










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkAgronomía Colombiana
versión impresa ISSN 0120-9965
Agron. colomb. vol.32 no.1 Bogotá ene./abr. 2014
https://doi.org/10.15446/agron.colomb.v32n1.38714
http://dx.doi.org/10.15446/agron.colomb.v32n1.38714
PROPAGATION AND TISSUE CULTURE
1 Special Technical Assistance Project, Cooperativa de Caficultores del Sur del Tolima (CAFISUR). Chaparral (Colombia).
2 Department of Agronomy, Faculty of Agricultural Sciences, Universidad Nacional de Colombia. Bogota (Colombia). galigarretom@unal.edu.co
Received for publication: 11 July, 2013. Accepted for publication: 19 March, 2014.
ABSTRACT
Andean blueberry propagation is done with wild plants and there is an absence of knowledge for the agronomical potential of the available materials for sexual propagation. Therefore, developing a propagation protocol that provides technical information for field growers is needed. The present paper aimed to evaluate the effect of the size of the fruits that were used to extract the seeds on the growth and development of Andean blueberry plants during the germination and nursery stages, comparing fruits from four different regions of Colombia: Antioquia, Cauca, Cundinamarca and Boyaca. During the germination stage, seeds from Antioquia and Cauca showed the best results with germination rates close to 90% and reached important physiological events, such as seed coat cracking and radicle extension earlier than seeds from Boyaca and Cundinamarca. In the nursery stage, fruit size significantly affected the growth variables, such as plant height and leaf length, in all the different seed origins. Seedlings from Boyaca showed better results for growth and development after transplanting, as indicated by a higher amount of leaves and presence of basal branching, followed by seedlings from Cauca, Cundinamarca and Antioquia, which showed delayed phenological events.
Key words: mortiño, seeds, promising Andean species, blueberry.
RESUMEN
La propagación del agraz se realiza a partir de plantas silvestres y existe una ausencia de información de la potencialidad agronómica de los materiales disponibles para la propagación sexual, por lo cual se hace necesario desarrollar un protocolo de propagación que permita disponer de información técnica para los productores. El presente estudio tuvo como objetivo evaluar el efecto del tamaño del fruto del que se extrajo la semilla sobre el crecimiento y desarrollo de plántulas de agraz, durante la etapa de germinación y semillero, a partir de frutos provenientes de cuatro áreas locales de las zonas alto-andinas de Antioquia, Cauca, Cundinamarca y Boyacá. En la fase de germinación se obtuvo un mejor comportamiento para las semillas de Antioquia y Cauca las cuales presentaron porcentajes de germinación cercanos al 90%, y alcanzaron eventos fenológicos importantes como el rompimiento de la testa y la extensión de la radícula de manera más temprana con respecto a las semillas provenientes de Cundinamarca y Boyacá. Mientras en la etapa de semillero se observó que el tamaño del fruto afecta significativamente las variables de crecimiento como la altura de la plántula y la longitud del limbo, para todos los materiales estudiados. Las plántulas provenientes de Boyacá presentaron los mejores resultados de crecimiento y desarrollo después del trasplante, reflejado en mayor número de hojas y evidencia de ramificación basal, seguido por las plántulas de Cauca, Cundinamarca y Antioquia las cuales presentaron eventos fenológicos más tardíos.
Palabras clave: mortiño, semillas, especie promisoria de los Andes, arándano.
Introduction
The Andean blueberry plant (Vaccinium meridionale Swartz) belongs to the Ericaceae family, Vaccinioideae subfamily, and Vaccinieae tribe (Abreu et al., 2008). The Vaccinium genus contains about 300 species which are spread mainly over the northern hemisphere and tropical high mountain areas (Ortega, 2011), mostly under wild conditions. In the high Andean areas of South America, several species are reported: V. floribundum, V. meridionale and V. corymbodendrum, found from Venezuela to Bolivia at altitudes between 2,600 and 4,000 m a.s.l. (Patiño and Ligarreto, 2006; Gaviria et al., 2009).
Based on the region it comes from, V. meridionale changes its name. In Ecuador, it is known as "manzanilla del cerro", "raspadura quemada", and the Andean blueberry and, in Colombia, as "agraz", "mortiño", "uvito de monte" and "arándano azul" (Ligarreto, 2009; Ortega, 2011). The berries are collected by extractive harvesting in forests, causing damage to the natural populations due to the high demand for the fruit (Gaviria et al., 2009).
