










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. vol.32 no.2 Bogotá maio/ago. 2014
https://doi.org/10.15446/agron.colomb.v32n2.43861
http://dx.doi.org/10.15446/agron.colomb.v32n2.43861
PROPAGATION AND TISSUE CULTURE
1 Research Group Andean Fruit Production, Faculty of Agricultural Sciences, Universidad de Nariño. Pasto (Colombia). hcriolloescobar@gmail.com
2 Miembro de Número, Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Bogota (Colombia).
3 Plant Tissue Laboratory, Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario (IMIDRA). Madrid (Spain).
4 Research Group Andean Fruit Production, Universidad de Nariño. Pasto (Colombia).
Received for publication: 4 May, 2014. Accepted for publication: 30 July, 2014.
ABSTRACT
Lulo is a species of great importance to the fruticulture of Colombia, but has significant phytosanitary problems that require an aggressive breeding program oriented toward the production of genotypes with tolerance to phytopathogens. These programs need to establish highly efficient mass plant propagation protocols, such as somatic embryogenesis. This study focused on research on the somatic embryogenesis of lulo using kinetin, naphthalene acetic acid-NAA (Plant Growth Regulators, PGRs), and different sucrose concentrations in a MS medium. Two lulo varieties, Solanum quitoense var. septentrionale and S. quitoense var. quitoense, and two explant types (hypocotyl and cotyledon) were used, incubated in dark conditions at 25±2°C. The highest production percentage of the embryos was obtained when 50 mM of NAA were added to the medium with sucrose (50.0 and 263.1 mM) for the two explant types used. In lulo with spines, the highest percentage of embryonic structures (50%) was observed with cotyledonary leaf explants and 50 mM of NAA ; while in the spineless lulo, the embryonic structures were observed in the same type of explant with 50 mM of NAA + 263.1 mM of sucrose (32%).
Key words: embryo culture, recalcitrance, plant growth regulators, tissue culture, plant propagation, fruit.
RESUMEN
El lulo es una especie de gran importancia en la fruticultura colombiana, con grandes problemas fitosanitarios que requieren un agresivo programa de mejoramiento orientado a la producción de variedades tolerantes. Estos programas necesitan establecer protocolos de propagación masiva de plantas de alta eficiencia como la embriogénesis somática. Nuestras investigaciones se orientaron al estudio de la embriogénesis somática de lulo, mediante la interacción de los fitorreguladores kinetina y ácido naftalenacético (ANA), y diferentes concentraciones de sacarosa en medio MS . Se utilizaron dos variedades de lulo (Solanum quitoense var. septentrionale y S. quitoense var. quitoense), dos tipos de explantes (hojas cotiledonares e hipocotilos), incubados en condiciones de oscuridad, a 25±2°C. El mayor porcentaje de producción de embriones se obtuvo cuando se adicionaron al medio 50 mM de ANA además de sacarosa (50,0 y 263,1 mM), para los dos tipos de explantes utilizados. En lulo con espinas, el mayor porcentaje de estructuras embrionarias (50%) se observó con hojas cotiledonares y 50 mM de NAA, mientras que en lulo sin espinas estos se observaron en el mismo tipo de explante con 50 mM NAA + 263,1 mM sacarosa (32%).
Palabras clave: cultivo de embriones, recalcitrancia, reguladoras del crecimiento vegetal, cultivo de tejidos, propagación de plantas, fruta.
Introduction
Lulo (Solanum quitoense Lam.) originates from the Andes and has great potential for the international markets. Cultivation in Colombia is carried out in marginal coffee regions, in small areas with low technology and with numerous phytosanitary problems. It is a species that is highly susceptible to diseases and pests that cause large losses for producers, which demonstrates the need to create breeding programs with tolerance characteristics, before starting an ambitious exportation program.
The plant breeding programs require efficient methods of plant propagation that allow the mass propagation of selected genotypes. Despite many difficulties, many authors, such as Chandel and Katiyar (2000), Rout et al. (2006), George and Debergh (2008), Oláh et al. (2009), and Parimalan et al. (2011) consider somatic embryogenesis to be the most efficient method, especially when it is necessary to regenerate plants using genetically modified cells, as expressed by Nakagawa et al. (2001), and Van Der Vyver (2010).
Rout et al. (2006) stressed that the regeneration of plants through somatic embryogenesis has big advantages, such as high multiplication rates that can be used in bioreactors, the possibility of encapsulating the embryos and producing artificial seed, cryoconservation, genetic fidelity in the offspring and in vitro selection of tolerant plants. Similarly, Campbell et al. (2003) considered these processes as valuable complements to the regeneration of genetically transformed plants. However, the principal limitation is related to the genotypic dependence of the somatic embryonic production and its low germination power (Wang et al., 2006), considering that the low germination power is not in all cases a generalized limitation of somatic embryogenesis. Another significant limitation in addition to the genotypic dependence is that only a few species have protocols that are sufficiently refined for mass somatic embryonic multiplication.
