










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkAgronomía Colombiana
Print version ISSN 0120-9965
Agron. colomb. vol.33 no.1 Bogotá Jan./Apr. 2015
https://doi.org/10.15446/agron.colomb.v33n1.49368
Doi: 10.15446/agron.colomb.v33n1.49368
Scientific note
1 Sistemática de Insectos Agronomía (SIA); Línea Taxonomía de Insectos de Importancia Agrícola; Museo Entomológico UNAB, Faculty of Agricultural Sciences, Universidad Nacional de Colombia. Bogota (Colombia). rrsimbaquebac@unal.edu.co
2 Museo Entomológico Francisco Luis Gallego (MEFLG), Faculty of Sciences, Universidad Nacional de Colombia. Medellín (Colombia).
Received for publication: 26 January, 2015. Accepted for publication: 30 March, 2015.
Abstract
The gall-forming aphid Tetraneura fusiformis is recorded for the first time for Northern South America. Its identity is clarified, and descriptions of this species and that of T. nigriabdominalis, with which it is commonly confused, are offered. The association of this sap sucking insect with the ant Linepithema angulatum (Hymenoptera: Formicidae) is recorded for the first time as well.
Key words: invasive species, insects-plants, taxonomy, trophobiosis.
Resumen
Se registra por primera vez para el norte de Sudamérica el pulgón formador de agallas de las raíces Tetraneura fusiformis. Su identidad es precisada y se ofrecen las descripciones de esta especie y de T. nigriabdominalis, con la cual es comúnmente confundida. También se registra por primera vez la asociación de este insecto chupador de savia con la hormiga Linepithema angulatum (Hymenoptera: Formicidae).
Palabras clave: especies invasoras, insecto-planta, taxonomía, trofobiosis.
Gall-forming aphids (Eriosomatinae-Eriosomatini) are commonly present in both primary and secondary hosts. Usually, the hosts belong to highly unrelated botanical species, such as decidous trees as primary hosts and herbaceous plants as secondary hosts. In subtropical regions, both sexual and parthenogenetic generations of Eriosomatini switch hosts, occupying species of Ulmus and Zelkova (Ulmaceae). These aphids also attack the roots of secondary species (Akimoto, 1985; Sano and Akimoto, 2011). Along with other genera of the tribe Eriosomatini, such as Colopha, Paracolopha, and Kaltenbachiella. Tetraneura is known to produce galls in its leaves. Species are monophagous and have high specificity (Akimoto, 1985; Sano and Akimoto, 2011). Its evolution is closely linked to the host and its life cycle and distribution ranges depend on host evolution (Akimoto, 1985; Sano and Akimoto, 2011). Additionally, ant-aphid associations have been largely recognized worldwide (Delabie, 2001; Velasco et al., 2007). Ants carry the aphid nymphs from one root to another, place them in open spaces for the development of the colony, and offer protection against natural enemies. In return, ants get honeydew excreted by the aphids (Delabie, 2001).
In the present study, we recorded the species T. fusiformis (Hem. Aphididae) for the first time in Colombia and reported its association with L. angulatum (Hym. Formicidae). We also found that T. fusiformis has been commonly recorded as T. nigriabdominalis in Latin America, according to Blackman and Eastop (2006). Considering this situation, we emphasize that, for Latin America, "T. nigriabdominalis" corresponds to T. fusiformis (correctly), and offer the redescription of T. fusiformis, as well as the description of T. nigriabdominalis.
Materials and methods
We studied samples preserved in 70%-alcohol solution, housed in two insect museums at the Universidad Nacional de Colombia: Museo Entomológico UNAB (Universidad Nacional Agronomía Bogotá), Faculty of Agricultural Sciences, in Bogota, and the Museo Entomológico"Francisco Luis Gallego" (MEFLG), Faculty of Sciences, in Medellin, Colombia.
