










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkAgronomía Colombiana
versão impressa ISSN 0120-9965
Agron. colomb. vol.33 no.1 Bogotá jan./abr. 2015
https://doi.org/10.15446/agron.colomb.v33n1.46747
Doi: 10.15446/agron.colomb.v33n1.46747
1 Weed Management Supervisor, Monsanto Latin America North region. Mexico DF (Mexico). javierramirezsuarez@gmail.com
2 Department of Agronomy, Faculty of Agricultural Sciences, Universidad Nacional de Colombia. Bogota (Colombia).
Received for publication: 23 October, 2014. Accepted for publication: 30 March, 2015.
ABSTRACT
Phytosociological studies allow for the characterization and descriptive analysis of weed communities in crops. This study aimed to characterize the weed communities associated with weed crops in the "Centro, Meseta, and Norte" zones of the Tolima Department. The study was conducted in 96 commercial lots, in which a 1 ha area was marked off for the sampling. The development stage, density and cover of the weeds were evaluated. The importance value index, the alpha diversity indices of Shannon-Wiener, Simpson and uniformity as well as the similarity indices of Jaccard, Sorensen and Steinhaus were calculated. For the entire department, 42 weed species were identified, with Echinochloa colona being the principal one in all of the zones. In the Centro zone, 27 species were identified; in the Meseta zone, 31 species were identified; and, in the Norte zone, 38 species were identified. The alpha indices demonstrated that the Meseta zone was the most diverse. The Jaccard and Sorensen indices showed dissimilarity in the weed community for all of the comparisons of the zones. The Steinhaus coefficient registered the highest similarity intensity between the Centro and Norte zones.
Keywords: weed communities, importance value index, diversity index, cereals.
RESUMEN
Los estudios fitosociológicos permiten caracterizar y hacer análisis descriptivos de las comunidades de malezas de cultivos. Este trabajo tuvo como objetivo caracterizar las poblaciones de malezas asociadas a cultivos de arroz de las zonas Centro, Meseta y Norte del departamento del Tolima. Los levantamientos fueron realizados en 96 lotes comerciales, en cada lote se demarcó un área de evaluación de 1 ha en la cual se realizaron los muestreos. Se evaluó el estado de desarrollo, densidad y cobertura de las malezas. Se calculó el índice de valor de importancia, los índices de diversidad alfa de Shannon-Wiener, Simpson y de uniformidad así como los índices de similitud de Jaccard, Sorensen y Steinhaus. En todo el departamento se identificaron 42 especies de malezas siendo Echinochloa colona, la principal en todas las zonas. En la zona Centro fueron identificadas 27 especies; en la zona Meseta 31 y en la zona Norte 38. Los índices alfa registraron que la zona Meseta fue la más diversa. Los índices Jaccard y Sorensen mostraron disimilitud en la comunidad de malezas en todas las comparaciones de las zonas. El coeficiente Steinhaus registró mayor intensidad de similitud entre las zonas Centro y Norte.
Palabras clave: poblaciones de malezas, índice de valor de importancia, índice de diversidad, cereales.
Introduction
Agricultural activities generate changes and filters for biological communities and weeds that are associated with crops are exposed to disruptive factors that make their populations dynamic over time (Booth et al., 2003). In terms of responses to these agents of change, not all species in an agricultural system are equally important, with differences in frequency, density, and growth habit making some species the principal ones that generate economic and secondary effects that normally do not present problems for yield (Pitelli, 2000).
One of the more utilized methods for the analysis of weed communities in agricultural systems is the phytosociological study. Phytosociology is defined as the science that studies plant communities from the floristic, ecological, and dynamic points of view or as the science that studies plant groupings, their interactions, and their dependence on their environment (Ferriol and Merle, 2006).
A quantitative phytosociological study of a weed community in a defined area and time provides a momentary analysis of the plant composition, providing a tool that supplies various inferences for a plant community (Erasmo et al., 2004). The analysis of weed communities can be approached with the description of their characteristics, employing tools such as similarity and diversity indices that clarify their performance.
Alpha diversity (a) in studies on weed populations measures the amount of diversity within a defined community in a zone (Booth et al., 2003). For weed ecology, the Margalef, Shannon-Wiener, and Simpson indices are more commonly used. The Margalef index focuses on the richness of the species found in a studied population. The Shannon-Wiener index is based on the proportional abundance of each species and the Simpson index is based on the probability that two individuals in a community sample will be of the same species (Booth et al., 2003).
