










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.33 no.1 Bogotá Jan./Apr. 2015
https://doi.org/10.15446/agron.colomb.v33n1.48412
Doi: 10.15446/agron.colomb.v33n1.48412
1 Tropical Forages, Centro Internacional de Agricultura Tropical (CIAT). Cali (Colombia). j.c.jimenez@cgiar.org
2 Department of Agronomy, Faculty of Agricultural Sciences, Universidad Nacional de Colombia. Bogota (Colombia)
Received for publication: 19 January, 2015. Accepted for publication: 30 March, 2015.
Abstract
As a consequence of global warming, rainfall is expected to increase in several regions around the world. This, together with poor soil drainage, will result in waterlogged soil conditions. Brachiaria grasses are widely sown in the tropics and, these grasses confront seasonal waterlogged conditions. Several studies have indicated that an increase in nutrient availability could reduce the negative impact of waterlogging. Therefore, an outdoor study was conducted to evaluate the responses of two Brachiaria sp. grasses with contrasting tolerances to waterlogging, B. ruziziensis (sensitive) and B. humidicola (tolerant), with two soil fertility levels. The genotypes were grown with two different soil fertilization levels (high and low) and under well-drained or waterlogged soil conditions for 15 days. The biomass production, chlorophyll content, photosynthetic efficiency, and macro- (N, P, K, Ca, Mg and S) and micronutrient (Fe, Mn, Cu, Zn and B) contents in the shoot tissue were determined. Significant differences in the nutrient content of the genotypes and treatments were found. An increase of redoximorphic elements (Fe and Mn) in the soil solution occurred with the waterlogging. The greater tolerance of B. humidicola to waterlogged conditions might be due to an efficient root system that is able to acquire nutrients (N, P, K) and potentially exclude phytotoxic elements (Fe and Mn) under waterlogged conditions. A high nutrient availability in the waterlogged soils did not result in an improved tolerance for B. ruziziensis. The greater growth impairment seen in the B. ruziziensis with high soil fertility and waterlogging (as opposed to low soil fertility and waterlogging) was possibly due to an increased concentration of redoximorphic elements under these conditions.
Key words: tropical pastures, adaptation, abiotic stress, nutrients.
Resumen
Como consecuencia del calentamiento global se prevén aumentos en las precipitaciones de algunas regiones alrededor del mundo. Esto, unido a malos drenajes de los suelos, provoca condiciones de encharcamiento. Las pasturas de Brachiaria son sembradas ampliamente en los trópicos americanos y periódicamente están expuestas a condiciones de anegamiento. Se ha demostrado que en condiciones de anegamiento, un aumento en la disponibilidad de nutrientes puede aumentar la producción de biomasa de las plantas. Por este motivo se condujo un estudio para evaluar las respuestas de dos genotipos de Brachiaria contrastantes por su tolerancia a inundación: B. ruziziensis (sensible) y B. humidicola (tolerante). Los genotipos crecieron en suelo con dos niveles contrastantes de fertilidad (alta y baja) y en condiciones de drenaje o anegamiento durante 15 días. Producción de biomasa, contenido de clorofila, eficiencia fotosintética y contenido de macro (N, P, K, Ca, Mg y S) y micro nutrientes (Fe, Mn, Cu, Zn y B) en parte aérea fueron determinados. Diferencias estadísticas en el contenido de nutrientes entre los diferentes genotipos y tratamientos fueron encontradas. Así mismo, se presentó un incremento en los elementos redoximórficos (Fe y Mn) en suelos anegados. La gran tolerancia de B. humidicola a suelos anegados pudo ser debida a un eficiente sistema radical capaz de tomar nutrientes (N, P, K) y excluir nutrientes potencialmente fitotóxicos (Fe y Mn) bajo condiciones de inundación. La mayor disponibilidad de nutrientes en suelo no aumentó la producción de biomasa en B. ruziziensis bajo condiciones de anegamiento. Se observó una gran disminución de crecimiento de B. ruziziensis en suelos anegados y alta fertilidad (en comparación a baja fertilidad y anegamiento), posiblemente asociada a un aumento en la concentración de elementos redoximórficos (Fe y Mn) en suelos anegados.