Due to the fact that this fruit is found in the wild in the Andes, it is necessary to know the genetic variability of the species in order to establish management of the populations and to make it a cultivable species in the near future (CEF, 2005). For the same reason, it is important to know all the agronomic features and adaptability of the taxon that is found in the different geographic regions of Colombia.
According to Lopera et al. (2009), the Andean blueberry is considered a promising fruit, which can produce several products with added values; due to its properties, it can be considered a nutraceutic food because it is rich in polyphenolic compounds that have the property of being colorants and antioxidants, potentially health protectors (Gaviria et al., 2009). On the national scale, its consumption is concentrated in the provinces of Antioquia, Boyaca and Cundinamarca. Nowadays, it has high potential for nationwide consumption and is included in the list of species with international marketability (Ligarreto, 2009). Due to these factors, interest for this product has increased as well as interest in establishing productive crops and supplying internal and export demand (Medina et al., 2009).
Each berry produces seeds with different sizes, numbers, colors and shapes; in some fruits, few well developed seeds and several rudiments with a whitish appearance are found (De Valencia and Ramírez, 1993). Andean blueberry seeds are very tiny, from 0.84 to 1.87 mm in length and 0.5 to 1.4 mm wide, with a mainly golden-stained color and some darker variations or reddish color. De Valencia and Ramírez (1993) suggested that the germination rate can vary due to the size of the seed, which was confirmed by the study of Contreras (2010), who found that big blueberry fruits (V. corymbosum) show a higher amount of viable seeds compared to berries of smaller-sized berries. Buseta (1997) suggested that, for blueberries, there is a correlation between size of the fruit and seed content, meaning bigger fruits have a higher amount of seeds.
Laynez et al. (2007) stated that seed size seems to be an important characteristic linked to the growth process of the seedlings and the success of the growth is determined by the physiological and biochemical characteristics of the seed, its reaction to the environment and the rate at which it uses its reserves for starting and sustaining growth in the earlier stages of development before it becomes an autotrophic organism able to process solar energy; the bigger seeds are the ones that are supposed have an advantage for emerging seedlings because they have better metabolic reserves available during the early stages of development, when the plant is not able carry out photosynthesis (Soltani et al., 2006; Quero et al., 2007). Based on the latter aspects in consideration, the main objective of this article was to evaluate the incidence of the size of the fruits from which the seeds were extracted on the growth and development of Andean blueberry seedlings during the germination and nursery stages, using seeds from four local areas of the Colombian Andean region.
Materials and methods
The present study was carried out at the Genetic Resources Laboratory of the Faculty of Agronomy, of the National University of Colombia, Bogota campus. For this purpose, the fruits were collected from four local areas of the following provinces: Antioquia, Cauca, Cundinamarca and Boyaca. The agraz berries of the Antioquia region came from the municipality of Santa Rosa, located at 06°08'06" N and 75°25'03" W, at an altitude of 2,550 m a.s.l.; berries from the Cauca region were collected from the municipality of Purace, located at 02°20'53" N and 76°30'03" W, at an altitude of 2,850 m a.s.l.; berries from the Boyaca region were collected from the municipality of San Miguel de Sema, from a plantation located at 05°31'15" N and 73°43'39" W, at an altitude of 2,615 m a.s.l. and the berries from the Cundinamarca area were collected from a location at 05°23'04" N and 73°41'25" W, at an altitude of 2,676 m a.s.l. in the municipality of Guacheta.
For the four collection areas, the berries were found in ripening stage number 5, corresponding to a dark purple color (L*=20,831, a*=4,026, b*=1,14), based on the classification system purposed by Rincón et al. (2012). The fruits were graded by size into two classes: the first one having berries with a diameter bigger than or equal to 1 cm and the second having berries with diameters smaller than 1 cm, establishing a midpoint based on earlier studies that showed that the fruit size variation was between 0.5 and 1.6 cm (Ligarreto, 2009; Hernández et al., 2009). From each berry, the seeds were extracted through mechanical pressure and water washing and dried under shade at 17°C and 78% RH for 24 h (Hernández et al., 2009).
Two phases were defined for this study: germination and seedling development after transplanting. The study was divided in two phases due to the differences in the substrate in each phase. For the first, the seeds were planted in four replications, two fruit sizes, from four locations and the second phase was ran with a complete randomized design with a 2 x 4 factorial arrangement, where the first factor was seeds from two different sizes of fruit and the second factor was berries from four different locations, each treatment had four replications, for a total of 32 experimental units, each plot formed by 18 seedlings.