The use of somatic embryogenesis and the regeneration of plants have been studied for several cultivated species, with differing results. For example, the research of Wang et al. (2006) on the regeneration of recalcitrant cotton genotypes, of Nakagawa et al. (2001) and Ren et al. (2012) on melon using cotyledonal tissue, and on citrus, Cardoso et al. (2012) and Nakano et al. (2013), and the cacao, Tan and Furtek (2003) and Quainoo et al. (2008); there have been several studies on the somatic embryogenesis of cultivated plants, such as, strawberry (Zhang et al., 2014), onion (Ramakrishnan et al., 2013), carrot (Li et al., 2013), sweet potato (Santa-Maria et al., 2009), chickpea (Mishra et al., 2012), broccoli and cabbage (Qin et al., 2007; Yang et al., 2010; Na et al., 2011), and groundnut (Singh and Hazra, 2009).
In the Solanaceae family, despite being considered a recalcitrant species for somatic embryogenesis, successful protocols have been produced by Kaparakis and Alderson (2002), Kantharajah and Golegaonkar (2004), Mir et al. (2008), and Ribeiro et al. (2009) for Solanum melongena; JayaSree et al. (2001), Sánchez-Enciso et al. (2005), Vargas et al. (2005), and Motallebi-Azar et al. (2013) for Solanum tuberosum; Kintzios et al. (2000), Bodhipadma and Leung (2002), Bárány et al. (2005), Koleva-Gudeva et al. (2007), Kaparakis and Alderson (2008), and Solís-Ramos et al. (2010) for Capsicum annuum; Chandel and Katiyar (2000) and Vinoth et al. (2014) for Solanum lycopersicum; Correia and Canhoto (2009) and Arahana et al. (2010) for Cyphomandra betacea; Dhavala et al. (2009) for Solanum trilobatum; Zapata-Castillo et al. (2007) for Capsicum chinense. However, there have been no reports for somatic embryogenesis in lulo (S. quitoense) and the attempts carried out by Segovia (2002) using a cultivation medium appropriate for C. betacea failed.
The present study aimed to evaluate the effect of different hormone combinations, explants and incubation conditions on the induction of somatic embryogenesis of two botanical varieties of lulo, S. quitoense var. septentrionale (with spines) and S. quitoense var. quitoense (without spines).
Materials and methods
This study was carried out in the Plant Tissue Laboratory of the Biology Program, Faculty of Science, Universidad Nacional de Colombia, Bogota and in the Plant Tissue Laboratory of the Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario (IMIDRA-Spain).
The donor plants were obtained by germinating the seeds extracted of fruits of one lulo plant with spines and one lulo plant without spines, sterilized using soap, sterile water and 15 min in 1.5% sodium hypochlorite; followed by five washings with sterile water and transfer to a MS culture medium (Murashige and Skoog, 1962) in petri dishes and germination trays.
After 30 d, hypocotyl and cotyledonary leaves were taken as explants from the lulo with spines and the lulo without spines and planted in a MS medium supplemented with different combinations of PGRs of NAA (19.8, 31.9 and 50.0 mM) as factor A, kinetin (0 and 13.9 mM) as factor B and different sucrose concentrations (14.6, 58.4 and 263.1 mM) as factor C. The explants were kept in dark conditions at 25±2°C. At 30 d, the plants were evaluated and transferred to a MS medium without PGRs, with a 16/8 h photoperiod.
Each botanical variety and each of the experiments were subjected to an unrestrictive random experimental design with a factorial arrangement, with 10 repetitions with four explants each.
A descriptive analysis was carried out on the characteristics of the produced calluses and the presence of roots and calluses was determined for each treatment. The average size of the formed calluses was determined by quantifying each callus using a scale of 1 to 5, determining the average for each experimental unit and transforming them to a percentage with 5 being 100% and 1 the absence of calluses; the embryos and embryo like structures formed in each experimental unit were counted. The experimental data were subjected to AN OVA and a Tukey mean comparison test for simple effects and interactions with P≤0.05.
Results
Effect of the concentrations of NAA , kinetin and sucrose on the growth of the explants of lulo with spines
The use of cotyledonary leaves from lulos with spines determined the interactions: NAA *kinetin and NAA *sucrose, in the formation of roots, and NAA *kinetin and Kinetin*sucrose, in the formation of calluses. The NAA *kinetin interaction was observed in the somatic embryo induction. As simple effects, the three factors significantly influenced the formation of the roots; with kinetin in the formation of the calluses; and NAA and kinetin in the induction of embryos and embryo like structures after 30 d of incubation.