Palabras clave: pasturas tropicales, adaptación, estrés abiótico, nutrientes.
Introduction
Due to global climate change, it is anticipated that rainfall will increase in magnitude and frequency (IPCC, 2007; Jiang et al., 2008). The combination of high rainfall and poor drainage conditions leads to waterlogged soils (Caetano and Dias-Filho, 2008). Waterlogging in soils is a major abiotic stress that affects the growth and productivity of several plant species around the world (Jackson and Colmer, 2005). Gas diffusion in waterlogged soils is about 10,000 times lower than in the air (Colmer, 2003). The lower gas diffusion, combined with root and microorganism respiration, results in a drastic reduction of oxygen in the soil (Elzenga and Van Veen, 2010; George et al., 2012). When there is limited soil oxygen, aerobic root respiration, energy production (Kulichikhin et al., 2008) and nutrient uptake are decreased in plants (Armstrong and Drew, 2002).
Waterlogged conditions usually result in a reduced soil status, which implies a change in the availability of some redoximorphic nutrients (i.e. Fe2+ and Mn2+). This change in redoximorphic nutrient solubility could reach phytotoxic concentrations in plants (Setter and Waters, 2003) or affect the uptake of other nutrients (Fageria, 2001).
Brachiaria grasses are widely planted in the tropics to sustain livestock production (Miles et al., 2004). In several tropical zones, pastures face seasonal conditions of waterlogged soils (Baruch, 1994). Waterlogging could result in significant reductions of Brachiaria grass production and, therefore, milk and meat production (Rao et al., 2011).
In the Brachiaria genus, highly phenotypic variations exist, in terms of production and adaptation to several stress conditions (Peters et al., 2011). For example, B. ruziziensis shows poor adaptation to waterlogged conditions but it has a high forage quality. In contrast, B. humidicola has low productivity and nutritional value for livestock, but it is waterlogging tolerant. The outstanding adaptation to waterlogged soil conditions seen in B. humidicola is mainly based on a combination of root morpho-anatomical attributes (Cardoso et al., 2013, 2014).
There is a lack of agronomic practices to counteract the deleterious effect of waterlogged soils in Brachiaria forage grasses. Some studies have indicated that tolerance to waterlogging could be improved with an increase of soil fertility (Day, 1987; Huang et al., 1995; Xie et al., 2009). Therefore, the main objective of the present study was to evaluate the responses of B. humidicola (waterlogging tolerant) and B. ruziziensis (waterlogging sensitive) under waterlogged soil conditions and grown in two different soil fertility levels (high and low). The knowledge developed here should improve our understanding of the physiological mechanisms related to adaptation to waterlogged soils and also may serve for the development of strategies aimed at reducing the negative impact of waterlogging on Brachiaria grasses and on other tropical forage grasses.
Materials and methods
Plant material and growth conditions
Vegetative propagules from B. humidicola (CIAT 679) and B. ruziziensis (Br 4402) with good vigor under uniform growing conditions were selected. Two vegetative propagules per genotype were planted in plastic pots (15.8 cm height, 20.0 cm outer diameter and 15.5 cm inner diameter) filled with 3.5 kg of an Oxisol soil collected in the Altillanura region of the Llanos Orientales in Colombia. This soil was classified as Typic Haplustox Isohipertermic kaolinitic (Soil Survey Staff, 1994). The soil was mixed with sand (2:1 w/w). The soil mixture was fertilized with two levels of nutrient application (high and low), in accordance with Rao et al. (1992).
The plants were grown in the patio of Tropical Forages Program of the International Center for Tropical Agriculture CIAT (Cali, Colombia) for 50 d. During the establishment period, the plants were irrigated daily to maintain the soil at field capacity. The nutrient application rate for the high soil fertility was (kg ha-1) 40.0 N, 50.0 P, 100.0 K, 101.0 Ca, 28.4 Mg, 20.0 S, 2.0 Zn, 2.0 Cu, 0.1 B and 0.1 Mo and, for the low soil fertility, it was: (kg ha-1) 0 N, 20.0 P, 20.0 K, 47.0 Ca, 14.2 Mg, 10.0 S and no micronutrients.