Material studied. Aphids, Tetraneura fusiformis (Eriosomatinae, Aphididae, Hemiptera). UNAB Museum, CTC, No. Catal. 701. Colombia: Antioquia, San Pedro, 6°27'N, 75°33'W, 2,450 m a.s.l., jul. 1972, coll. J. Medina ex kikuyo grass Pennisetum clandestinum and Melinis minutiflora, family Poaceae. Five slides, 5 alcohol-preserved specimens (apterous females). Ants, Linepithema angulatum. UNAB Museum, CTC, No. Catal. 997. Same data. Fourteen workers. Tetraneura nigriabdominalis (Eriosomatinae, Aphididae, Hemiptera). UNAB Museum, No. Catal. 815. Colombia, Cundinamarca, Fusagasuga, Sesquile. Represa Tomine, 5°1'55.96"N, 73°48'30.46", 2,639 m a.s.l., 19-may-2012, coll. A. Caballero, ex roots of Trifolium repens (white clover) family Fabaceae. One slide (apterous female).
For aphid clearing and mounting, we employed the procedure followed by Simbaqueba et al. (2014), established at the UNAB museum, and Posada-Flórez et al. (2014). The curatorial process consisted of specimen-clearing, slide-mounting, collection data labeling (white label), identification, and cataloguing in a database. A green label was added, containing the corresponding Catalogue Number, taxon name and determiner. The alcohol preserved, slide-mounted and pinned ants were placed in the two Central Taxonomic Collections (CTC) of both the UNAB and MEFLG museums. The aphids were determined following keys and descriptions in Blackman and Eastop (2000, 2006). For the identification of the ants, we followed the keys and descriptions shown in Wild (2007) and Longino (2010). The morphological terminology used for the ant diagnosis was adapted from Serna and Mackay (2010).
Results
Tetraneura fusiformis Matsumura, 1899 (Hemiptera: Aphididae) (Fig. 1A; often recorded in Latin America as T. nigriabdominalis (Sasaki, 1899)).
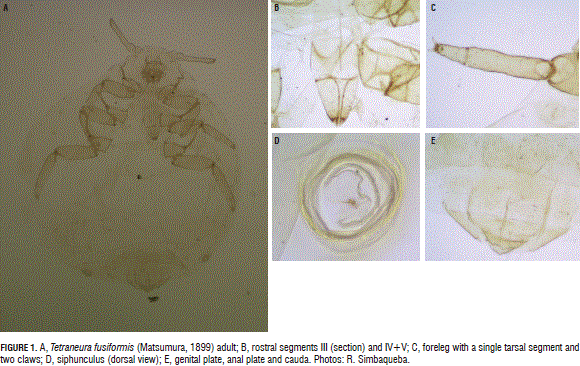
Redescription. Light green, light brown or white, apterous, globose, ovoid, appendages short; without waxy plates, or few and small when present, composed of one or two glands. Dorsum membranous, except segments VII and VIII. Head distinctly separated from thorax. Antenna 4-5 segmented, two proximal (scape and pedicel) short and wide, with one or two flagellomeres much longer and tapering, ending in a short apical one. All segments with fine setae. Processus terminalis almost same length as its base width. Secondary rhinaria lacking, primary rhinaria arranged at the posterior margin of the previous segment to the apical one, and around the apical one. Frons convex, nearly smooth, antennal tubercles not developed. Eye reduced to one triommatidium. Rostrum 5-segmented, IV and V partially or entirely fused, short and obtuse, anterior margin of the last rostral segment (IV + V) nearly equal or same length as its lateral margin, ending by the middle coxae (Fig. 1B). Thorax. Pro, meso and metathorax clearly disctinct. Mesothorax with a pair of spiracles, located at the anterior margin. Metathorax with a pair of spiracles located at the posterior margin. Legs. Fore and middle legs same size, posterior slightly longer. Tarsus one-segmented, with two claws, each with two unguitractor plates (Fig. 1C). Abdomen, membranous, except segments VII and VIII; with a lateral row of long and thick setae; without spiracular sclerites. Siphuncles with small pre- and postsiphuncular sclerite, short, similar to a low cone, wrinkled, and rimmed at the margin (Fig. 1D). Genital plate located in the segment VII, with a distinctive genital aperture. Anal plate posterior to the genital plate, with a distinctive anal aperture. Cauda semicircular, with two setae (Fig. 1E).