For the germination phase, daily observations and measurements were taken from day 0, when the seeds were planted in Petri dishes on absorbent paper and constant water spraying for stimulate imbibitions until day 50, under conditions of 20°C with natural light (8 h) as suggested by Duran et al. (2008). In this phase, the germination rate was measured at 50 days after planting (dap), the cracking of the seed coat and presence of cotyledons, measured based on the general BBCH scale created by Hack et al. (1992), which suggest a phenological identification in 10 main stages of growth for all monocotyledon and dicotyledonous species (Meier, 2001). To evaluate the cracking of the seed coat of the blueberry seeds, growth stage number 0 was used, which corresponded to germination, shout and development of the young leaves.
The second phase started with the end of the germination stage at 50 dap, that is, from the moment of transplanting of the seedling from Petri dishes to substrate plots. The substrate used for transplantation was composed of 70% ground mulch, 10% dry rice husks, 10% carbon ash and 10% sterile substrate (sphagnum peat moss). Measurements started at 103 dap due to a settle down period for adaptation after transplanting; later, measurements were taken at 108, 117, 124, 138, 145, 152, 159, 169, 176, 183, 190 and 201 dap. The measured variables included: plant height, leaf length, leaf number, and phenological stage of the seedlings, based on the BBCH general scale. The interaction between fruit size and seed origin was measured only on the last measurement date due to the fact that it was on that date that the main objectives of the study were evidenced, given the slow growth of the species.
Data processing was done with analysis of variance, Tukey test, considering whether the differences between two averages of each date of sampling were bigger than the LSD (Least Significant Difference) with a difference of P≤0.05 and a Least Square Means test to evaluate the interaction between factors in the purposed design. The software used for this study was SAS v. 9.2.
Results and discussion
Germination phase
Germination rate: At 50 d after the blueberry seeds were conditioned to germination, no significant differences were observed between the germination rates of seeds from fruits of different sizes. However, there was a significant interaction between the seeds from the different fruit sizes and the location (Fig. 1). This can be explained by the fact that each fruit produces seeds that vary in size, amount, color and shape; some fruits contain few well developed seeds and several whitening rudiments (De Valencia and Ramírez, 1993).
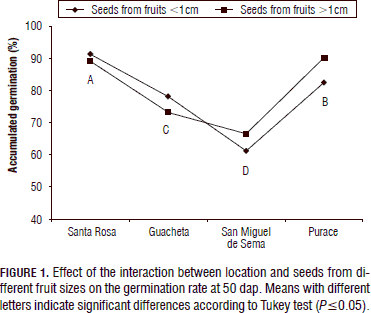
There was a significant effect of the origin of the seeds on the germination rate (Tab. 1); the means comparing test showed that the seeds from the Antioquia location presented the higher germination rates in smaller fruits compared to the other geographic regions, with a mean of 91.40%, followed by the Cauca, Cundinamarca and Boyaca locations, which were significantly different from each other in the same order. The germination rate in Santa Rosa (Antioquia) in this study was higher than the one recorded by Hernández et al. (2009) who observed 63% after 30 d for berries in the complete ripening stage of maturity without applying any treatment over the seed. For San Miguel de Sema (Boyaca), Buitrago and Rincón (2011) recorded 20% germination under the same environmental conditions and the same stages of maturity, very low compared with the rate seen in the present study, 61.33%. The percentage of germination for the Andean blueberry was similar to that reported in previous studies with light and a temperature of 18 to 22ºC with in vitro germination (Castro et al., 2012).
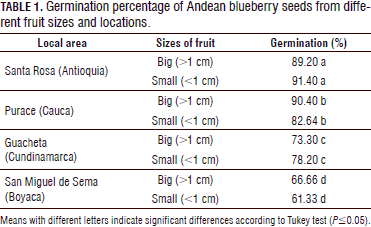
Large berries with diameters higher than or equal to 1 cm showed better results in the seeds from Purace with a germination percentage of 90.40%. In the municipalites of San Miguel de Sema and Guacheta, a lower germination rate was found, as compared to the other regions; perhaps these values were affected by the storage periods of the seeds.