Once the plants were established, the waterlogging and well-drained treatments were imposed for 15 d. In the waterlogging treatment, pots were inserted into similarly sized pots and lined with a plastic bag to prevent water leakage. During the waterlogging treatment, a water lamina of 2 cm above the soil surface was maintained throughout the experimental period. In the well-drained treatment, pots were inserted into similarly sized pots without a plastic bag and were irrigated regularly in order to maintain soil moisture of 80 - 100% of the field capacity.
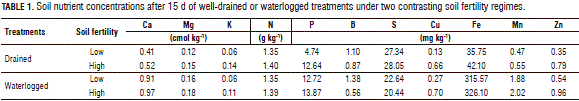
A complete block design with a factorial structure was used, three replications and two treatments with two sub-factors (water level "waterlogging and irrigation" and soil fertility "high and low"). The soil characteristics of the different treatments are in Tab. 1. During the experiment, the average temperature was 24.4°C, with a relative humidity of 59.7%, radiation at 15.9 MJ m-2 d-1 and effective sunlight at 4.1 h d-1.
Plant measurements
The leaf chlorophyll content in the SPAD units (soil plant analysis development) was estimated with a SPAD chlorophyll meter (Konica, Minolta, Tokyo, Japan). The total chlorophyll content in the leaves was first determined for B. ruziziensis and B. humidicola with cold-methanol extracted chlorophyll, using the method described by Lichtenthaler and Wellburn (1983) and plotted against the corresponding SPAD values. The SPAD values were converted to total chlorophyll content (mg g-1) using the following formula: Chl (mg g-1) = 0.4036 (SPAD meter reading value) - 0.5527 (R2 = 0.93). The leaf photosynthetic efficiency (fv'/fm') was determined using an Fp100 fluorpen (Photon Systems Instruments, Drásov, Czech Republic). Measurements of the SPAD and photosynthetic efficiency were performed in three fully expanded leaves after 15 d of the waterlogging treatments. Fifteen days after the imposition of the treatments, green leaves were separated from the stems for determination of the green leaf area and dry weight. The green leaf area per plant was quantified using a Licor-3100 (LI-COR, Lincoln, NE). The shoot dry mass was measured after drying the samples for 3 d at 60°C. After that, a waterlogging tolerance coefficient (WTC) was obtained following Liu et al. (2010) and expressed as a percentage:

Nutrient concentration in shoot tissue
The shoot nutrient concentration was determined by the Analytical Services Laboratory of CIAT. Both dry stems and dry green leaves were mixed, ground and passed through a 0.1 mm sieve. After that, the ground samples were dissolved in an acid solution (percloric-sulfuric acid). The contents of Ca, Mg, K, Na, Cu, Fe, Mn and Zn were determined by atomic absorption spectrophotometry and the P, B and N contents by spectrometry (Mackean, 1993).
Statistical analysis
Analyses of variance (ANOVA) were performed to determine the statistical differences between the treatments and variables. Additionally, post-ANOVA were performed using the Duncan test to observe differences between the treatments and genotypes. The statistical analysis was carried out using the Agricolae library (Mendiburu, 2014) of the statistical software R (R core team, 2012).
Results and discussion
Under waterlogged conditions and independent of the soil fertility level, the shoot biomass, green leaf area, chlorophyll content and photosynthetic efficiency of B. ruziziensis were significantly reduced (Tab. 2). Contrary to what was expected, the reduction of these parameters was more affected by high soil fertility and waterlogged conditions (Tab. 2). Independent of the soil fertility, the shoot biomass, green leaf area, chlorophyll content and photosynthetic efficiency of B. humidicola did not present significant differences with waterlogged conditions when compared to the plants grown under drained conditions (Tab. 2).