Root induction progressively decreased when the NAA concentration increased from 19.8 and 50.0 mM (Fig. 1a). Kinetin exerted a strong influence in the low NAA doses (19.8 mM), with statistical differences between the highest percentage of rooted explants without the addition of kinetin and that observed with kinetin (13.9 mM); with 31.9 mM of NAA , there was also a significant reduction in the percentage of rooted explants with kinetin applications; however, with the highest NAA dose (50.0 mM), the effect of the kinetin application was not significant. Sucrose concentration behaved differently; with a low concentration of NAA, the largest rooting was observed with 58.4 mM sucrose, while with the average dose of NAA, the highest rooting was obtained with 58.4 mM and 263.1 mM sucrose; with the highest concentration of ANA, the effect of the sugar concentration was not significant (Fig. 1b).
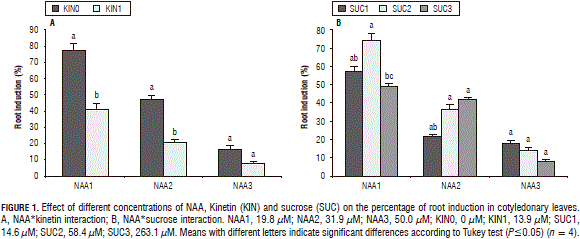
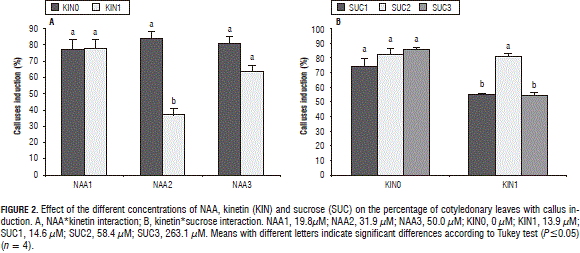
With the low and high NAA concentrations, kinetin did not present a significant effect on the percentage of cotyledonary leaves with formed calluses but, in the medium dose (31.9 mM), the explants that grew in the absence of kinetin presented a higher induction of calluses than those obtained in the medium with kinetin (Fig. 2a). In the kinetin*sucrose interaction, the influence of the sucrose concentration was not significant in the medium without kinetin, while, in the medium with kinetin, the medium sucrose concentration (58.4 mM) induced a higher production of explants with calluses (Fig. 2b).
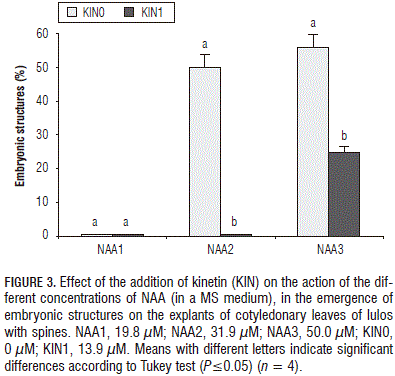
The addition of kinetin to the cultivation medium reduced the induction of somatic embryos when it interacted with NAA ; with 19.8 mM of NAA , with or without kinetin, embryos did not form, while, with 31.9 and 50.0 mM of NAA , embryos were observed in 56 and 50% of the explants, respectively, with significant differences with respect to these NAA treatments supplemented with 13.9 mM of kinetin (Fig. 3).
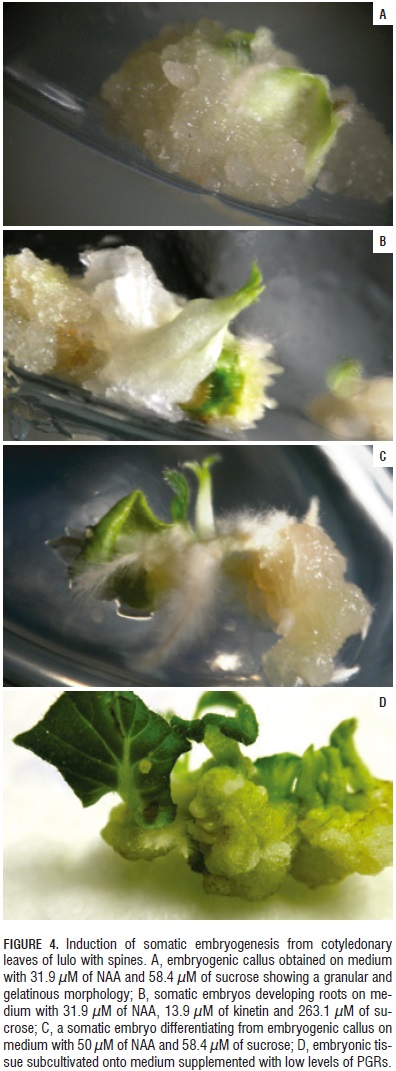
The presence of embryonic structures was characterized by the initial formation of white callus zones in the basal regions of the cotyledonary leaves, calluses of gelatinous textures and granular appearance; the aberrant embryo always started with an enlarged base structure making up the caulinar apex and giving rise to the cotyledonary leaves with a radical system immersed in the callus (Fig. 4).
When the explants were hypocotyls, interactions were observed in the formation of roots between NAA and kinetin, NAA and sucrose and kinetin*sucrose; during the development of embryonic structures, interactions were seen between NAA with kinetin and sucrose and between kinetin and sucrose. The three factors influenced the formation of roots, the sucrose induced calluses and NAA and sucrose induced embryos.