The study of the beta diversity (b) in weeds measures the change in weed species diversity between zones and samples the similarity of the composition of the community between location pairs through the calculation of similarity indices (Booth et al., 2003). The Jaccard and Sorensen index and the Steinhaus coefficient are more common in studies on weeds in agricultural systems. The Jaccard and Sorensen indices only consider how many species are in-common in a pair of evaluated communities and do not take into account the abundance of each species. For its part, the Steinhaus similarity coefficient or index incorporates abundance data into its analysis, taking into account the differences that occur in this data, and so is considered more valuable than the other indices (Booth et al., 2003).
For its part, the importance value index (IVI) determines the dominance of the species and the degree of heterogeneity of the agroecosystem (Pitelli, 2000), allowing for the evaluation of the horizontal structure of the community through the relative dominances, abundances, and frequencies (Lamprecht, 1990). The relative density is the number of individuals of a species or absolute density of a species over the total number of individuals or total density of all of the species (Brighenti et al., 2003). Relative frequency is defined as the frequency of a species over the sum of the frequencies of all of the species or the total frequency of all of the species (Mueller-Dombois and Ellenberg, 1976). Relative dominance (relative cover) is defined as the absolute dominance of a species over the dominance of all of the species (Mueller-Dombois and Ellenberg, 1976) or as the cover of a species over the total cover of all of the species, expressed as a percentage (Cantillo et al., 2006).
Based on the these considerations and considering the evident importance that this type of study has for the characterization and study of plant communities associated with commercial crops, the present study was developed with the aim of identifying the floristic composition and of characterizing the weed populations of commercial rice crops in the Centro, Meseta, and Norte zones of the department of Tolima in four evaluations carried out between emergence and formation of the flower primordia in the crops.
Materials and methods
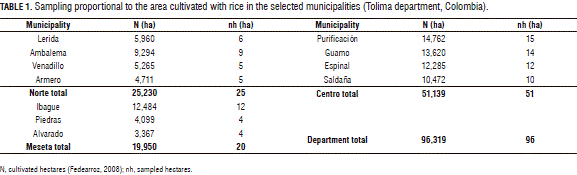
The present study was carried out between July of 2012 and February of 2013 in commercial crops in the department of Tolima, which were divided into three production zones in accordance with the different climatic, topographic, edaphical, and irrigation conditions. In each zone, the municipalities with the larger cultivated areas were used, with 96,319 ha of cultivated rice per year (Fedearroz, 2008), distributed as follows: the Norte zone (26% of the area), the Meseta zone of Ibague (21% of the area) and the Centro zone (53% of the area) with the municipalities of Purificación, Guamo, Espinal and Saldaña (Colombia) (Tab. 1).
The sample size was 0.1% (96 ha) of the total area in accordance with the following equation (Spiegel, 1988):
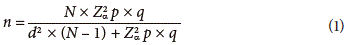
Where,
n = sample size.
N = total of the universal sample (in this case the 96,319 ha cultivated per year). 1.962 with a confidence of 95%.
p = expected proportion (5% = 0.05).
q = 1-p (in this case: 1-0.05 = 0.95).
d2 = precision (in this case 10%).
The number of hectares sampled per zone and municipality was distributed in proportion to the stratum, using the cultivated area as the criterion (Tab. 1). The methodology used to distribute the sampling units in the study area resulted in the sampling of an actual area of 384 ha for the entire study (0.4% of the total cultivated area in the selected municipalities) and 0.8 ha in each lot.
For the phytosociological study and characterization of the weed communities, a 0.04 m2 sampling square was used, which was thrown randomly five times (5) within the marked off hectare in each lot, following a zigzag pattern. Each hectare represented a commercial lot in accordance with reports from Erasmo et al. (2004) and Plaza and Hernández (2014). Four samples were conducted during the development of the crop: before the application of the first post-emergence control method (7 to 22 days after sowing, das), after the first control method (22-35 das), after the second post-emergence control method (37-52 das) and once the herbicide applications were finished, during the flower primordia formation stage of the crop (52-65 das). In each sampling, the variables of density and cover were measured for each of the encountered weed species. The species were identified using the studies conducted by Fuentes et al. (2006a), Fuentes et al. (2006b) and Montealegre (2011) as references.
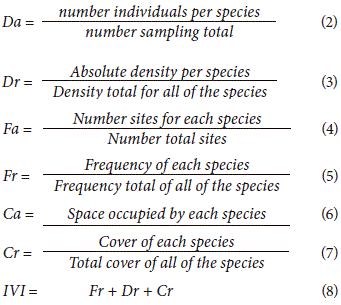
The importance value index (IVI) was calculated under the following parameters for each species: absolute density (Da), relative density (Dr), absolute frequency (Fa), relative frequency (Fr), cover (Ca), and relative cover (Cr), in accordance with Curtis and Mclntosh (1950) and Mueller-Dombois and Ellenberg (1976):
Among the alpha diversity indices (a), the Shannon-Wiener index (H), the Simpson dominance index (D) and the uniformity index (E) were calculated in accordance with the equations cited by Booth et al. (2003).