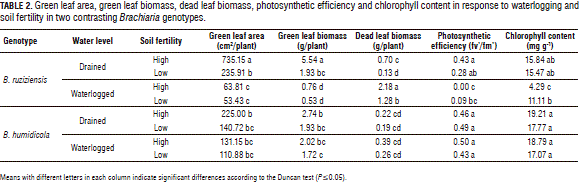
Under waterlogged conditions and low soil fertility, B. ruziziensis showed a higher chlorophyll content and a higher photosynthetic efficiency, as compared to the plants grown with waterlogged conditions and high soil fertility (Tab. 2). It is known that aerenchyma formation in roots can be induced by nutrient deficiency in soils (Fan et al., 2003; Postma and Lynch, 2011). Aerenchyma refers to a vegetal tissue with large and interconnected spaces filled with gas (Evans, 2004). Under waterlogged conditions, aerenchyma formation maintains the aerobic respiration of roots (Visser et al., 2000; Colmer, 2003), allowing the plant to adapt to waterlogged soil conditions. Recent studies have indicated that several genotypes of Brachiaria develop root aerenchyma as a mechanism of adaptation to waterlogged soil conditions (Cardoso et al., 2013, 2014). According to this, it was possible that the establishment of B. ruziziensis with low soil fertility promoted root aerenchyma formation before the imposition of waterlogging stress, which in turn could have increased the fitness of B. ruziziensis under the combined stresses of low fertility and waterlogging.
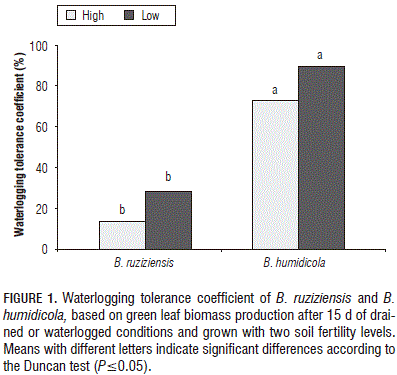
In the present study, B. ruziziensis presented a greater shoot vigor than B. humidicola, under drained or waterlogged conditions (Tab. 2). However, the B. ruziziensis WTC was significantly reduced under waterlogged conditions, particularly with the high soil fertility (Fig. 1). WTC is an index that is used to detect waterlogging tolerance based on specific characteristics (Liu et al., 2010). Using the WTC, we were able to detect that vigor and waterlogging tolerance are not related in these two genotypes.
The shoot N concentration of B. ruziziensis with waterlogged and high soil fertility conditions was significantly higher than under drained conditions (Fig. 1). However, the chlorophyll content was lower under the same conditions (Tab. 2). It is therefore likely that these greater shoot N concentrations could be associated with a high remobilization of this nutrient from senescing leaves (Tab. 2).
The concentrations of K, Mg, S, Cu and Mn in the B. ruziziensis tissues with waterlogged conditions were significantly reduced in comparison to the drainage treatment (Figs. 2, 3 and 4). Armstrong and Drew (2002) observed that hypoxia in the root zone can inhibit the uptake and transport of nutrients. Likewise, a low nutrient concentration in tissues can be associated with a lack of energy for nutrient uptake (George et al., 2012). Under waterlogged and high soil fertility conditions, the B concentration in the B. ruziziensis leaves was significantly increased (Fig. 3), which could be one of the possible toxic effects of waterlogged soils (Setter et al., 2009).