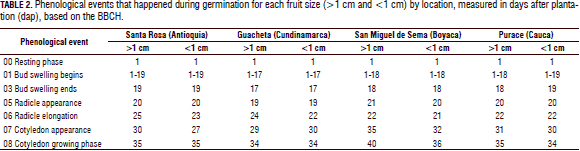
Cracking of seed coat: In Tab. 2, the subdivision of growth stage 0 is seen, based on the BBCH, the phenological event 5 radicle emergence was evidenced between 19 and 21 dap, which matches with previous studies carried out by Buitrago and Rincón (2011), who found that Andean blueberry seeds show germination at an average of 20 dap, and Castro et al. (2012), who indicated that germination begins at 14 d and ends at day 56.
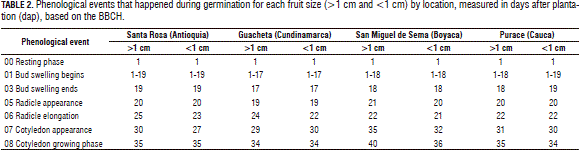
Cotyledon apparition: was evaluated using the BBCH scale with phenological event 7, which suggests the showing up of the cotyledons, as observed in Tab. 2, seeds from the smaller berries of the Santa Rosa location showed an earlier appearance of cotyledonal leaves when compared to the other treatments, with an 8 d difference from the most delayed seeds of San Miguel de Sema. These results are different from the studies of Buitrago and Rincón (2011), who found that seeds from Boyaca show entirely spread up cotyledons 30 d after planting under the same environmental conditions of the current study.
In previous studies done by De Valencia and Ramírez (1993), blueberry seeds from Cundinamarca showed cracking of the seed coat and appearance of cotyledonal leaves at 35 dap, which is different from the seeds of Guacheta in the current study, which showed cotyledonal leaves at 30 dap. These differences in germination rates may be due to differences in altitude and temperature between both studies and the environmental conditions where the seeds develop in natural conditions in each region. Baskin et al. (2000) reported that, in some studies made with several taxa of Vaccinium, under constant light conditions but varying temperatures, changes of at least 5°C can have a strong influence on the germination rate as well as on the rate of seedling growth (Hernández, 2008).
Development phase
Plant height: this variable showed significant differences for the size of the fruit from which the seeds were extracted, seedlings from higher diameter berries showed a difference of 5.7 mm on the last measurement date (201 dap) as compared to the seedlings of seeds extracted from smaller diameter fruits, as observed in Figs. 2 and 3A. This result coincides with Cordazzo (2002), who suggested that larger size seeds generate bigger plants, when compared with those generated by smaller seeds. Seedlings from Purace showed an increasing significant difference in growth to the other regions, as observed at 117 dap; this was due to the fact that the berries from Purace were larger, as compared with the other berries. As reported by Ligarreto et al. (2011), big fruits contain more seeds of a larger size when compared with small berries. Generally, the size of the seed has an influence on the shape of the plant, that is, smaller seeds generate shorter roots and a greater susceptibility to fungi (De Valencia and Ramírez, 1993).
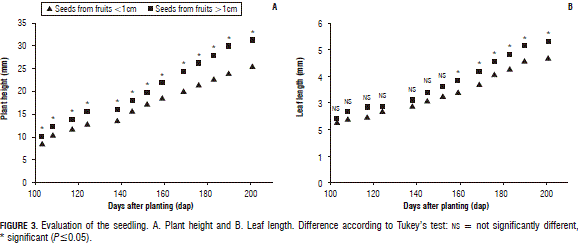
The seedlings from Santa Rosa showed the lowest values after 152 dap, reaching a difference of 9.64 mm when compared with the Purace seedlings on the last measurement date (Fig. 4A); however, during the first three measurement dates, they presented a higher height, as compared to the other regions. After 124 dap, the height of those seedlings presented a non-evident increase until the end of the study. No significant differences were found in the plant height between seedlings from Guacheta and San Miguel de Sema during the whole phase, but a significant interaction was found on the last measurement date, as shown in Fig. 2, being highly significant for the seedlings from Guacheta where berries with diameters higher than or equal to 1 cm showed a better response when compared to the seedlings from San Miguel de Sema which showed a significant interaction, at a lower proportion. However, in seedlings from Purace and Santa Rosa interactions were not significant. Fig. 2 shows that berries with higher diameters from Guacheta, San Miguel de Sema and Purace produced the larger seedlings.