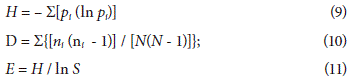
Where,
pi= proportional abundance of each species
ni = number of individuals per species
N = number of total individuals
S = number of total of species richness
H = Shannon-Wiener index
On the other hand, of the beta diversity indices (b), the similarity indices of Jaccard (SJ) and Sorensen (SS) and the Steinhaus coefficient (SST) were calculated in accordance with the methodology cited by Booth et al. (2003).

Where,
j = number of species found in both communities
a = number of species found only in community a
b = number of species found only in community b
W = Sum of the lower of the two abundances of each species
A and B = total of the abundances of each community
Results and discussion
Floristic composition
In the rice production areas of Tolima, 42 weed species were identified, grouped into 2 classes, 20 families, and 31 genera. In the zones, the Centro zone presented 27 weed species from 14 families and 21 genera; the Meseta zone had 31 species from 12 families and 23 genera; and the Norte zone contained 38 species from 18 families and 29 genera. In all of the zones, the Poaceae family contributed the highest number of total species: 9 species in the Centro zone, 12 species in the Meseta zone and 12 species in the Norte zone (Tab. 2).
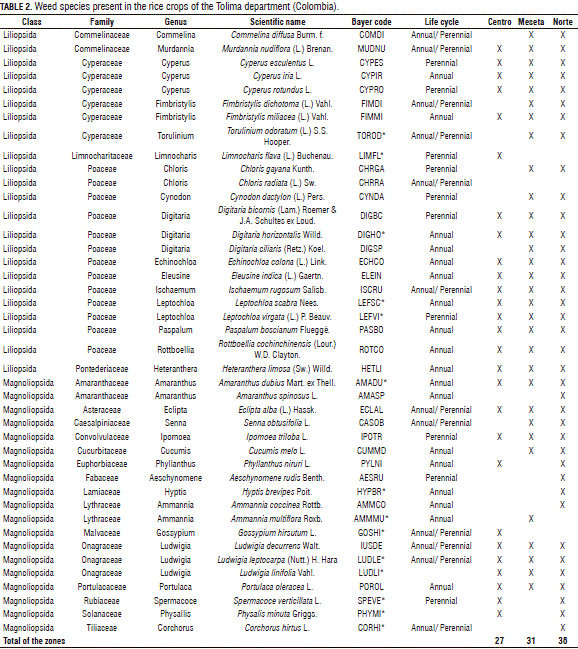
The Liliopsida class contributed the highest number of species, grouped into five (5) families, notably Poaceae and Cyperaceae. The Poaceae family contributed 13 species to the total number of weed plants in the crop. The Digitaria and Leptochloa genera contributed the higher numbers of species, with 3 and 2, respectively. These results coincide with a report made by Erasmo et al. (2004), evidencing the importance of weed species from the Liliopsida class. Rao et al. (2007) and Erasmo et al. (2004) stated that the more damaging species in this region of Colombia belong to the Poaceae and Cyperacea families. This situation is possibly due to the use of the same cultivation system for several years and to the phylogenetic relationship between the weeds and crops as they share the same requirements for resources (Radosevich et al., 1997; Puentes, 2003; Cobb and Reade, 2010). Considering these facts as well as the revelations of Inoue et al. (2012), the management of weeds in the rice crops of the Tolima department must be directed toward this segment of plants.
Importance value index (IVI)
The analysis of the IVI for the entire department demonstrated that 10 species made up 50% of the maximum importance value index, representing the more damaging weeds in the rice production systems of the region (Tab. 3). E. colona was the most important species in the rice crops of Tolima with an importance value index (IVI) of 30.4, a presence in 91.7% of the lots of the departments, a frequency of 0.39 and a density of 77.2 individuals/m2 (Tab. 3). The level of importance of the species was markedly influenced by the relative frequency, which indicated that it is a species that is adapted to the prevailing conditions of the crops. The importance of this species has been reported by different authors (Holm et al., 1991; Puentes, 2003; Rao et al., 2007; Chauhan and Johnson, 2010a). Erasmo et al. (2004) reported on the importance of this species in weed communities in rice crops of Brazil through the use of phytosociological indices.