The biology of the species is not well understood. However, total viviparity is most likely since its life cycle is developed in both tropical and subtropical regions and off of its primary host Ulmus japonica (Ulmaceae) (Blackman and Eastop, 1994). The presence of this species is often indicated by a purple-reddish coloration on leaves (Blackman and Eastop, 2006). The species forms colonies on roots of Poaceae, including the genera Agropyron, Axonopus, Cenchrus, Chloris, Cynodon, Dactyloctenium, Echinochloa, Eleusine, Eragrostis, Oryza, Panicum, Paspalum, Pennisetum, Saccharum, Setaria, and Sorghum.
Partenogenetic apterous forms of T. fusiformis and winged females have been reported in herbaceous neotropical plants, such as grasses.
Tetraneura fusiformis is a widely distributed species in the Middle East, Africa, India, Nepal, Bangladesh, Pakistan, Sri Lanka, Andaman Islands, Thailand, Japan, Korea, Indonesia, Malaysia, Philippines, New Britain, Australia, Fiji, and Tonga. In the Americas, it is recorded in Brazil, Argentina and Central America (Blackman and Eastop, 2006; Foottit et al., 2012)
This is the first record of T. fusiformis Matsumura, 1899 for Colombia, as well as the first record of its association with Linepithema angulatum (Hymenoptera: Formicidae). Tetraneura fusiformis, possibly of Palearctic origin, has spread widely throughout warm regions of the world on Poaceae, originally its secondary host (Villalobos et al., 2010).
Tetraneura nigriabdominalis (Sasaki, 1899) (Hemiptera: Aphididae) (Fig. 2A).
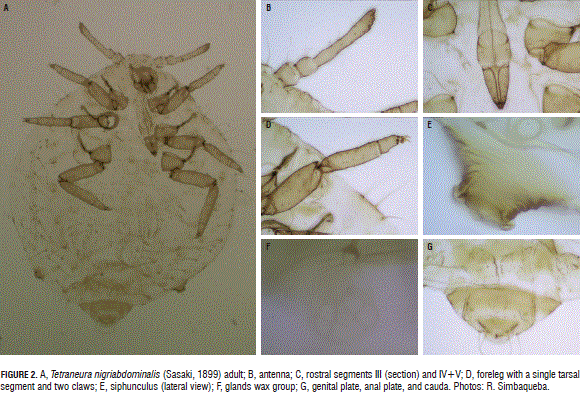
This black aphid of the roots belongs to the subfamily Eriosomatinae, tribe Eriosomatini (Blackman and Eastop, 2006).
Apterous, white or brown, globose, ovoid, appendages (legs and antennae) short. Dorsum membranous, except segments VII and VIII. Antenna 4-5-segmented (Fig. 2B), shorter than foreleg. Processus terminalis very short. Rostrum 5-segmented, apex ending between coxae III, with four accessory setae (Fig. 2C). Tarsus one-segmented (Fig. 2D). Abdomen with long lateral and stout setae, from segment I to VII. Siphuncles short and conical, with distinctive rim (Fig. 2E). With well-defined groups of waxy plates with 2 to 8 glands (Fig. 2F). Cauda rounded, possessing three setae, middle shorter (Fig. 2G). Alates. Rostral segment IV + V shorter than tarsal segment 2. Antennal segment V, 2-4X longer than antennal segment VI and same length as segment III. Median vein of forewing simple; posterior wing with one obliquos vein only. Siphuncles very short, ringlike. Cauda trapezoidal, short (Cermelli, 1973).
According to Blackman and Eastop (2006), this is an exoamerican species, distributed in Africa, Nepal, Bangladesh, Pakistan, Sri Lanka, Thailand, Indonesia, Japan, Korea, Malaysia, Philippines, Nova Britannia, Australia, Fiji, and Tonga, Colombia, Venezuela, Brazil, Peru, Argentina, and Central America (Posada, 1989; Ortego et. al., 2004; Delfino, 2005; Quiros and Emmen, 2006; Foottit et al., 2012). T. nigriabdominalis is a known pest with a worldwide distribution, especially in rice crops (Blackman and Eastop, 2000).
