatments was obtained by Tukeys test. The data were analyzed using Statistix (version 8.0; Analytical Software, Tallahassee, FL).
Results and discussion
Varietal differences were only observed in our experiment on the number of tillers and panicles (Tab. 1). The "F50" rice plants had a higher number of tillers and panicles than the "F733" rice plants. Studies performed by Venkateswarlu (1977) reported that low light intensity treatments carried out from the early vegetative growth stage until harvesting time significantly reduced the number of tillers and panicles in rice plants. In addition, Fageria (2007) stated that tillering is determined by genetic characteristics during the vegetative growth phase, and is also influenced by environmental factors such as large stress due to low irradiance. In our experiment, the lack of differences for the tiller number between the rice cultivars in the shading treatments at three different phenological stages could be due to the fact that the LI treatments were not sufficiently prolonged in order to cause a negative effect on this yield component, since our treatments were only carried out for five consecutive days in the IP, F and GF phases. Regarding the number of panicles per square meter, Srisvastava (2011) reported that this yield component can also be conditioned by low light intensity; however, Fageria (2007) observed that the number of panicles per square meter are correlated with the number of tillers obtained in the vegetative growth phase. In this trial, it was observed that the LI treatments did not have any negative effects on the tillering capacity of the rice cultivars, meaning that the number of panicles per square meter was not affected.
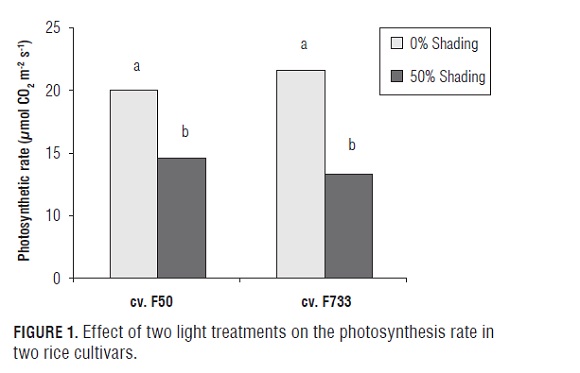
Differences were found in the light treatments for the leaf photosynthesis rate (Fig.1). The rice plants under low irradiance conditions had a lower net leaf photosynthesis than the plants under full sun light conditions in both cultivars.Shading caused a decrease in the carbon exchange rate by around 60 and 45% in plants of the cv. F733 and cv. F50, respectively. On other hand, stomatal conductance (gs) was negatively affected by shading, with the more adverse effect seen in the "F733" rice plants (Fig.2 A and B). In the "F50" rice plants under full sun light conditions, differences were not observed for gs throughout the different phenological phases of the experiment. When the "F50" rice plants were exposed to LI treatments, gs diminished by ∼28, ∼25 and ∼35% in the IP, F, and GF stages, respectively (Fig.2A). Similar trends were found regarding the "F733" rice plants. Nevertheless, the negative effect of the shading was higher in this cultivar at the grain filling stage, since gs decreased by ∼50% under this stress condition (Fig.2B). Overall, rice leaves in both cultivars showed a low gas exchange capacity under shading, but above all, in the "F733" plants. Studies performed by Murchie et al. 2002 on "IR 72" rice plants (indica cultivar) also presented a lack of acclimation of the photosynthetic rate to LI condition in rice leaves. The low response of the rice leaves to the shading treatment could be due to the fact that acclimation to irradiance in rice is based on a morphological alteration of the leaf area ratio rather than of the photosynthetic components per se (Makino et al., 1997).