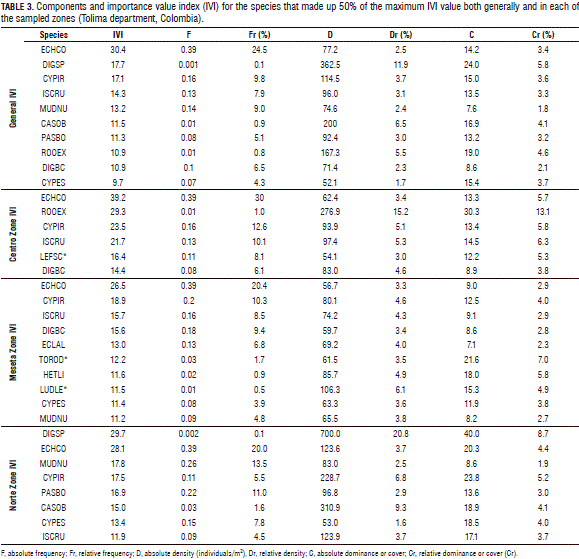
For importance, the above species was followed by D. ciliaris, C. iria, I. rugosumand M. nudiflora, which presented IVIs of 17.7, 17.1, 14.3 and 13.2%, respectively. These species together with D. bicornis and P. boscianum offered the higher values of frequency after E. colona (Tab. 3). The important presence of these species coincides with findings for rice crops in Colombia and in different locations in the world and are related to the adaptation of these species to humid conditions (Erasmo et al., 2004; Rao et al., 2007; Chauhan and Johnson, 2009a; Montealegre, 2011).
Phytosociological indices and parameters, such as the importance value index (IVI), offer a view of the composition and the distribution of plant species in a community through ecological evaluation methods (Concenço et al., 2013). IVI parameters contemplate the importance of populations within a weed community that, together with the analysis of the number of individuals and produced mass, allow for the inference of which species are more important in terms of infestation (Pitelli, 2000).
In the Centro zone, six species represented 50% of the maximum importance value index, representing the principal problem for rice crops in this zone. E. colona was the most important species with an IVI value of 39.2 (Tab. 3). It was reported in 88% of the lots of the zone with a frequency of 0.39 and a density of 62.4 individuals/m2 (Tab. 3), with relative frequency being the most important variable. R. cochinchinensis, C. iria and I. rugosum presented the higher density values (Tab. 3). R. cochinchinensis was the weed with the second highest importance in the Centro zone, with an IVI of 29.3 and a density of 276.9 individuals/m2 (Tab. 3).
In Meseta, the analysis of the zone's data demonstrated that 10 species made up 50% of the maximum importance value index, where E. colona was the most important species of the zone with an IVI value of 26.5 (Tab. 3), a presence in 90% of the lots, a frequency of 0.39, and a density of 56.7 individuals/m2 (Tab. 3); again, the relative frequency was the component with the most influence on this level of importance. C. iria, I. rugosum and D. bicornis had frequency levels of 0.20, 0.16 and 0.18, and IVI values of 18.9, 15.7 and 15.6, respectively (Tab. 3). The species with the higher densities included L. leptocarpa, H. limosa and C. iria (Tab. 3).
The IVI in the Norte zone had 8 species that made up 50% of the maximum index. In this zone, D. ciliaris was the most important weed with an IVI of 29.7 (Tab. 3). It was found in 4% of the area with a frequency of 0.002 and a density of 700 individuals/m2. The index for this species was influenced by the relative density by a high degree (Tab. 3). S. obtusifolia and C. iria presented density values of 310.94 and 228.70 individuals/m2 (Tab. 3). E. colonawas registered in 100% of the cultivated area in the north, with a frequency of 0.39, a density of 123.60 individuals/m2 and an IVI of 28.1 (Tab. 3).
There was not a significant difference between the importance indices of E. colona and D. ciliaris due to divergences in the contribution of the components. The absolute density and the relative density were higher in D. ciliaris; while the absolute frequency and relative frequency were more important in E. colona (Tab. 3). Balduino et al. (2005), in sociological studies of tree species, suggested that relative density is the parameter that contributes the most to the importance of relevant species. However, the results of the present study demonstrated that the relative frequency was determinant in the importance of the principal species. The level of adaptation of the species to the ecological conditions of the agricultural environment determined the frequency of weeds in the lots and the number of individuals that competed with the crop.