Roots did not present with the use of kinetin, especially with the medium (31.9 mM) and high NAA dose (50.0 mM), while a reduction in rooting was seen with sucrose (263.1 mM) and NAA (19.8 mM) and with sucrose (14.6 mM) and NAA (58.4 mM). The interaction between kinetin and sucrose demonstrated an inhibitory effect on the roots with high concentrations of sucrose and kinetin but with a minor effect when the regulator was not applied.
The formation of calluses in the hypocotyls with the application of PGRs was high and there were only statistical differences between the percentages of explants with calluses, seen in the low (77.9%) and the medium (79.3%) sucrose concentrations, and the percentages of hypocotyls with callus formation, seen in the high concentration of sucrose (52.6%).
Looking for the appearance of embryonic structures evaluated after 30 d of incubation, the higher percentages of explants with embryos (25%) were achieved without kinetin and a medium dose of NAA . The same dose presented a higher percentage of embryos with the medium and high sucrose doses. Similarly, with the cotyledonary leaf explants, the hypocotyl embryos originated in white, bright calluses that had a granular, gelatinous appearance and friable consistency with wide, round bases that surrounded the caulinar apex, submerged in the callus body (Fig. 5).

Effect of NAA , kinetin, and sucrose on the growth of explants of lulo without spines
The results obtained with lulo without spines were very similar to those obtained from lulo with spines. The rooting of the cotyledonary leaves was influenced by the three studied factors and the NAA *sucrose interaction. There were no significant differences between the factors for the formation of the calluses. The induction of embryonic structures was determined by the effect of NAA , kinetin, sucrose and by the NAA *kinetin, NAA *sucrose and kinetin*sucrose interactions.
The percentage of rooted explants was higher without the addition of kinetin. The sucrose concentrations did not affect the rooting percentages with the NAA dose but, with the medium and high doses, the higher concentrations of sucrose significantly reduced the rooting. With the low and medium NAA doses, the inclusion of kinetin reduced the formation of embryonic structures but, with the high dose, the addition of kinetin did not affect the percentage of explants with embryos. Similarly, the positive effect of the higher concentrations of sucrose on the appearance of embryos was obtained without kinetin in the medium (Fig. 6).
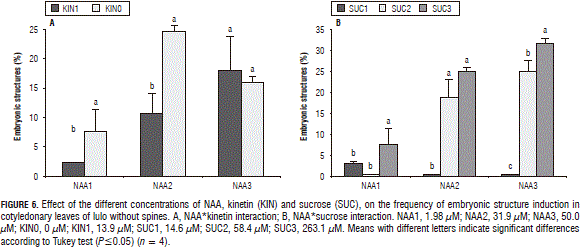
The formation of roots was higher (58.7%) with the hypocotyls of lulo without spines as explants with the lowest NAA concentration (19.8 mM), as compared with the medium (31.9 mM) (34.17%) and high doses (50.0 mM) (24.89%), which did not differ. Similarly, a kinetin and sucrose interaction was observed, presenting a higher inhibitory effect of the sucrose without kinetin in the medium. In the formation of calluses in the explants, marked effects were seen from NAA and kinetin, without an interaction between them. The higher percentages of hypocotyls with the presence of calluses were observed with NAA (19.8 and 31.9 mM) and without kinetin in the medium. Each of the studied factors (NAA , kinetin and sucrose) significantly affected the percentage of somatic embryos that appeared in the medium with interactions between NAA and kinetin and between kinetin and sucrose. In all the NAA concentrations, the addition of kinetin significantly reduced the presence of embryos (Fig. 7a). With the 31.9 and 50 mM of NAA concentrations, the higher percentages of somatic embryos were obtained (22.5 and 17.0%, respectively). The kinetin*sucrose interaction demonstrated a very significant effect on the sucrose when kinetin was not applied, with higher percentages of somatic embryos with 50.1 and 263.1 mM of sucrose in the MS medium (Fig. 7b).

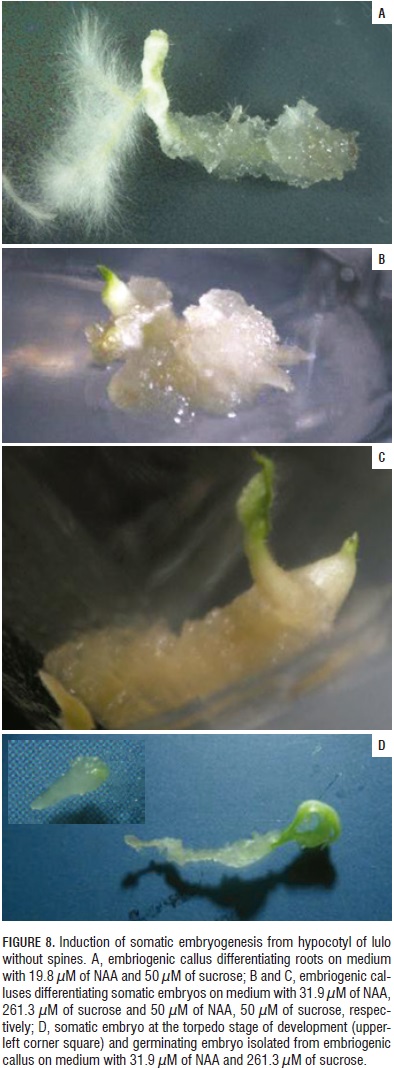
The somatic embryos, as in the lulo with spines, originated from calluses with a friable appearance that were white to creamy and had a granular appearance. The embryos that germinated in the induction medium presented basal widening that culminated in the apex where the emergence of cotyledons started and, in the distal part, the radical grew through the calluses without achieving complete independence during the initial development (Fig. 8). In all the cases in which the calluses were subcultures in a medium without regulators, problems of phenolization or growth detention were seen.