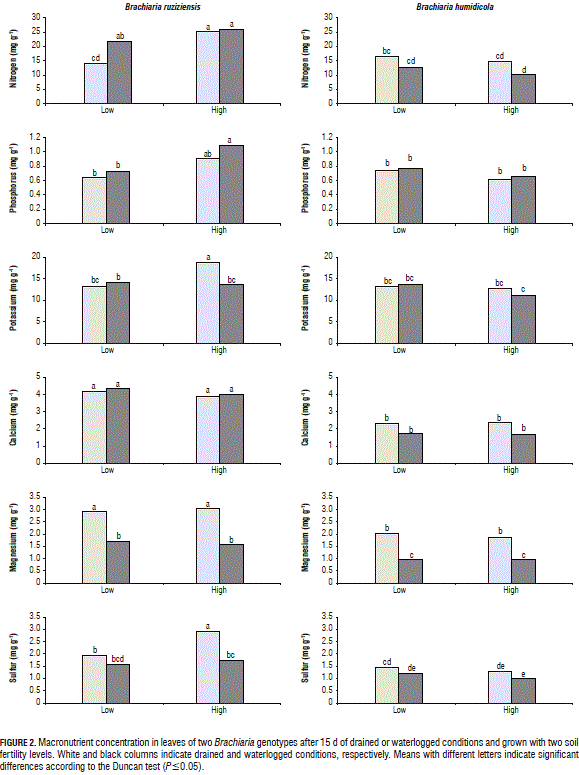
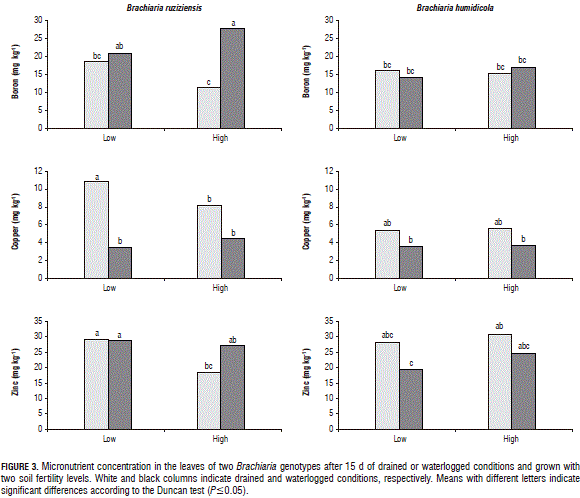
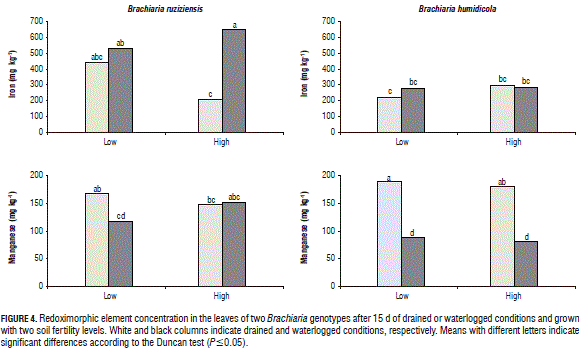
Under waterlogged conditions and independent of the soil fertility level, the concentration of N, P, K, Ca, S, B, Cu, Zn and Fe in the shoot biomass of B. humidicola was not affected (Figs. 2, 3 and 4). Under waterlogged conditions, the Mg concentration in the shoot biomass of both genotypes was significantly reduced with respect to the drained conditions (Fig. 2). A similar situation was found in wheat (Stieger and Feller, 1994). This decrease in the Mg concentration in the shoot biomass could be related to excess Mn (Turner and Barkus, 1983) or Fe in waterlogged soils, which leads to a nutrient antagonism in root uptake.
With waterlogging and high soil fertility, B. ruziziensis showed an excess Fe concentration in the shoots, as compared to the drained conditions (Fig. 4). This excess was classified by Setter et al. (2009) as one of the major problems faced by plants in waterlogged conditions. This could indicate that the significant reduction in the shoot biomass, chlorophyll content and photosynthetic efficiency exhibited by B. ruziziensis with waterlogged conditions and high soil fertility (Tab. 2) was associated with the elemental toxicity of redoximorphic elements. Similar cases have been reported in wheat plants growing in waterlogged soils, in which the concentration of Mn and Fe in the tissues increased 10 or more times, in comparison with plants grown with drained conditions (Khabaz-Saberi et al., 2006; Setter et al., 2009).
Setter et al. (2009) noted that waterlogging tolerance is not exclusively associated with differences in the root aeration capacity (i.e. aerenchyma development) but also with traits that are in turn associated with tolerance to microelement toxicity. Likewise, Close and Davidson (2003) found that the waterlogging acclimation of Eucalyptus is associated with tolerance to nutritional imbalances in soils. Several authors have indicated that, though waterlogged conditions reduce plant growth, an exogenous application of nutrients generates a positive effect on plant performance under these conditions (George et al., 2012), as shown in Acer rubrum (Day, 1987), wheat (Huang et al., 1995), and Deyeuxia angustifolia (Xie et al., 2009).