This study verified that E. colona was the most important species in the rice zones of Tolima. It was the most frequent weed with an average density of 77 plants/m2. Its negative effect means that it is considered the most problematic weed of the Graminea family in rice crops, with losses due to competition reported at 76% under densities of 280 plants/m2 (Mercado and Talatala, 1977). Chauhan and Johnson (2010b) suggested that the shading effect caused by the aerial part of E. colona could be the principal mechanism responsible for yield losses. Its level of predominance in rice crops was highlighted in Colombia and Latin America by Plaza and Hernández (2014), Fuentes et al. (2010) and Puentes (2003). This indicates that E. colona has the ability to colonize humid environments where rice crops are developed in the tropics (Puentes, 2003). Adaptive advantages such as a high capacity for production and for the germination of seeds under humid conditions (Chauhan and Johnson, 2010a, 2009a) and the plant's metabolizing of C4 (Halvorson and Guertin, 2003; Montealegre, 2011) facilitate the adaptation and establishment of E. colonapopulations under the conditions of the agricultural system in Tolima. The importance of E. colonain this region, even with the use of herbicides specific for its control, has been reported by Puentes (2003).
Diversity indices
Alpha indices analyze the diversity within a weed community. Taking into account the Shannon-Wiener (H), Simpson and Uniformity (E) indices, it was possible to observe that the zones considered in this study were differentiated by their diversity. The Meseta zone was the most diverse (Tab. 4). According to the Shannon-Wiener index, the three evaluated rice zones presented a low species diversity; however, the Meseta zone possessed a proportionally higher species diversity, with the highest value at 2.7, followed by the Norte zone with 2.6 and the Centro zone with 2.3 (Tab. 4). For its part, the lowest value for the Simpson dominance index (0.09) indicated that the weed community in the Meseta zone of Ibague has a low probability of being dominated by few species; therefore, it was more diverse (Tab. 4). For its part, the high value of the Uniformity index was also seen in this zone (0.8) (Tab. 4), suggesting a high species diversity (Booth et al., 2003).

The diversity of the Meseta zone possibly presented itself in response to the fact that the number of individuals of each species was more balanced within the community (Concenço et al., 2013). In a broad sense, the Simpson dominance coefficient and uniformity coefficient indicated that the communities were dominated by various species. This could be the explanation for the number of applications and the quantity of active ingredients of the herbicides in use (data not shown) because it is thought that, when a weed community is more diverse, it tends to require complementary control treatments and that the weeds require a high quantity of herbicides due to the differential sensitivity of the species (Kuva et al., 2007).
Similarity indices
The beta diversity indices of Jaccard and Sorensen facilitate the comparison of areas in terms of composition of the weed communities (Concenço et al., 2012a). According to Felfili and Venturoli (2000), these indices are considered elevated when they are above 0.5 (50%), at which a high similarity can be interpreted between areas. Booth et al. (2003) indicated that the values must be interpreted on a scale of 0 to 1, where 0 indicates total dissimilarity and 1 indicates absolute similarity.
The similarity indices of Jaccard and Sorensen seen in the present study showed that the composition was not homogenous, that is, there was dissimilarity for all of the comparisons carried out between the zones (Tab. 5), suggesting dissimilarity in the composition of the weed communities between the Centro, Meseta, and Norte zones despite the fact that underlying similarity factors were seen in the zones, such as the nonexistence of crop rotation and sowing intensification (Erasmo et al., 2004). However, Concenço et al. (2011) suggested that, in zones where a crop is developed in a continuous manner or with rotation over a long period of time, there will be disconnection (dissimilarity). The sampled lots in the department of Tolima have been cultivated with rice for 60 years and, for the most part, have not been the subject of a crop rotation plan at any time of the year.
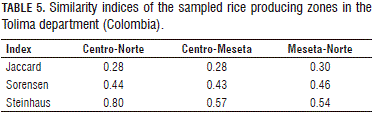
The literature contains results for the Jaccard index that vary in accordance with the climatic and agronomic management conditions; Hyvonen et al. (2003) and Fried et al. (2008) demonstrated homogeneity in weed communities of cereal crops under conditions of a temperate climate in response to the selection effect of the seasons, registering values between 0.5 and 0.8. Under tropical conditions, Concenço et al. (2012a) and Concenço et al. (2013) reported values between 0.2 and 1 due to variations in management. Ramírez (2010), for tobacco crops in the department of Huila (Colombia), found high values for this coefficient, close to 1. For the Sorensen index, Concenço et al. (2012b) and Concenço et al. (2011) reported low levels of 0.2 and high levels of 1 for tropical conditions. Furthermore, Erasmo et al. (2004) reported values between 0.22 and 0.75 in rotated rice crops: the low value was found when comparing areas of irrigated rice crops without rotation with areas with a rice-watermelon rotation, while the 0.75 value was found when comparing areas without rotation to areas with a rice-soy rotation. These authors noted the importance of the type of applied herbicide, the application timing, and the abundance of some species.