Comments. Blackman and Eastop (2006) considered T. nigriabdominalis to be an exoamerican species, probably from the Paleartic region and with a subcosmopolitan distribution (Blackman and Eastop, 2006). In light of this, they agreed with considering previous records of T. nigriabdominalis for Central America (Honduras, Nicaragua and Costa Rica) as T. fusiformis. According to our results, we also agreed that previous records of T. nigriabdominalis for Colombia, northern South America and Central America (Cermeli, 1970; ICA, 1972; Bustillo and Sanches, 1977; Posada, 1989; Ortego et al., 2004; Delfino, 2005; Quiros and Emmen, 2006) correspond to T. fusiformis and T. nigriabdominalis.
In the municipality of Puerto Boyaca (Colombia), T. nigriabdominalis (=T. fusiformis, correctly) is known to be associated with Paratrechina sp. (Formicidae), in the grasslands of "puntero" (ICA, 1972).
Linepithema angulatum (Mayr, 1870) (Hymenoptera: Formicidae) (Fig. 3). Worker diagnosis (Wild, 2007): Notopropodeal groove strongly impressed; dorsopropodeum straight to slightly concave; mesopleuron and metapleuron lacking pubescence and strongly shining; frons usually lacking standing setae (occasionally with a pair of subdecumbent setae in Central America); head width > 0.53 mm; color testaceous to medium brown.
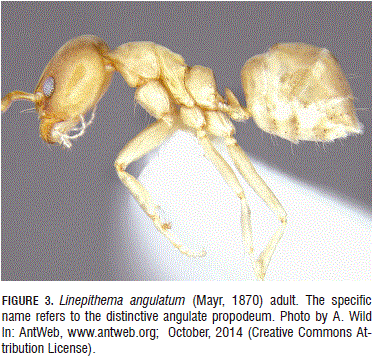
In Ecuador, there is a record for one collection of ants tending root-aphids in a nest (Wild, 2007).
Discussion of the L. angulatum identity. According to Wild (2007), some worker specimens from Central America are difficult to identify at the species level as they occasionally have a small pair of subdecumbent setae on the frons. However, this setal character is easily seen in well-curated specimens. In South America, its absence is the most reliable characteristic for separating L. angulatum from the similar but more pilose L. piliferum and L. tsachila. Central American specimens are variable in size and in color; some specimens have large eyes with nearly 70 ommatidia (elsewhere < 60), but otherwise are similar in body proportion and in the diagnostic shape of the dorsopropodeum and notopropodeal groove. Pubescence varies within the species without much geographic variation from fine and appressed to longer and somewhat wooly. Specimens from parts of Colombia and Costa Rica are frequently darker in color than specimens from elsewhere.
Wild (2007) concludes that the species boundaries of L. angulatum are still somewhat unclear. Although the broad sympatry of L. angulatum with the closely related species L. piliferum, L. tsachila and L. fuscum generally supports the recognition of these four species, variation among Central American populations and between Central American and South American populations suggests that further division of these species may become necessary. Finally, Wild stated that, given that most variations are allopatric, it is preferable to treat these ants as a single species until more material becomes available.
Acknowledgments
We thank the museums UNAB and MEFLG and their students.