Discussion
Solanaceaes are considered recalcitrant species in studies related to somatic embryogenesis, although, currently, important species of this family have been cloned through embryonic induction, such as eggplant (S. melongena) (Huda and Bari, 2007; Mir et al. 2008; Ribeiro et al. 2009); tomato (S. lycopersicum) (Kaparakis and Alderson, 2002; Vinoth et al., 2014), the potato (S. tuberosum) (JayaSree et al., 2001; Sánchez-Enciso et al., 2005; Vargas et al., 2005; Sharma et al., 2008) and pepper (C. annuum) (Kintzios et al., 2001; Bodhipadma and Leung, 2002; Bárány et al., 2005; Koleva-Gudeva et al., 2007; Solís-Ramos et al., 2010). Despite the importance of lulo (S. quitoense) as a fruit species of high nutritional value, there have been no successful reports on its cloning with somatic embryogenesis; only studies have been carried out by Segovia (2002), who tried without success to regenerate lulo with embryogenic methods.
Levels of NAA , kinetin and sucrose determined the induction rates of roots, callus and somatic embryos in both explants (cotyledonary leaves and hypocotyls) confirming that the addition of kinetin inhibited the rooting action of small NAA doses and reduced the percentage of explants with calluses, which agrees with the findings of Van Staden et al. (2008), who confirmed that high cytokinin concentrations (0.5-10.0 mg L-1) prevented radical growth and the rooting promoting effects of auxins, but is contrary to the report of Thornton et al. (2013), who stated that organogenic processes must be supplemented with auxins and cytokinins.
The appearance of somatic embryos in the induction medium confirmed the recalcitrant characteristic of lulo and the presence of only a few embryonic cells in the differentiated cell medium. These observations agree with Santos and Fevereiro (2002), who stated that cells with an embryogenic capacity can coexist in the same callus with cells with no embryogenic capacity. According to González et al. (2005), embryogenic cells originate in meristematic cells and from their division in specific meristematic tissues, as confirmed by permanent embryonic cells.
Similarly, a positive effect was observed from the sucrose on the appearance of embryogenic structures, possibly due to the fact that, in addition to being an energy source for the process, sucrose acts as an osmotic stress factor that improves the efficiency of somatic embryogenesis (Daigny et al., 1996; Iraqi and Tremblay, 2001; Jheng et al., 2006). Hong et al. (2008) reported that the water stress originating from high sucrose concentrations induces the accumulation of protein and carbohydrates in the cells, which favors the induction of embryogenic cells and inhibits the early germination of embryos in S. melongena. This finding is contrary to the observations of the present study, which confirmed the recalcitrant characteristic of lulo, with embryos appearing sporadically in the calluses that grew in the induction medium without generating the continuation of the recurrent process; moreover, the lack of the embryogenic cell proliferation necessary for the formation of PEMs, despite the high hormone concentrations present in the medium, allowed for the appearance of fewer embryos in the cultured explants.
When the regulators were eliminated from the medium, the calluses experienced necrosis without generating new embryonic structures or other organogenesis processes, as occurs in other species. It is possible that, as observed by Arahana et al. (2010) in studies on the somatic embryogenesis of S. betacea, calluses with embryogenic potential do not form embryos due to a reduction in the cellular maturation due to a lack of sucrose at suitable concentrations, which allows for the induction of nutrient accumulation in embryogenic cells.
Conclusions
Lulo can be considered a recalcitrant species for the induction of somatic embryogenesis that does not respond suitably to protocols that are successful for other Solanaceae species. In dark conditions at 25±2°C with a MS medium and young lulo explants, with or without spines (hypocotyls and cotyledonary leaves), it was possible, for the first time, to obtain germinated embryos in the induction medium with additions of NAA (31.9 and 50 mM) and sucrose (50.0 and 263.1 mM), 30 d after sowing, that are easily reproduced in a MS medium.
The elimination of PGRs from the cultivation medium caused a reduction in growth and necrosis in the calluses, completely eliminating the possibility of obtaining a recurrent embryogenic process.
Acknowledgements
The authors express their gratitude to the Universidad Nacional de Colombia, the Universidad de Nariño, the Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario (IMIDRA).