The low similarity between the weed communities was possibly due to the differences in the management of the populations because divergences were seen between the zones at the time of post-emergence herbicide application, in the application equipment, in the volumes of utilized water, and in the provenience of the seeds (data not shown). In this sense, Bernardes et al. (2011) stated that the dissimilarity of weeds between agricultural areas is explained by differences in the conditions of the soil, in the weed control methods (mechanical, cultivation, and chemical) and, mainly, in the utilization of herbicides with different mechanical actions that contribute to the selection of a more diverse flora.
The Steinhaus coefficient calculates the similarity of communities, taking into account differences in the abundance of the species, making it more precise (Booth et al., 2003). The results of the Steinhaus index for the present study registered similarity in all of the zone comparisons because values above 0.5 were found in all of the cases. The similarity was higher in the comparison between the Centro and Norte zones, indicating a high quantity of in-common species with similar levels of abundance (Tab. 5). This situation was possibly due to the similarity in the temperatures of the zones: the Centro zone registered mean maximum and minimum temperatures of 33 and 23°C, while the Norte zone registered mean maximum and minimum temperatures of 35 and 23°C.
The climatic differences between the Centro and Norte zones and the Meseta of Ibague zone were clearly observed (mean maximum and minimum temperatures of 27 and 20°C), determinant factors for the yield of the crops. The average production of the Meseta zone was 8.7 t ha-1, while, in the Centro and Norte zones, it was 7.9 and 7.6 t ha-1, respectively (Fedearroz, 2008). The critical temperatures (extremes), maximum and minimum, facilitated these divergences because they caused serious disturbances in the development of the plants. The average optimal temperature for the development of the rice plants in the vegetative phase was found between 21 and 31°C (night/day) (Yoshida, 1977). Likewise, Yoshida (1978) and Nakayama (1974) observed that temperatures that are equal to or above 35°C during the vegetative phase, which are common in the Centro and Norte zones, generate reductions in the tillering, plant height, and subsequent yield.
On the other hand, and in agreement with reports from Bernardes et al. (2011), Andreasen and Streibig (2010) and Rao et al. (2007), the repeated use of herbicides, especially with the same action mechanism, may possibly be responsible for the similarity reported in the present study due to the fact that, in all of the zones of the department, only 4 action mechanisms are used for the post-emergence active ingredients. The similarity of the weed communities agree with the results of the species inventory because it revealed that the breadth of the weed problem is represented by the same species in all of the zones of the department.
In regards to the obtained results, it was concluded that E. colonawas the principal weed for the three evaluated zones due to the fact that it presented the highest value for the importance value index and frequency. In order of importance, the following species were observed: C. iria, I. rugosum, D. bicornis, P. boscianumand M. nudiflora. The weed community of the Meseta zone was the most diverse, followed by the community of the Norte zone. The composition of the weed community in the three zones was dissimilar according to the similarity coefficients of Jaccard and Sorensen. However, the Steinhaus coefficient demonstrated that the weed communities in these rice producing zones were similar, with the highest level of similarity occurring between the communities of the Centro and Norte zones.
Literature cited
Andreasen, C. and J.C. Streibig. 2010. Evaluation of changes in weed flora in arable fields of Nordic countries - based on Danish long-term surveys. Weed Res. 51, 214-226. Doi: 10.1111/j.1365-3180.2010.00836.x [ Links ]