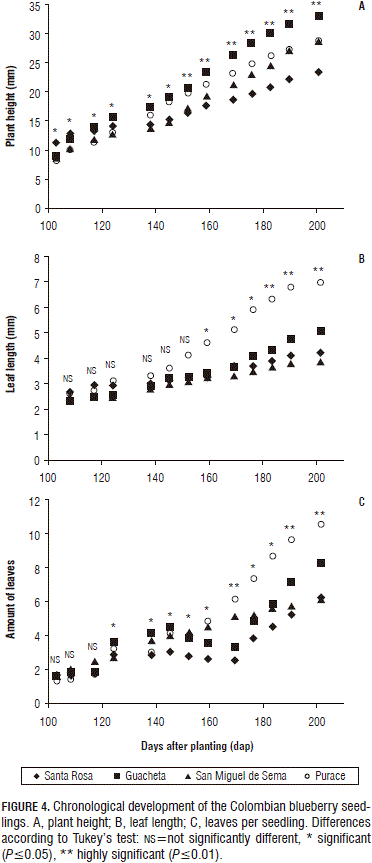
On average, seedlings in the current study showed an approximate growth of 28.51 mm in the shoots during a 201 d period, which suggests a very slow growth of the species, this is confirmed by Calderón and Socha (2009), who reported that, due to the slow growth of V. meridionale, it is difficult to research. This growth characteristic, is generalized for Vaccinium, Sierra et al. (2005), described V. corymbodendron as a slow growth plant for the early stages of development.
Leaf length: no significant differences were found for this variable until 159 dap, after this measurement date, seedlings from berries with diameters smaller than 1 cm showed lower values when compared with seedlings from higher diameter berries, as shown in Fig. 3B.
During the six earlier measurement dates, no significant differences were found between the different locations of the seeds. Seedlings from San Miguel de Sema showed an increasing growth from 145 dap to the end of the study, gaining an elongation of the leaf axis of 7 mm. Seedlings from Purace showed a growth of 5.06 mm, being statistically different from the seedlings from Santa Rosa and Guacheta, which showed values of 4.25 and 3.87 mm, respectively (Fig. 4B). A higher leaf length can be related to a greater leaf area, which can increase its capacity to intercept more sunshine and carry out more photosynthesis.
In most plants, morphological characteristics stimulated by environmental changes include stem elongation and leaf growth (Fordyce, 2006), this fact is appreciable in the blueberry seedlings from the four locations of this study and is even more evident in the seedlings of seeds from San Miguel de Sema which presented better growth characteristics for all variables except plant height, probably due to a better adaptation to the light conditions, seed storage, temperatures and state of fruit maturity in the current study.
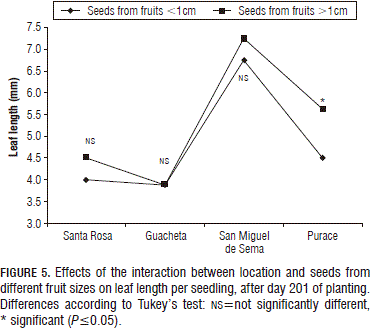
In respect to the interaction of the factors on the last measurement date, the size of the fruit in seedlings from the Purace location was statistically significant, as observed in Fig. 5; however, this was not very representative due to the fact that the seedlings from San Miguel de Sema showed the longest leaf lengths despite the berry-size origin of the seeds.
Leaves per seedling: no significant differences were found regarding the size of the fruits from which the seeds were extracted; however, different locations showed some statistically significant differences starting on the third measurement date (Fig. 4C).
The study showed that, until 145 dap, there was a very slight increase in the amount of leaves for all treatments; however, at 152 dap, some stress symptoms were observed which were evidenced by yellowing of the young leaves due to an approximated increase of the temperature of 3°C, caused by a lack of aeration; the seedlings from Guacheta seemed not to be affected, whereas the seedlings from San Miguel de Sema showed a slight impact due to the temperature increase, which caused a regeneration of the seedlings and basal branching. This event caused an increase in the amount of leaves and a better adaptation to adverse environmental conditions. Statistically, the seedlings from San Miguel de Sema showed important differences when compared to the other regions, reaching values of 11 leaves per plant, followed by seedlings from Purace with 8 leaves per plant, Santa Rosa and Guacheta, with six leaves per plant.
Starting at 145 until 169 dap, the seedlings from Santa Rosa showed a decrease in the amount of leaves; this fact was evidenced by a decrease in growth in each of the evaluated variables due to the temperature stress found in the environment during the study. Starting at 169 dap, the seedlings from Purace and Santa Rosa showed an accelerated increase in the amount of leaves, as shown in Fig. 4C; however, they didn't show any basal branching. The interaction between the factors was not significant for this variable during the study.