Literature cited
Arahana B., V., A.R. Cabrera V., and M.L. Torres P. 2010. Propagación de tomate de árbol (Solanum betaceum) vía embriogénesis somática. Avances 2, B16-B21. [ Links ]
Bárány, I., P. González-Melendi, B. Fadón, J. Mitykó, M.C. Risueño, and P.S. Testillano. 2005. Microspore-derived embryogenesis in pepper (Capsicum annuum L.): subcellular rearrangements through development. Biol. Cell 97, 709-722. [ Links ]
Bodhipadma, K. and D.W.M. Leung. 2002. Factors important for somatic embryogenesis in zygotic embryo explants of Capsicum annuum L. J. Plant Biol. 45, 49-55. [ Links ]
Campbell, M.M., A.M. Brunner, H.M. Jones, and S.H. Strauss. 2003. Forestry's fertile crescent: the application of biotechnology to forest trees. Plant Biotechnol. J. 1, 141-154. [ Links ]
Cardoso, J.C., A.P. Martinelli, and R.R. Latado. 2012. Somatic embryogenesis from ovaries of sweet orange cv. Tobias. Plant Cell Tiss. Organ Cult. 109, 171-177. [ Links ]
Chandel, G. and S.K. Katiyar. 2000. Organogenesis and somatic embryogenesis in tomato (Lycopersicon esculantum Mill.). Adv. Plant Sci. 13, 11-17. [ Links ]
Correia, S. and J. Canhoto. 2009. Somatic embryogenesis induction is promoted in knocked-out lines of Arabidopsis thaliana for a cDNA identified in non-embryogenic calluses of tamarillo (Cyphomandra betacea). New Biotechnol. 25, S300-S301. [ Links ]
Daigny, G., H. Paul, R. Sangwan, and B.S. Sangwan-Narreel. 1996. Factors influencing secondary somatic embryogenesis in Malus x domestica Borkh. (cv. 'Gloster 69'). Plant Cell Rep. 16, 153-157. [ Links ]
Dhavala, V.N.C., R.D. Tejeswara, V.R. Yechuri, and K. Prabavathi. 2009. Effect of explant age, hormones on somatic embryogenesis and production of multiple shoot from cotyledonary leaf explants of Solanum trilobatum L. Afr. J. Biotechnol. 8, 630-634. [ Links ]
George, E.F. and P.C. Debergh. 2008. Micropropagation: uses and methods. pp. 29-64. In: George, E.F., M.A. Hall, and G.-J. De Klerk (eds.). Plant propagation by tissue culture. Vol. 1. Springer, Dordercht, The Netherlands. [ Links ]
González, O.S., O. Sam, M.M. Hernández, M.J. Coronado, and J.J. Silva. 2005. Caracterización histológica de la embriogénesis somática a partir de limbos foliares de boniato (Hipomoea batatas L. Lam.). Cultivos Trop. 26, 37-41. [ Links ]
Hong, P.I., J.T. Chen, and W.C. Chang. 2008. Promotion of direct somatic embryogenesis of Oncidium by adjusting carbon sources. Biol. Plant. 52, 597-600. [ Links ]
Huda, A.K.M.N. and M.A. Bari. 2007. Production of synthetic seed by encapsulating asexual embryo in eggplant (Solanum melongena L.). Int. J. Agric. Res. 2, 832-837. [ Links ]
Iraqi, D., and F.M. Tremblay. 2001. The role of sucrose during maturation of black spruce (Picea mariana) and white spruce (Picea glauca) somatic embryos. Physiol. Plant. 111, 381-388. [ Links ]
JayaSree, T., U. Pavan, M. Ramesh, A.V. Rao, K. Jagan Mohan Reddy, and A. Sadanandam. 2001. Somatic embryogenesis from leaf cultures of potato. Plant Cell Tiss. Organ Cult. 64, 13-17. [ Links ]
Jheng, F.-Y., Y.-Y. Do, Y.-W. Liauh, J.-P. Chung, and P.-L. Huang. 2006. Enhancement of growth and regeneration efficiency from embryogenic callus cultures of Oncidium 'Gower Ramsey' by adjusting carbohydrate sources. Plant Sci. 170, 1133-1140. [ Links ]
Kantharajah, A.S. and P.G. Golegaonkar. 2004. Somatic embryogenesis in eggplant. Sci. Hortic. 99, 107-117. [ Links ]
Kaparakis, G. and P.G. Alderson. 2002. Influence of high concentrations of cytokinins on the production of somatic embryos by germinating seeds of tomato, aubergine and pepper. J. Hort. Sci. Biotechnol. 77, 186-190. [ Links ]
Kaparakis, G. and P.G. Alderson. 2008. Role for cytokinins in somatic embryogenesis of pepper (Capsicum annuum L.)? J. Plant Growth Regul. 27, 110-114. [ Links ]