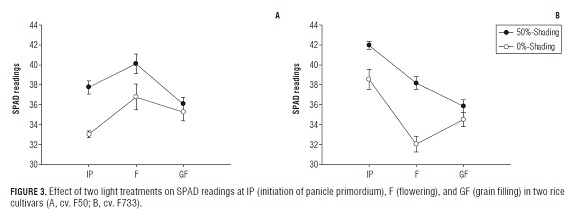
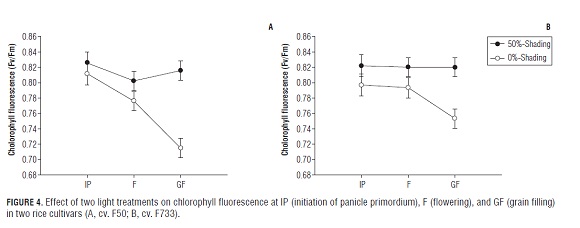
Changes in SPAD readings are shown in Fig.3 (A and B). According to the SPAD readings, the leaf chlorophyll content was higher in rice leaves under low irradiance than in rice leaves exposed to full sun light conditions in both cultivars. In addition, the chlorophyll content from the shade treatment apparently remained constant until the GF phase, then decreased slightly in the "F50" rice plants and noticeably in the "F733" rice plants. Differences were not found for SPAD readings among the treatments at the grain filling stage in both cultivars. Results obtained by Murchie et al. (2002) also found that values for chlorophyll declined in rice leaves under high irradiance conditions. Furthermore, studies carried out by Venkateswarlu et al. (1977) and Hidema et al. (1991) showed that leaf chlorophyll content was lower in full sun conditions than in low irradiance environments. Differences in the leaf chlorophyll content among the light treatments could be the response of the plant to increase its protection against excess light, since plants show an increase in the xanthophyll cycle pool size under full light conditions (Bilger et al., 1995), and a regulated loss of chlorophyll and pigment proteins per chloroplast (Anderson, 1986). A decline in leaf chlorophyll content at the GF phase is mainly due to leaf senescence which can cause a more rapid decrease in the reaction centers and core chlorophyll than in the light harvesting center of photosystem II. On the other hand, the maximum efficiency of PSII photochemistry (Fv/Fm) showed no large changes under the shade treatments during the experiment in both rice cultivars. The Fv/Fm ratio decreased only slightly from around 0.83 at the F phase to around 0.73 at the GF phase in both rice varieties exposed to full sun conditions (Fig.4 A and B). Hirotsu et al. (2005) also observed little changes during leaf senescence for efficiency of PSII photochemistry (Fv/Fm) in rice leaves under high irradiance. In our experiment, it was observed that the Fv/Fm ratio diminished in sun plants in both cultivars at the GF stage. This means that a decrease in Fv/Fm is widely used as an indicator of photoinhibition (Hirotsu et al., 2005), furthermore, there is little change in the structural rearrangements and the rate of energy transfer between complexes in the remaining PSII apparatus; otherwise, Fv/Fm would show a significant decrease during leaf senescence (Tang et al., 2005).
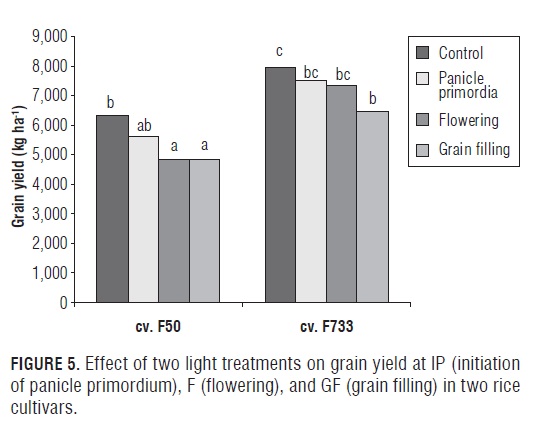
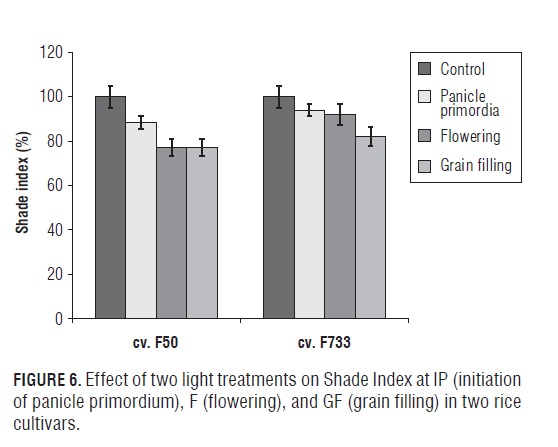
Differences were observed in the triple interaction cultivar x light treatments x phenological phase on grain yield (Fig.5). At the (F) and (GF) phases, grain yield was reduced by shading by around ∼20% in the "F50" rice plants. While, the "F733" rice plants were only affected by shading at the GF stage, causing a decrease of around 25%. On the other hand, similar trends were observed when the data were expressed as shade index (%) (grain yield under shading over grain yield of the control) (Fig.6). Our results indicated that the cultivars clearly differed in their yield performance under shade at different phenological phases due to irradiance (solar radiation) stress, with "F733" being superior to "F50" at the F and GF stages. In addition, the shade index was similar, 77% for the "F50" rice plants at the flowering and grain filling phases; but, "F733" had a greater shade index at 82%. Likewise, the photosynthetic behavior of "F733" was particularly affected by shading at the GF stage, showing a decline of around 60% as compared to 45% in the "F50" plants. The obtained results seem to indicate that "F733" has an advantage over "F50" in the partitioning of dry matter, since "F50" can depend more on current photosynthetic contributions to grain yield, while "F733" can utilize more of the accumulated photosynthetic reserve. The present findings are in agreement with the observations of Venkateswarlu et al. (1977) and Singh (2005), who also concluded that low irradiance treatments significantly diminished grain yield, particularly during the reproductive and ripening phases, indicating that photosynthetic activity during the reproductive and ripening stages had profound influence on the yield in rice cultivars.