Under waterlogged conditions, B. humidicola did not show significant differences in the microelement concentration in tissues, with respect to drained conditions (Fig. 3). Under waterlogged conditions, this grass presents suberized cells in the outer part of the root (Cardoso et al., 2013). Suberized cells in the outer part of the root have been linked to the development of a barrier that impedes radial oxygen loss (De Simone et al., 2003; Soukup et al., 2007; Kotula et al., 2009). This barrier, besides impeding oxygen leakage from the roots to the rhizosphere, prevents the entrance of phytotoxic substances commonly found in waterlogged soils (Kotula et al., 2014; Manzur et al., 2015). Therefore, it is likely that this adaptive trait improves the root properties for the internal aeration system of B. humidicola, prevents the entry of excess redoximorphic elements found in waterlogged soils and, therefore, contributes to a better adaptation to this stressful condition, as shown by the unaffected levels of chlorophyll content, photosynthetic efficiency and growth.
Conclusions
The results arising from this study suggest that increasing soil fertility does not improve the waterlogging tolerance of the sensitive genotype B. ruziziensis. The likely development of aerenchyma in B. ruziziensis under low soil nutrient concentrations before the imposition of waterlogging probably maintained the nutrient uptake starting at the onset of the stress. This in turn was reflected in similar values of chlorophyll content, photosynthetic efficiency and biomass production of the plants grown with drained conditions and low soil fertility. With waterlogging, B. humidicola showed no differences in the shoot Fe concentration, suggesting that this species has a mechanism that limits the entry of toxic elements found in excess in waterlogged soils. This could contribute to the outstanding plant performance that B. humidicola exhibits under waterlogged conditions.
Acknowledgements
This work was partially funded by Fontagro (USA) (project number: FTG-8060/08).
Literature cited
Armstrong, W. and M.C. Drew. 2002. Root growth and metabolism under oxygen deficiency. pp. 729-761. In: Waisel, Y., A. Eshel, and U. Kafkafi (eds.). Plant roots: the hidden half. 3rd ed. Marcel Dekker, New York, NY. [ Links ]
Baruch, Z. 1994. Responses to drought and flooding in tropical forage grasses. Plant Soil 164, 87-96, 97-105. Doi: 10.1007/BF00010114; 10.1007/BF00010115 [ Links ]
Caetano, L.P.S. and M.B. Dias-Filho. 2008. Responses of six Brachiaria spp. accessions to root zone flooding. R. Bras. Zootec. 37, 795-801. Doi: 10.1590/S1516-35982008000500003 [ Links ]
Cardoso, J.A., J. Rincón, J.C. Jiménez, D. Noguera, and I.M. Rao. 2013. Morpho-anatomical adaptations to waterlogging by germoplasm accessions in a tropical forage grass. AoB Plants 5, plt047. Doi: 10.1093/aobpla/plt047 [ Links ]
Cardoso, J.A., J.C. Jiménez, and I.M. Rao. 2014. Waterlogging-induced changes in root architecture of germplasm accessions of the tropical forage grass Brachiaria humidicola. AoB Plants 6, plu017 Doi: 10.1093/aobpla/plu017 [ Links ]
Close, D.C. and N.J. Davidson. 2003. Long-term waterlogging: nutrient, gas exchange, photochemical and pigment characteristics of Eucalyptus nitens saplings. Russ. J. Plant Physiol. 50, 843-847. Doi: 10.1023/B:RUPP.0000003284.25827.95 [ Links ]
Colmer, T.D. 2003. Long-distance transport of gases in plants: a perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ. 26, 17-36. Doi: 10.1046/j.1365-3040.2003.00846.x [ Links ]
Day Jr, F.P. 1987. Effects of flooding and nutrient enrichment on biomass allocation in Acer rubrum seedlings. Amer. J. Bot. 74, 1541-1554. Doi: 10.2307/2444048 [ Links ]