Balduino, A.P.C., A.L. Souza, J.A.A.M. Neto, A.F. Silva, and M.C.S. Júnior. 2005. Fitossociologia e análise comparativa da composição florística do cerrado da flora de Paraopeba-MG. Rev. Árvore 29, 25-34. Doi: 10.1590/S0100-67622005000100004 [ Links ]
Bernardes S., M.B., E.L. Finoto, D. Bolonhezi, W. Carrega, J.A.A. Albuquerque, and M.Z. Pirotta. 2011. Fitossociologia de plantas daninhas sob diferentes sistemás de manejo de solo em áreas de reforma de cana crua. Rev. Agro@mbiente On-line 5, 173-181. [ Links ]
Booth, D.B., S.D. Murphy, and C.J. Swanton. 2003. Weed ecology in natural and agricultural systems. Editorial CABI Publishing, Wallingford, UK. Doi: 10.1079/9780851995281.0000 [ Links ]
Brighenti, A.M., C. Castro, D.L.P. Gazziero, F.S. Adegas, and E. Voll. 2003. Cadastramento fitossociológico de plantas daninhas na cultura de girasol. Pesq. Agropec. Bras. 38, 651-657. [ Links ]
Cantillo, E.E., F.A. Lozada, and J.D. Pinzón. 2006. Diseño metodológico de restauración de la reserva forestal carpatos Guasca-Cundinamarca. Undergraduate thesis. Faculty of Environment and Natural Resources, Universidad Distrital Francisco José de Caldas, Bogota. [ Links ]
Concenço, G., J.C. Salton, M.L. Secretti, P.B. Mendes, R.C. Brevilieri, and L. Galon. 2011. Effect of long-term agricultural management systems on occurrence and composition of weed species. Planta Daninha 29, 515-522. Doi: 10.1590/S0100-83582011000300005 [ Links ]
Concenço, G., C.J. Silva, L.A. Staut, C.S. Pontes, L.C.A.S. Laurindo, and N.C.D.S. Souza. 2012a. Weeds occurrence in areas submitted to distinct winter crops. Planta Daninha 30, 747-755. Doi: 10.1590/S0100-83582012000400008 [ Links ]
Concenço, G., G. Ceccon, R.C. Sereia, I.V.T. Correia, and L. Galon. 2012b. Phytosociology in agricultural areas submitted to distinct wintercropping management. Planta Daninha 30, 297-304. Doi: 10.1590/S0100-83582012000200008 [ Links ]
Concenço, G., M. Tomazi, I.V.T. Correia, S.A. Santos, and L. Galon. 2013. Phytosociological surveys: tools for weed science? Planta Daninha 31, 469-482. Doi: 10.1590/S0100-83582013000200025 [ Links ]
Chauhan, B.S. and D.E. Johnson. 2009a. Ecological studies on Cyperus difformis, Cyperus iria and Fimbristylis miliacea: three troublesome annual sedge weeds of rice. Ann. Appl. Biol. 155, 103-112. Doi: 10.1111/j.1744-7348.2009.00325.x [ Links ]
Chauhan, B.S. and D.E. Johnson. 2009b. Seed germination ecology of junglerice (Echinochloa colona): a major weed of rice. Weed Sci. 57, 235-240. Doi: 10.1614/WS-08-141.1 [ Links ]
Chauhan, B.S. and D.E. Johnson. 2010a. Growth and reproduction of junglerice (Echinochloa colona) in response to water stress. Weed Sci. 58, 132-135. Doi: 10.1614/WS-D-09-00016.1 [ Links ]
Chauhan, B.S. and D.E. Johnson. 2010b. Relative importance of shoot and root competition in dry-seeded rice growing with junglerice (Echinochloa colona) and ludwigia (Ludwigia hyssopifolia). Weed Sci. 58, 295-299. Doi: 10.1614/WS-D-09-00068.1 [ Links ]
Cobb, A.H. and J.P.H. Reade. 2010. Herbicides and plant physiology. 2nd ed. J. Wiley and Sons, Oxford, UK. Doi: 10.1002/9781444327793 [ Links ]
Curtis, J.T. and R.P. McIntosh. 1950. The interrelations of certain analytic and synthetic phytosociological characters. Ecology 31, 434-455. Doi: 10.2307/1931497 [ Links ]
Erasmo, E.A.L., L.L.A. Pinheiro, and N.V. Costa. 2004. Levantamento fitossociológico das comunidades de plantas infestantes em áreas de produção de arroz irrigado cultivado sob diferentes sistemas de manejo. Planta Daninha 22, 195-201. Doi: 10.1590/S0100-83582004000200004 [ Links ]
Fedearroz, Federación Nacional de Arroceros. 2008. III Censo nacional arrocero. Bogota. [ Links ]
Felfili, J.M. and F. Venturoli. 2000. Tópicos em análise de vegetação. Comunicações Técnicas Florestais 2, 1-25. [ Links ]
Ferriol M., M. and H.B. Merle F. 2006. El Inventario fitosociológico. In: Universitat Politècnica de València, http://hdl.handle.net/10251/16818; consulted: March, 2015 [ Links ]
Fuentes, C.L., A.S. Osorio G., J.C. Granados T. and W. Piedrahita C. 2006a. Flora arvense asociada con el cultivo del arroz en el departamento del Tolima-Colombia. Bayer Cropscience; Universidad Nacional de Colombia, Bogota. [ Links ]