Conclusion
In summary, the acclimation response of Colombian rice cultivars to shading may be important because light levels change throughout the year in rice-growing areas. This is reflected in a variation for the grain yield (Diago and Barrero, 2003; Garcés et al., 2005). However, "F733" yields are consistently and significantly higher than "F50" ones under low irradiance, mainly, at the ripening stages. We suggest that this is at least partially due to the better capacity for partitioning dry matter in "F733" plants in spite of the fact that gas exchange measurements were negatively affected by shading treatments. Further studies are necessary in order to assess partitioning patterns of Colombian rice cultivars under low light intensity.
Literature cited
Anderson, J.M. 1986. Photoregulation of the composition, function and structure of thylakoid membranes. Annu. Rev. Plant Physiol. Plant Mol. Biol. 37, 93-136. [ Links ]
Bilger, W., J. Fisahn, W. Brummet, J. Kossmann, and L. Willmitzer. 1995. Violaxanthin cycle pigment contents in potato and tobacco plants with genetically reduced photosynthetic capacity. Plant Physiol. 108, 1479-1486. [ Links ]
Castilla, L.A., D. Pineda, J. Ospina, J. Echeverry, R. Perafan, G. Garcés, J. Sierra, and A. Díaz. 2010. Cambio climático y producción de arroz. Rev. Arroz 58, 4-11. [ Links ]
Diago, M, and B. Barrero. 2003. Rendimiento del arroz en el Espinal y el Guamo durante 2002. Rev. Arroz 51, 15-18. [ Links ]
Fageria, N.K. 2007. Yield physiology of rice. J. Plant Nutr. 30, 843-879. [ Links ]
Fedearroz. 2011. Durante semestre B-2010 arroz crece en áreas y mejora rendimientos. Rev. Arroz 5, 4-9. [ Links ]
Garcés, G., P. Garcés, and M. Diago. 2005. Resultados de monitoreo de cosecha 2004 Sur del Cesar. pp. 23-26. In: Compendio resultados de investigación 2003-2005. Federación Nacional de Arroceros; Fondo Nacional del Arroz, Bogota. [ Links ]
Hirotsu, N., A. Makino, S. Yokota, and T. Mae. 2005. The photosynthetic properties of rice leaves treated with low temperature and high irradiance. Plant Cell Physiol. 46, 1377-1383. [ Links ]
Hidema, J., A. Makino, T. Mae, and K. Ojima. 1991. Photosynthetic characteristics of rice leaves aged under different irradiances from full expansion through senescence. Plant Physiol. 97, 1287-1293. [ Links ]
Lakshmi-Prada, M., M. Vanangamudi, and V. Thandapi. 2004. Effect of low light on yield and physiological attributes of rice. IRRN 29(2), 71-73. [ Links ]
Makino, A., T. Sato, H. Nakano, and T. Mae. 1997. Leaf photosynthesis, plant growth and nitrogen allocation in rice under different irradiances. Planta 203, 390-398. [ Links ]
Markwell, J., J.C. Osterman, and J.L. Mitchell. 1995. Calibration of the Minolta SPAD-502 leaf chlorophyll meter. Photosynth. Res. 46, 467-472. [ Links ]
Moula, G. 2009. Effect of shade on yield of rice crops. Pakistan J. Agric. Res. 22, 24-27. [ Links ]
Murchie, E.K., S. Hubbart, Y. Chen, S. Peng, and P. Horton. 2002. Acclimation of rice photosynthesis to irradiance under field conditions. Plant Physiol. 130, 1999-2010. [ Links ]
Shaiful-Islama, M., M. Hasanuzzamanb, M. Rokonuzzamanc, and K. Nahard. 2009. Effect of split application of nitrogen fertilizer on morphophysiological parameters of rice genotypes. Intl. J. Plant Prod. 3, 51-62. [ Links ]
Singh, S. 2005. Effect of low-light stress at various growth phases on yield and yield components of two rice cultivars. IRRN 30(2), 36-37. [ Links ]
Srivastava, G.C. 2011. Crop physiology. Biotech Books, New Delhi. [ Links ]
Tang, Y., X. Wen, and C. Lu. 2005. Differential changes in degradation of chlorophyll-protein complexes of photosystem I and photosystem II during flag leaf senescence of rice. Plant Physiol. Bioch. 43, 193-201. [ Links ]
Venkateswarlu, B. 1977. Influence of low light intensity on growth and productivity of rice, Oryza sativa L. Plant Soil 47, 713-719. [ Links ]
Venkateswarlu, B., V.V.S.S. Prasad, and A.V. Rao. 1977. Effects of low light intensity on different growth phases in rice (Oryza sativa L.). Plant Soil 47, 37-47. [ Links ]
Viji, M.M., M. Thangaraj, and M. Jayapragasam. 1997. Low irradiance stress tolerance (Oryza sativa L.). Biol. Plant. 39, 251-256. [ Links ]