De Simone, O., K. Haase, E Müller, W.J. Junk, K. Hartmann, L. Schreiber, and W. Schmidt. 2003. Apoplasmic barriers and oxygen transport properties of hypodermal cell walls in roots from four Amazonian tree species. Plant Physiol. 132, 206-217. Doi: 10.1104/pp.102.014902 [ Links ]
Elzenga, J.T.M. and H. Van Veen. 2010. Waterlogging and plant nutrient uptake. pp. 23-35. In: Mancuso, S. and S. Shabala (eds.). Waterlogging signaling and tolerance in plants. Springer Verlag, Heidelberg, Germany. Doi: 10.1007/978-3-642-10305-6_2 [ Links ]
Evans, D.E. 2004. Aerenchyma formation. New Phytol. 161, 35-49. Doi: 10.1046/j.1469-8137.2003.00907.x [ Links ]
Fageria, V.D. 2001. Nutrient interactions in crop plants. J. Plant Nutr. 24, 1269-1290. Doi: 10.1081/PLN-100106981 [ Links ]
Fan, M., J. Zhu, C. Richards, K.M. Brown, and J.P. Lynch. 2003. Physiological roles for aerenchyma in phosphorus-stressed roots. Funct. Plant Biol. 30, 493-506. Doi: 10.1071/FP03046 [ Links ]
George, E., W.J. Horst, and E. Neumann. 2012. Adaptation of plant to adverse chemical soil conditions. pp. 409-472. In: Marschner, P. (ed.). Mineral nutrition of higher plants. 3rd ed. Academic Press, London. Doi: 10.1016/B978-0-12-384905-2.00017-0 [ Links ]
Huang, B., J.W. Johnson, D.S. NeSmith, and D.C. Bridges. 1995. Nutrient accumulation and distribution of wheat genotypes in response to waterlogging and nutrient supply. Plant Soil 173, 47-54. Doi: 10.1007/BF00155517 [ Links ]
IPCC, 2007. Climate change 2007: Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, UK. [ Links ]
Jackson, M.B. and T.D. Colmer. 2005. Response and adaptation by plants to flooding stress. Ann. Bot. 96, 501-505. Doi: 10.1093/aob/mci205 [ Links ]
Jiang, D., X. Fan, T. Dai, and W. Cao. 2008. Nitrogen fertiliser rate and post-anthesis waterlogging effects on carbohydrate and nitrogen dynamics in wheat. Plant Soil 304, 301-314. Doi: 10.1007/s11104-008-9556-x [ Links ]
Khabaz-Saberi, H., T.L. Setter, and I. Waters. 2006. Waterlogging induces high to toxic concentrations of iron, aluminum, and manganese in wheat varieties on acidic soil. J. Plant Nutr. 29, 899-911. Doi: 10.1080/01904160600649161 [ Links ]
Kotula, L., T.D. Colmer, and M. Nakazono. 2014. Effects of organic acids on the formation of the barrier to radial oxygen loss in roots of Hordeum marinum. Funct. Plant Biol. 41, 187-202. Doi: 10.1071/FP13178 [ Links ]
Kotula, L., K. Ranathunge, and E. Steudle. 2009. Apoplastic barriers effectively block oxygen permeability across outer cell layers of rice roots under deoxygenated conditions: roles of apoplastic pores and of respiration. New Phytol. 184, 909-917. Doi: 10.1111/j.1469-8137.2009.03021.x [ Links ]
Kulichikhin, K.Y., T.V. Chirkova, and K.V. Fagerstedt. 2008. Intracellular pH in rice and wheat root tips under hypoxic and anoxic conditions. Plant Signal Behav. 3, 240-242. Doi: 10.4161/psb.3.4.5151 [ Links ]
Lichtenthaler, H.K. and A.R. Wellburn. 1983. Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 11, 591-592. [ Links ]
Liu, Y.-Z. T Bin, Y.L. Zheng, K.-J. Ma, S.-Z. Xu, and F.-Z. Qiu. 2010. Screening methods for waterlogging tolerance at maize (Zea mays L.) seedling stage. Agr. Sci. China 9, 362-369. Doi: 10.1016/S1671-2927(09)60105-X [ Links ]
Mackean, S. 1993. Manual de análisis de suelos y plantas. International Center for Tropical Agriculture (CIAT), Cali, Colombia. [ Links ]
Manzur, M.E., A.A. Grimoldi, P. Insausti, and G.G. Striker. 2015. Radial oxygen loss and physical barriers in relation to root tissue age in species with different types of aerenchyma. Funct. Plant Biol. 42, 9-17. 10.1071/FP14078 [ Links ]