Kintzios, S., J.B. Drossopoulos, E. Shortsianitis, and D. Peppes. 2000. Induction of somatic embryogenesis from young, fully expanded leaves of chilli pepper (Capsicum annuum L.): effect of leaf position, illumination and explant pretreatment with high cytokinin concentrations. Sci. Hortic. 85, 137-144. [ Links ]
Kintzios, S., J.B. Drossopoulos, and Ch. Lymperopoulos. 2001. Effect of vitamins and inorganic micronutrients on callus growth and somatic embryogenesis from leaves of chilli pepper. Plant Cell Tiss. Organ Cult. 67, 55-62. [ Links ]
Koleva-Gudeva, L.R., M. Spasenoski, and F. Trajkova. 2007. Somatic embryogenesis in pepper anther culture: The effect of incubation treatments and different media. Sci. Hortic. 111, 114-119. [ Links ]
Li, J.-R., F.Y. Zhuang, Ch.-G. Ou, H. Hu, Z.-W. Zhao, and J.-H. Mao. 2013. Microspore embryogenesis and production of haploid and doubled haploid plants in carrot (Daucus carota L.). Plant Cell Tiss. Organ Cult. 112, 275-287. [ Links ]
Mir, K.A., A.S. Dhatt, J.S. Sandhu, and S.S. Gosal. 2008. Genotype, explant and culture medium effects on somatic embryogenesis in eggplant (Solanum melongena L.). Hortic. Environ. Biotechnol. 49, 182-187. [ Links ]
Mishra, S., I. Sanyal, and D.V. Amla. 2012. Changes in protein pattern during different developmental stages of somatic embryos in chickpea. Biol. Plant. 56, 613-619. [ Links ]
Motallebi-Azar, A., S. Kazemiani, and F. Yarmohamadi. 2013. Effect of sugar/osmotica levels on in vitro microtuberization of potato (Solanum tuberosum L.). Russ. Agric. Sci. 39, 112-116. [ Links ]
Murashige, T. and F. Skoog. 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15, 473-497. [ Links ]
Na, H., J.-H. Kwak, and Ch. Chun. 2011. The effects of plant growth regulators, activated charcoal, and AgNO3 on microspore derived embryo formation in broccoli (Brassica oleracea L. var. italica). Hort. Environ. Biotechnol. 52, 524-529. [ Links ]
Nakagawa, H., T. Saijyo, N. Yamauchi, M. Shigyo, S. Kako, and A. Ito. 2001. Effects of sugars and abscisic acid on somatic embryogenesis from melon (Cucumis melo L.) expanded cotyledon. Sci. Hortic. 90, 85-92. [ Links ]
Nakano, M., K. Kigoshi, T. Shimizu, T. Endo, T. Shimada, H. Fujii, and M. Omura. 2013. Characterization of genes associated with polyembryony and in vitro somatic embryogenesis in Citrus. Tree Genet. Genomes 9, 795-803. [ Links ]
Oláh, R., A. Zok, A. Pedryc, S. Howard, and L.G. Kovács. 2009. Somatic embryogenesis in a broad spectrum of grape genotypes. Sci. Hortic. 120, 134-137. [ Links ]
Parimalan, R., A. Venugopalan, P. Giridhar, and G.A. Ravishankar. 2011. Somatic embryogenesis and Agrobacterium-mediated transformation in Bixa orellana L. Plant Cell Tiss. Organ Cult. 105, 317-328. [ Links ]
Qin, Y., H.-L. Li, and Y.-D. Guo. 2007. High-frequency embryogenesis, regeneration of broccoli (Brassica oleracea var. italica) and analysis of genetic stability by RAPD. Sci. Hortic. 111, 203-208. [ Links ]
Quainoo, A.K., A.C. Wetten, and J. Allainguillaume. 2008. The effectiveness of somatic embryogenesis in eliminating the cocoa swollen shoot virus from infected cocoa trees. J. Virol. Methods 149, 91-96. [ Links ]
Ramakrishnan, M., S.A. Ceasar, V. Duraipandiyan, M.A. Daniel, and S. Ignacimuthu. 2013. Efficacious somatic embryogenesis and fertile plant recovery from shoot apex explants of onion (Allium cepa L.). Vitro Cell. Dev. Biol. Plant 49, 285-293. [ Links ]
Ren, Y., H. Bang, I.S. Curtis, J. Gould, B.S. Patil, and K.M. Crosby. 2012. Agrobacterium-mediated transformation and shoot regeneration in elite breeding lines of western shipper cantaloupe and honeydew melons (Cucumis melo L.). Plant Cell Tiss. Organ Cult. 108, 147-158. [ Links ]
Ribeiro, A.P.O., E.A.T. Picoli, E.R.G. Lani, W.A. Vendrame, and W.C. Otoni. 2009. The influence of flask sealing on in vitro morphogenesis of eggplant (Solanum melongena L.). In Vitro Cell. Dev. Biol. Plant 45, 421-428. [ Links ]