Conclusions
The seedlings of the seeds from the berries of Santa Rosa and Purace showed high germination rates, with values higher than 80%. The seedlings from San Miguel de Sema, presented basal branching and a better result in the variables evaluated during the growth phase.
Agraz berries with diameters higher than or equal to 1 cm favor high quality seeds, which in turn generate vigorous seedlings in terms of height and leaf length; for this reason, using them can constitute a good practice to ensure the growth and development of the species during the nursery-propagation phase.
Literature cited
Abreu, O., A. Cuéllar, and S. Prieto. 2008. Fitoquímica del género Vaccinium (Ericaceae). Rev. Cubana Plant Med. 13(3), 1-11. [ Links ]
Baskin, C., P. Milberg, L. Andersson, and J.M. Baskin. 2000. Germination studies of three dwarf shrubs (Vaccinium, Ericaceae) of northern hemisphere coniferous forests. Can. J. Bot. 78(12), 1552-1560. [ Links ]
Buitrago, C.M., and M.C. Rincón. 2011. Determinación de los estados de madurez y evaluación del comportamiento del fruto de agraz (Vaccinium meridionale Swartz) bajo refrigeración, Undergraduate thesis. Faculty of Agronomy, Universidad Nacional de Colombia, Bogota. [ Links ]
Buseta, A. 1997. Chile: berries para el 2000. Departamento Agroindustrial, Fundación Chile, Santiago. [ Links ]
Calderón, L.C. and A.C. Socha. 2009. Evaluación del estado fitopatológico del agraz (Vaccinium meridionale Sw.) en localidades del altiplano cundiboyacense (dos de Boyacá y una de Cundinamarca). Undergraduate thesis. Faculty of Basic Sciences, Universidad Militar Nueva Granada, Bogota. [ Links ]
Castro, C., Y. Olarte, L. Rache, and J. Pacheco. 2012. Development of a germination protocol for blueberry seeds (Vaccinium meridionale Swartz). Agron. Colomb. 30(2), 196-203. [ Links ]
CEF, Centro de Excelencia Fitosanitaria. 2005. Distribución de agraz o mortiño (Vaccinium spp.) en Antioquia. Newsletter CEF No. 9. In: www.ica.gov.co/CEF/boletines/boletin09.htm; consulted: November, 2013. [ Links ]
Contreras, M. 2010. Efecto de la aplicación de CPPU sobre la calidad de fruta en arandano alto (Vaccinium corymbosum L.) cultivar Elliott. Undergraduate thesis. Faculty of Agricultural and Forestry Sciences, Universidad de la Frontera, Temuco, Chile. [ Links ]
Cordazzo, C.V. 2002. Effect of seed mass on germination and growth in three dominant species in southern Brazilian coastal dunes. Braz. J. Biol. 62(3), 427-435. [ Links ]
De Valencia, M. and F. Ramírez. 1993. Notas sobre la morfología, anatomía y germinación del agraz (Vaccinium meridionale Swartz). Agron. Colomb. 10(2), 151-159. [ Links ]
Duran, S., V. Molina, D. Uzcategui, C. Molina, and K. Yatzury. 2008. Factores que determinan la germinación del Vaccinium meridionale Swartz (chivacu) del páramo la culata. Rev. Cient. Juvenil 12, 19-24. [ Links ]
Fordyce, J.A. 2006. The evolutionary consequences of ecological interactions mediated through phenotypic plasticity. J. Exp. Biol. 209, 2377-2383. [ Links ]
Gaviria, M.C., C. Ochoa, N. Sánchez, C. Medina, M. Lobo, P. Galeano, A. Mosquera, A. Tamayo, Y. Lopera, and B. Rojano. 2009. Actividad antioxidante e inhibición de la peroxidación lipídica de extractos de frutos de mortiño (Vaccinium meridionale Swartz). Bol. Latinoam. Caribe Plant. Med. Aromát. 8(6), 519-528. [ Links ]
Hack, H., H. Bleiholder, L. Buhr, U. Meier, U. Schnock-Fricke, E. Weber, and A. Witzenberger. 1992. Einheitliche Codierung der phänologischen Entwicklungsstadien mono â und dikotyler Pflanzen â erweiterte BBCH-Skala, allgemein. Nachrichtenblatt. Deut. Pflanzenschutzd. 44, 265-270. [ Links ]