Mendiburu, F. 2014. Agricolae: statistical procedures for agricultural research. R package version 1.1-8. In: http://CRAN.R-project.org/package=agricolae; consulted: March, 2015. [ Links ]
Miles, J.W., C.B. Do Valle, I.M. Rao, and V.P.B. Euclides. 2004. Brachiaria grasses. pp. 745-783. In: Moser, L.E., B.L. Burson, and E.L. Sollenberger (eds.). Warm-season (C4) grasses. The American Society of Agronomy, Crop Science Society of America, Soil Science Society of America, Madison, WI. [ Links ]
Peters, M., L. Franco, A. Schmidt, and B. Hincapié. 2011. Especies forrajeras multipropósito: Opciones para productores del trópico americano. International Center for Tropical Agriculture (CIAT), Cali, Colombia. pp. 8-20. [ Links ]
Postma, J.A. and J.P. Lynch. 2011. Root cortical aerenchyma enhances the growth of maize on soils with suboptimal availability of nitrogen, phosphorus, and potassium. Plant Physiol. 156, 1190-1201.Doi: 10.1104/pp.111.175489 [ Links ]
R Core Team. 2012. A language and environmental for statistical computing. In: R Foundation for Statistical Computing, Vienna, Austria, www.R-project.org/; consulted: March, 2015. [ Links ]
Rao, I.M., W.M. Roca, M.A. Ayarza, E. Tabares, and R. Garcia. 1992. Somaclonal variation in plant adaptation to acid soil in the tropical forage legume Stylosanthes guianensis. Plant Soil 146, 21-30. Doi: 10.1007/BF00011991 [ Links ]
Rao, I., J. Miles, P. Wenzl, A. Louw-Gaume, J.A. Cardoso, J. Ricaurte, J. Polania, J. Rincon, V. Hoyos, E. Frossard, T. Wagatsuma, and W. Horst. 2011. Mechanisms of adaptation of brachiariagrasses to abiotic stress factors in the tropics. pp. 361-383. In: Proc. III International Symposium on Forage Breeding. Bonito, Brazil. [ Links ]
Setter, T.L. and I. Waters. 2003. Review of prospects for germplasm improvement for waterlogging tolerance in wheat, barley and oats. Plant Soil 253, 1-34. Doi: 10.1023/A:1024573305997 [ Links ]
Setter, T.L., I. Water, S.K. Sharma, K.N. Singh, N. Kulshreshtha, N.P.S. Yaduvanshi, P.C. Ram, B.N. Singh, J. Rane, G. McDonald, H. Khabaz-Saberi, T.B. Biddulph, R. Wilson, I. Barclay, R. McLean, and M. Cakir. 2009. Review of wheat improvement for waterlogging tolerance in Australia and India: the importance of anaerobiosis and element toxicities associated with different soils. Ann. Bot. 103, 221-235. Doi: 10.1093/aob/mcn137 [ Links ]
Soil Survey Staff. 1994. Keys to soil taxonomy. Pocahontas Press, Blacksburg, VA. [ Links ]
Soukup, A., W. Armstrong, L. Schreiber, R. Franke, and O. Votrubová. 2007. Apoplastic barriers to radial oxygen loss and solute penetration: a chemical and functional comparison of the exodermis of two wetland species, Phragmites australis and Glyceria maxima. New Phytol. 173, 264-278. Doi: 10.1111/j.1469-8137.2006.01907.x [ Links ]
Stieger, P.A. and U. Feller. 1994. Nutrient accumulation and translocation in maturing wheat plants grown on waterlogged soil. Plant Soil 160, 87-95. Doi: 10.1007/BF00150349 [ Links ]
Turner, D.W. and B. Barkus. 1983. Long-term nutrient absorption rates and competition between ions in banana in relation to supply of K, Mg and Mn. Fert. Res. 4, 127-134. Doi: 10.1007/BF01053249 [ Links ]
Visser, E.J.W., T.D. Colmer, C.W.P.M. Blom, and L.A.C.J. Voesenek. 2000. Changes in growth, porosity, and radial oxygen loss from adventitious roots of selected mono- and dicotyledonous wetland species with contrasting types of aerenchyma. Plant Cell Environ. 23, 1237-1245. Doi: 10.1046/j.1365-3040.2000.00628.x [ Links ]
Xie, Y., B. Ren, and F. Li. 2009. Increased nutrient supply facilitates acclimation to high-water level in the marsh plant Deyeuxia angustifolia: the response of root morphology. Aquatic Bot. 91, 1-5. Doi: 10.1016/j.aquabot.2008.12.004 [ Links ]