Fuentes, C.L., A. Fúquene, E.M. Perdomo, and S.C. Pinto. 2006b. Plántulas de especies arvenses frecuentes en la zona centro de Colombia. Universidad Nacional de Colombia, Bogota. [ Links ]
Fuentes, C.L., A. Osorio, J.C. Granados, and W. Piedrahita. 2010. Malezas de los arrozales de América Latina. pp. 387-341. In: Degiovani B., V., C.P. Martínez R., and F. Motta O. (eds.). Producción eco-eficiente del arroz en América Latina. Internacional Center of Tropical Agriculture (CIAT), Cali, Colombia. [ Links ]
Fried, G., L.R. Norton, and X. Reboud. 2008. Environmental and management factors determining weed species composition and diversity in France. Agric. Ecosyst. Environ. 128, 68-76. Doi: 10.1016/j.agee.2008.05.003 [ Links ]
Halvorson, W.L. and P. Guertin. 2003. Echinochloa Beauv. spp. U.S. Geological Survey, Southwest Biological Science Center, University of Arizona. Tucson, AZ. [ Links ]
Holm, L.G., D.L. Plucknett, J.V. Pancho, and J.P. Herberger. 1991. The world's worst weeds: distribution and biology. University Press of Hawaii, Malabar, FL. [ Links ]
Hyvonen, T., E. Ketoja, J. Salonen, H. Jalli, and J. Tiainen. 2003. Weed species diversity and community composition in organic and conventional cropping of spring cereals. Agric. Ecosyst. Environ. 97, 131-149. Doi: 10.1016/S0167-8809(03)00117-8 [ Links ]
Inoue, M.H., B.E. Silva, K.M. Pereira, D.C. Santana, P.A. Conciani, and C.L. Sztoltz. 2012. Levantamento fitossociológicoem pastagens. Planta Daninha 30, 55-63. Doi: 10.1590/S0100-83582012000100007 [ Links ]
Kuva, M.A., R.A. Pitelli, T.P. Salgado, and P.L.C.A. Alves. 2007. Fitossociologia de comunidades de plantas daninhas em agroecossistema cana-crua. Planta Daninha 25, 501-511. Doi: 10.1590/S0100-83582007000300009 [ Links ]
Lamprecht, H. 1990. Silvicultura en los trópicos: los ecosistemas forestales en los bosques tropicales y sus especies arbóreas; posibilidades y métodos para un aprovechamiento sostenido. Deutsche Gesellschaft fürTechnische Zusammenarbeit (GTZ), Eschborn, Germany. [ Links ]
Mercado, B.L. and R.L. Talatala. 1977. Competitive ability of Echinochloa colonum L. against direct-seeded lowland rice. pp. 161-165. In: Proc. 6th Asian-Pacific Weed Sci. Soc. Conf. Jakarta, Indonesia. [ Links ]
Montealegre, F.A. 2011. Morfología de plántulas de malezas de clima cálido. Produmedios, Bogota. [ Links ]
Mueller-Dombois, D. and H. Ellenberg. 1976. Aims and methods of vegetation ecology. Geogr. Rev. 66, 114-116. Doi: 10.2307/213332 [ Links ]
Nakayama, H. 1974. Panicle senescence in rice plant. Bull. Hokuriku Natl. Agric. Exp. Stn. 16, 15-17. [ Links ]
Pitelli, R.A. 2000. Estudos fitossociológicosem comunidades infestantes de agroecossistemas. J. Conserb. 1, 1-7. [ Links ]
Plaza, G. and F.A. Hernández. 2014. Effect of zone and crops rotation on Ischaemum rugosum and resistance to bispyribac-sodium in Ariari, Colombia. Planta Daninha 32, 591-599. Doi: 10.1590/S0100-83582014000300015 [ Links ]
Puentes, B. M. 2003. Flora arvense asociada al cultivo de arroz (Oryza sativa L.). MSc thesis. Faculty of Agronomy, Universidad Nacional de Colombia, Bogota. [ Links ]
Radosevich, S., J. Holt, and C. Ghersa. 1997. Weed ecology: implications for management. 2nd ed. J. Wiley and Sons, New York, NY. [ Links ]
Ramírez, R.A. 2010. Análisis de la composición florística de arvenses asociadas al cultivo de tabaco en cinco municipios del departamento del Huila. Undergraduate thesis. Faculty of Agronomy, Universidad Nacional de Colombia, Bogota. [ Links ]
Rao, A.N., D.E. Johnson, B. Sivaprasad, J.K. Ladha, and A.M. Mortimer. 2007. Weed management in direct-seeded rice. Adv. Agron. 93, 153-255. Doi: 10.1016/S0065-2113(06)93004-1 [ Links ]
Spiegel, M. 1988. Estadística: teoría y 875 problemas resueltos. 2nd ed. McGraw Hill, Madrid. [ Links ]
Yoshida, S. 1977. Rice. pp. 57-87. In: Alvim, P. and T. Kozlowski (eds.). Ecophysiology of tropical crops. Academic Press. New York, NY. Doi: 10.1016/B978-0-12-055650-2.50008-3 [ Links ]
Yoshida, S. 1978. Tropical climate and its influence on rice. International Rice Research Institute, Los Baños, Philippines. [ Links ]