Hernández, M.I. 2008. Estudio morfo-fisiológico de la semilla de mortiño (Vaccinium meridionale Swartz). M.Sc. thesis. Postgraduate School, Universidad Nacional de Colombia, Medellin, Colombia. [ Links ]
Hernández, M., M. Lobo, C. Medina, J. Cartagena, and O. Delgado. 2009. Comportamiento de la germinación y categorización de la latencia en semillas de mortiño (Vaccinium meridionale Swartz). Agron. Colomb. 27(1), 15-23. [ Links ]
Laynez, J.A., J.R. Méndez, and J.M. Figueroa. 2007. Influencia del tamaño de la semilla de maíz (Zea mays L.) en el crecimiento de plántula en condiciones de salinidad. Idesia 25(2), 23-35. [ Links ]
Ligarreto, G. 2009. Descripción del género Vaccinium, estudio de caso: agraz o mortiño (Vaccinium meridionale Swartz). pp. 13- 27. In: Ligarreto, G. (ed.). Perspectivas del cultivo de agraz o mortiño (Vaccinium meridionale Swartz) en la zona altoandina de Colombia. Universidad Nacional de Colombia, Bogota. [ Links ]
Ligarreto, G., M. Patiño, and S. Magnitskiy. 2011. Phenotypic plasticity of Vaccinium meridionale (Ericaceae) in wild populations of mountain forests in Colombia. Rev. Biol. Trop. 59(2), 569-583. [ Links ]
Lopera, Y., C. Gaviria, and B. Rojano. 2009. Fermentación alcohólica del zumo de mortiño (Vaccinium meridionale Swartz) y determinación de la capacidad antioxidante. p. 105. In: XIII Congreso Nacional de Biotecnologia y Bioingenieria. Acapulco, Mexico. [ Links ]
Medina, C., M. Lobo, M.P. Patiño, G. Ligarreto, O. Delgado, S. Lopera, and J. Toro. 2009. Variabilidad morfológica en agraz en la zona altoandina de Colombia. pp. 57-74. In: Ligarreto, G. (ed.). Perspectivas del cultivo de agraz o mortiño (Vaccinium meridionale Swartz) en la zona altoandina de Colombia. Universidad Nacional de Colombia, Bogota. [ Links ]
Meier, U. 2001. Estadios de las plantas mono y dicotiledóneas. BBCH Monografia. 2nd ed. Centro Federal de Investigaciones Biológicas para Agricultura y Silvicultura (BBCH), Limburgerhof, Germany. [ Links ]
Ortega, H. 2011. Desarrollo de un vino de mortiño (Arandanos) en la Corporación Grupo Salinas de Ecuador. Undergraduate thesis. Universidad Pública de Navarra, Madrid. [ Links ]
Patiño, M.P. and G.A. Ligarreto. 2006. Caracterización morfológica in situ de poblaciones espontáneas de Vaccinium sp. en los departamentos de Boyacá, Cundinamarca y Nariño. p. 112. In: Memorias 1er Congreso Colombiano de Horticultura. Sociedad Colombiana de Ciencias Hortícolas, Bogota. [ Links ]
Quero, J.L., R. Villar., T. Marañon., R. Zamora, and L. Poorter. 2007. Seedmass effects in four Mediterranean Quercus species (Fagaceae) growing in contrasting light environments. J. Bot. 94(11), 1795-1803. [ Links ]
Rincón, M.C., C.M. Buitrago, G.A. Ligarreto, W.S. Torres, and H.E. Balaguera. 2012. Comportamiento del fruto de "agraz" (Vaccinium meridionale Swartz) cosechado en diferentes estados de madurez y almacenado en refrigeración. Rev. Fac. Nal. Agron. Medellin 65(2), 6615-6625. [ Links ]
Sierra, J., F. Alzate, H. Soto, B. Duran, and L. Losada. 2005. Plantas silvestres con potencialidad ornamental de los bosques montano bajos del oriente Antioqueño, Colombia. Rev. Fac. Nal. Agron. Medellin 58(1), 2651-2663. [ Links ]
Soltani, A., M.J. Robertson, B. Torabi, M. Yousefi-Daz, and R. Sarparast. 2006. Modelling seedling emergence in chickpea as influenced by temperature and sowing depth. Agric. Forest Meteorol. 138, 156-167. [ Links ]