Literature cited
Akimoto, S. 1985. Taxonomic study on gall aphids, Colopha, Paracolopha and Kaltenbachiella (Aphidoidea: Pemphigidae) in East Asia, with special reference to their origins and distributional patterns. Insecta matsumurana. New Series 31, 1-79. [ Links ]
Blackman, R.L. and V.F. Eastop. 1994. Aphids on the world's trees. An identification guide. CAB International, Wallingford, UK. [ Links ]
Blackman, R.L. and V.F. Eastop. 2000. Aphids on the world's crops. An identification and information guide. 2nd ed. J. Wiley & Sons, Chichester, London. [ Links ]
Blackman, R.L. and V.F. Eastop. 2006. Aphids on de world's herbaceous plants and shrubs. Host lists and keys. Vol. 1. J. Wiley & Sons, Chichester, UK. [ Links ]
Bustillo P., A.E. and G. Sánchez G. 1977. Los áfidos en Colombia: plagas que afectan los cultivos agrícolas de importancia económica. Instituto Colombiano Agropecuario (ICA), Bogota. [ Links ]
Cermeli L., M. 1970. Los áfidos (Homoptera, Aphididae) de Venezuela y sus plantas hospederas. Agron. Trop. 20(suppl. 1), 249-256. [ Links ]
Delabie, J.H.C. 2001. Trophobiosis between Formicidae and Hemiptera (Sternorrhyncha and Auchenorrhyncha): an overview. Neotrop. Entomol. 30, 501-516. Doi: 10.1590/S1519-566X2001000400001 [ Links ]
Delfino, M.A. 2005. Inventario de las asociaciones áfido-planta en el Perú. Ecol. Apl. 4, 143-158. [ Links ]
Foottit, R.G., H.E.L. Maw, K.S. Pike, and R.H. Messing. 2012. Aphids (Hemiptera: Aphididae and Adelgidae) of Hawai'i: annotated list and key to species of an adventive fauna. Pac. Sci. 66, 1-30. Doi: 10.2984/66.1.1 [ Links ]
ICA, Instituto Colombiano Agropecuario. 1972. Notas y noticas entomológicas ICA. Bogota. [ Links ]
Longino, J.T. 2010. The Ants of Costa Rica. In: The Evergreen State College, http://academic.evergreen.edu/projects/ants/home.htm; consulted: March, 2015. [ Links ]
Ortego, J., M.E. Difabio, and M.P. Mier Durante. 2004. Nuevos registros y actualización de la lista faunística de los pulgones (Hemiptera: Aphididae) de la Argentina. Rev. Soc. Entomol. Argent. 63, 19-30. [ Links ]
Posada O., L. 1989. Lista de insectos dañinos y otras plagas en Colombia. Tech. Bull. No. 43. Instituto Colombiano Agropecuario (ICA), Bogota. [ Links ]
Posada-Flórez, F.J., L. Téllez-Farfán, R. Simbaqueba, and F. Serna. 2014. Reconocimiento y observaciones bionómicas de Sipha flava (Hemiptera: Aphididae) atacando el pasto kikuyo en dos zonas de Colombia. Rev. UDCA Act. & Div. Cient. 17, 95-104. [ Links ]
Quiros, D.I. and D.A. Emmen. 2006. Diversidad biológica de los áfidos (Hemiptera: Aphididae) de Panamá. Tecnociencia 8, 63-75. [ Links ]
Sano, M. and S.I. Akimoto. 2011. Morphological phylogeny of gall-forming aphids of the tribe Eriosomatini (Aphididae: Eriosomatinae). Syst. Entomol. 36, 607-627. Doi: 10.1111/j.1365-3113.2011.00589.x [ Links ]
Serna, F. and W. Mackay. 2010. A descriptive morphology of the ant genus Procryptocerus (Hymenoptera: Formicidae). J. Insect Sci. 10. Doi: 10.1673/031.010.11101 [ Links ]
Simbaqueba C., R., F. Serna, and F.J. Posada-Flórez. 2014. Curaduría, morfología e identificación de áfidos (Hemiptera: Aphididae) del Museo Entomológico UNAB. Primera aproximación. Bol. Cient. Mus. Hist. Nat. 18, 222-246. [ Links ]
Velasco C., C., M.C. Corona-Vargas, and R. Peña-Martínez. 2007. Liometopum apiculatum (Formicidae: Dolichoderinae) y su relación trofobiotica con Hemiptera Sternorrhyncha en Tlaxco, Tlaxcala, México. Acta Zool. Mex. 23, 31-42. [ Links ]
Villalobos M., W., N. Pérez H., M.P. Mier D., and J.M. Nieto N. 2010. Aphididae (Hemiptera: Sternorrhyncha) from Costa Rica, with new records for Central America. Boln. Asoc. Esp. Ent. 34, 145-182. [ Links ]
Wild, A.L. 2007. Taxonomic revision of the ant genus Linepithema (Hymenoptera: Formicidae). Vol. 126. University of California Press, Berkeley, CA. [ Links ]