Rout, G.R., A. Mohapatra, and S.M. Jain. 2006. Tissue culture of ornamental pot plant: a critical review on present scenario and future prospects. Biotechnol. Adv. 24, 531-560. [ Links ]
Sánchez-Enciso, M.C., L.J. Rodríguez de la O., G.P. Zárate-de Lara, A. López-Herrer, S. Barrales-Domínguez, and G. González-de la Cruz. 2005. Embriogénesis somática en papa cvs. Atlantic y Fritolay-1867. Rev. Chapingo Serie Hortic. 11, 219-224. [ Links ]
Santa-Maria, M., K.V. Pecota, C.G. Yencho, G. Allen, and B. Sosinski. 2009. Rapid shoot regeneration in industrial 'high starch' sweetpotato (Ipomoea batatas L.) genotypes. Plant Cell Tiss. Organ Cult. 97, 109-117. [ Links ]
Santos, D. and P. Fevereiro. 2002. Loss of DNA methylation affects somatic embryogenesis in Medicago truncatula. Plant Cell Tiss. Organ Cult. 70, 155-161. [ Links ]
Segovia B., V. 2002. Optimización de la regeneración de lulo (Solanum quitoense) orientada a la transformación genética de las plantas. M.Sc. thesis. Master in Plant Biotechnology, Universidad Internacional de Andalucía; International Center for Tropical Agriculture (CIAT ), Huelva, Spain. [ Links ]
Sharma, S.K., S. Millam, P.E. Hedley, J. McNicol, and G.J. Bryan. 2008. Molecular regulation of somatic embryogenesis in potato: an auxin led perspective. Plant Mol. Biol. 68, 185-201. [ Links ]
Singh, S. and S. Hazra. 2009. Somatic embryogenesis from the axillary meristems of peanut (Arachis hypogaea L.). Plant Biotechnol. Rep. 3, 333-340. [ Links ]
Solís-Ramos, L.Y., S. Nahuath-Dzib, A. Andrade-Torres, F. Barredo-Pool, T. González-Estrada, and E. Castaño de la Serna. 2010. Indirect somatic embryogenesis and morphohistological analysis in Capsicum chinense. Biologia 65, 504-511. [ Links ]
Tan, C.L. and D.B. Furtek. 2003. Development of an in vitro regeneration system for Theobroma cacao from mature tissues. Plant Sci. 164, 407-412. [ Links ]
Thornton, M.K., J. Lee, R. John, N.L. Olsen, and D.A. Navarre. 2013. Influence of growth regulators on plant growth, yield, and skin color of specialty potatoes. Am. J. Potato Res. 90, 271-283. [ Links ]
Van Der Vyver, C. 2010. Genetic transformation of the euploid Saccharum officinarum via direct and indirect embryogenesis. Sugar Tech. 12, 21-25. [ Links ]
Van Staden, J., E. Zazimalova, and E.F. George. 2008. Plant growth regulators II: cytokinins, their analogues and antagonists. pp. 205-226. In: George, E., P. Debergh, and G. De Klerk (eds.). Plant propagation by tissue culture. Springer, Dordercht, The Netherlands. [ Links ]
Vargas, T.E., E. de García, and M. Oropeza. 2005. Somatic embryogenesis in Solanum tuberosum from cell suspension cultures: histological analysis and extracellular protein patterns. J. Plant Physiol. 162, 449-456. [ Links ]
Vinoth, S., P. Gurusaravanan, and N. Jayabalan. 2014. Optimization of somatic embryogenesis protocol in Lycopersicon esculentum L. using plant growth regulators and seaweed extracts. J. Appl. Phycol. 26, 1527-1537. [ Links ]
Wang, Y.-X., X.F. Wang, Z.-Y. Ma, G.-Y. Zhang, and G.-Y. Hang. 2006. Somatic embryogenesis and plant regeneration from two recalcitrant genotypes of Gossypium hirsutum L. Agric. Sci. China 5, 323-329. [ Links ]
Yang, J.L., E.S. Seong, M.J. Kim, B.K. Ghimire, W.H. Kang, W.Y. Yu, and Ch.H. Li. 2010. Direct somatic embryogenesis from pericycle cells of broccoli (Brassica oleracea L. var. italica) root explants. Plant Cell Tiss. Organ Cult. 100, 49-58. [ Links ]
Zapata-Castillo, P.Y., A.-C. Flick, G. López-Puc, A. Solís-Ruiz, F. Barahona-Pérez, and N. Santana-Buzzy. 2007. Somatic embryogenesis in habanero pepper (C. chinense Jacq.) from cell suspensions. HortScience 42, 329-333. [ Links ]
Zhang, Q., K.M. Folta, and T.M. Davis. 2014. Somatic embryogenesis, tetraploidy, and variant leaf morphology in transgenic diploid strawberry (Fragaria vesca subspecies vesca 'Hawaii 4') (online). BMC Plant Biol. 14, http://www.biomedcentral.com/content/pdf/1471-2229-14-23.pdf; consulted: June, 2014. [ Links ]
